










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.41 n.3-4 Córdoba ago./dic. 2006
Leaf anatomy of Medicinal Shrubs and Trees from Gallery Forests of the Paranaense Province (Argentina). Part 1.
Ana M. Arambarri1,*, Susana E. Freire1,2,3, Marta N. Colares1, Néstor D. Bayón1, María C. Novoa1, Claudia Monri1, and Sebastián A. Stenglein1,3
1Área de Botánica, Facultad de Ciencias Agrarias y Forestales, Calle 60 y 119, C.C. 31, 1900 La Plata, Universidad Nacional de La Plata, Argentina.
2División Plantas Vasculares, Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata, Argentina.
3Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina (CONICET).
*Corresponding author. Tel: +54-221-423-6758 int. 461; Fax: +54-221-425-2346. E-mail: anaramba@infovia.com.ar; botgral@ceres.agro.unlp.edu.ar
Summary: Sixty two species of shrubs and trees belonging to 28 families inhabiting gallery forests of the Paranaense biogeographic province (Argentina) have been cited with medicinal properties. The aim of this study was to provide a tool to recognize these taxa from fragmented leaf samples. Fresh material and hydrated herbarium specimens fixed in FAA were surveyed. The main differential traits are: hypodermis presence (e.g. Myrceugenia glaucescens ); stomata and trichome types (e.g. ciclocytic stomata in Baccharis spp.; cystolith-like structure in trichomes in Aloysia gratissima var. gratissima ); midvein transection outlines (e.g. midvein convex and keel-shaped on the adaxial side in Allophylus edulis ), mesophyll of the leaf blade (e.g. indifferentiated in Poiretia tetraphylla ); presence of idioblastic sclereids (e.g. in the petiole of Rollinia emarginata ), presence and types of crystals (e.g. crystal sand in Sambucus australis ); presence of secretory structures (e.g. large secretory cavities in Malvaceae and Myrtaceae). We can conclude that the leaf features have diagnostic value to identify species. As a result, we offer a key to distinguish these 62 species and illustrations to clarify it. Ecological interpretation of leaf structures is also given.
Key words: Anatomy; Argentina; Gallery forests; Leaves; Medicinal plants; Paranaense biogeographic province.
Resumen: Anatomía foliar de árboles y arbustos medicinales de las selvas en galería de la provincia Paranaense (Argentina). Parte 1. Entre los árboles y arbustos que forman las selvas en galería de la provincia biogeográfica Paranaense (Argentina), 62 especies pertenecientes a 28 familias son citadas como medicinales. El objetivo de este trabajo es proveer elementos para el reconocimiento de estos taxones a partir de hojas fragmentadas. Para el estudio se utilizó material fresco y ejemplares de herbario recuperados y fijados en FAA. Algunos de los principales caracteres de identificación son: la presencia de una hipodermis (e.g. en Myrceugenia glaucescens ); los tipos de estomas (e.g. los estomas ciclocíticos en Baccharis spp.) y de tricomas (e.g. los tricomas cistolíticos en Aloysia gratissima var. gratissima ); el contorno de la vena media (e.g. la vena media convexa y aquillada en la cara adaxial en Allophylus edulis ); el mesofilo foliar (e.g. indiferenciado en Poiretia tetraphylla ); la presencia de esclereidas idioblásticas (e.g. en el pecíolo de Rollinia emarginata ); la presencia y los tipos de cristales (e.g. arena cristalina en Sambucus australis ) y las estructuras secretoras (e.g. la presencia de grandes cavidades secretoras en las familias Malvaceae y Myrtaceae). Se brinda una clave e ilustraciones para la determinación de las especies así como una interpretación ecológica de las diferentes estructuras halladas en las hojas.
Palabras clave: Anatomía; Argentina; Hoja; Plantas medicinales; Provincia biogeográfica Paranaense; Selvas en galería.
INTRODUCTION
Gallery forests are situated in the northeastern Argentina, extending from Misiones to Buenos Aires political provinces. They are on the margins of the Paraná and Uruguay rivers and Delta of the Paraná river, reaching Punta Lara city on the right margin of the Río de la Plata (Fig. 1) (Dimitri, 1975; Cabrera, 1976). From a biogeographic point of view, gallery forests belong to Paranaense province within the Amazonian dominion of Neotropical region (Cabrera & Willink, 1973). In this region, more than 100 native shrubs and trees have been cited as used for food, dressing, tinctoric, religious and magic ceremonies, and in folk medicine by native population (Hieronymus, 1882; González et al ., 1939; Martínez Crovetto, 1963, 1964, 1965, 1967, 1981; Burkart, 1952, 1969; Sorarú & Bandoni, 1978; Ratera & Ratera, 1980; Toursarkissian, 1980; Amat & Vajía, 1991; Schulz, 1997; Lahitte & Hurrell, 1994, 1999; Lahitte et al ., 1998; Rondina et al., 2003). It must be noted that the flora of this area could represent a genetic reserve of great interest as a source of productive resources, forest, ornamental, aromatic, medicinal and ecological (Vervoorst, 1979; Kalesnik & Malvárez, 1996), that justifies its conservation in situ and ex situ .
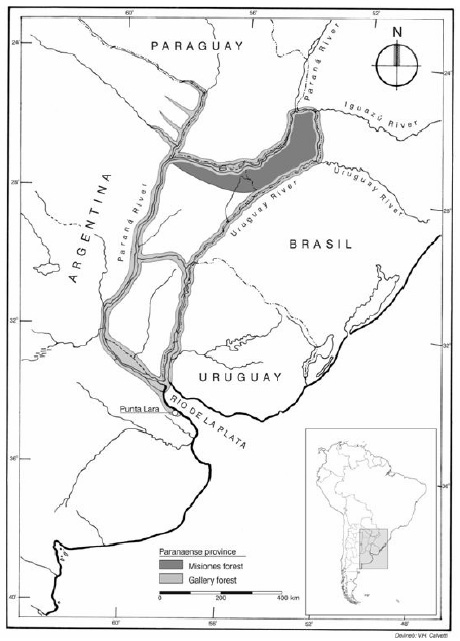
Fig. 1. Map of Paranaense province (area in gray) including the gallery forests along the Paraná and Uruguay rivers, Delta of the Paraná river, reaching Punta Lara city on the right margin of the Río de la Plata.
Throughout history, plants have been the most important source of medicines for human health. In China and India, for example, traditional medicine is largely based on plants. In the western countries the use of plants for medicine had declined, however, recently there has been a remarkable revival in the use of herbal medicines in many undeveloped and developed countries. In order to help the commercialization of medicinal plants, it would be interesting to have a tool to identify the entire or fragmented leaves of medicinal plants. Some studies of leaf architecture and leaf anatomy of trees have been carried out by Roth (1995, 1996), Ponessa et al. (1998), Leonardi et al . (2002), Mantese & Montaldo (2002), Suárez et al . (2004), Wagner & Ponessa (2004). However, it would be useful to increase the knowledge of leaf anatomy in a major number of species. In fact, as the first chapter of our leaf micromorphological analysis of native shrubs and trees with medicinal properties, we studied the leaf anatomy of 62 species belonging to 49 genera which are contained into 28 families from the marginal forests.
Owing to their relative stability, some anatomical characters like those concerning with indumentum and stomata, mesophyll and vascular system proved to be very valuable for the identification of the species. In this paper we focus our attention on leaf epidermis and leaf structure characteristics, with the aim of exploring the diagnostic value to distinguish the taxa from fragmented leaf material.
MATERIAL and METHODS
Plant materials studied: The study was performed using specimens from the herbaria (ERA, LP, LPAG, and SI) (abbreviations according to Holmgren et al ., 1990). Living leaf samples were also collected from plants growing in situ and ex situ , e.g. Paraná river, Río de La Plata and in the Botanical Garden and Arboretum "Carlos Spegazzini", Facultad de Ciencias Agrarias y Forestales de La Plata where is located the LPAG herbarium and the laboratory in which the study was carried out. The 62 studied taxa alphabetically ordered by scientific name with the family, specimens vouchers, and ethnobotanical use are shown in Appendix 1. For each studied species a summary of leaf characteristics in relation with environment conditions is shown in Table 1.
Morphology: Mature, fully developed leaves were selected for analysis. Data were obtained from the centre of the mid-lamina in the interveinal area, and from both adaxial and abaxial surfaces. For reconstitution of dried leaves we followed D'Ambrogio de Argüeso (1986). To avoid alterations on leaf samples, the material was fixed in formalin, glacial acetic acid, and 50% ethanol at 5:5:90 ratio (F. A. A.). Epidermal studies were done on transparent leaf blades after treatment with the method of Dizeo de Strittmatter (1973). In some cases, the replica method (according to Freeman, 1984) was used, and in others it was carried out by the peeling technique (D'Ambrogio de Argüeso, 1986). To analize the leaf structure free hand semithin transverse sections were obtained in the middle part of the petiole and leaf blade when they were simples. In compound leaves, transections were also obtained from the rachis and petiolule. The cleared leaf material and transections were stained with aqueous Toluidine blue 0.5% or Safranin O in 80% ethanol and mounted in gelatine-glycerine. Observations and original drawings were made by means of a light microscope provided with a camera lucida. The number of epidermal cells and stomata per field were counted twice for each species. Stomata and epidermal cells that were intersecting the edges of the observational area were not counted. The microscopic field used as the standard area was 560 µm in diameter. Stomatal index was calculated as: [no. of stomata / (no. of stomata + no. of epidermal cells)] x 100 (Salisbury, 1927). A key was prepared and a double entry was given for glabrescent species. The nomenclature follows Zuloaga & Morrone (1999). Epidermal and leaf structural features were described according to the terminology of Metcalfe & Chalk (1950, 1979), Stace (1965), and Fryns-Claessens & Van Cotthem (1973). Metcalfe & Chalk's (1950) symbols were used to represent each tissue.
RESULTS and DISCUSSION
Epidermal cells: Epidermal cells of both surfaces are arranged in a single layer, except in Ficus luschnathiana where there are 2-3-layers. This species has laticifer ducts and in its adaxial epidermis, cystoliths or calcified bodies situated in cells called lithocysts (Fig. 2C). All these traits are in agreement with diagnostic characters of Moraceae cited by Metcalfe & Chalk (1950, 1979). According to Roth (1984) pluri-stratified epidermis and thick outer epidermal cell walls on the adaxial epidermis are sun leaf traits or xeromorphic adaptations to exposed situations with strong insolation. Curiously, this species shows sun leaf traits combined with hygromorphic ones, as the parenchyma with large lacunes in the petiole, and a mesomorphic stomatal density value. In terms of size, adaxial epidermal cells are usually slightly larger than those of the abaxial epidermis. Nevertheless, Calliandra parvifolia constitutes an exception since shows very small epidermal cells and they are similar in shape and size in both surfaces. The periclinal cell walls, especially on the adaxial side, are impregnated with a layer of cutin. In general, the cuticle is not very thick, corresponding to hygromorphic or mesomorphic leaf structure (Roth, 1984). However, the family Lauraceae is characterized by the presence of stomata with curved guard cells, poral cuticle and pore plugged with cuticular substance or guard cells of stoma with scale-like extensions of cuticle. These features were found in Nectandra and Ocotea species. The cuticular ornamentation is an interesting character to distinguish genera of some families, e.g. Lauraceae, Rutaceae, and Loranthaceae as were previously cited by Metcalfe (1987), Roth (1995), Spegazzini et al . (2002), and Varela & Gurni (2003). We added Myrsine laetevirens (Myrsinaceae) with the stomata surrounded by peristomatal rim (thick ridge of cuticle). The epicuticular waxes over the cuticle are conspicuous in some taxa, e.g. Bauhinia, Calliandra, Enterolobium, Erythrina, Inga, Senna, Sesbania (Fabaceae), Myrsine (Myrsinaceae), Pouteria (Sapotaceae), Schinus (Anacardiaceae). Thick cuticles and epicuticular waxes are ways of giving an effective protection against strong winds and solar radiation that produce excesive transpiration. They also constitute the outermost defensive barrier of plant leaves against pathogens because their repellents properties (Ditsch et al ., 1995; Barthlott & Neinhuis, 1997; Barthlott et al . , 1998; Stenglein et al ., 2005).
Epicuticular waxes and/or striate cuticular ornamentation were detected on one or both surfaces. They were visible after diafanied treatment in: Allophylus edulis, Bauhinia forficata subsp . pruinosa, Calliandra parvifolia, Cestrum spp. , Croton urucurana, Enterolobium contortisiliquum, Erythrina crista-galli, Fagara spp., Ficus luschnathiana, Guarea macrophylla subsp. spicaeflora, Heteropterys angustifolia, Ligaria cuneifolia, Myrsine laetevirens, Nectandra angustifolia, Ocotea acutifolia, Pavonia sepium subsp. sepium, Phytolacca dioica, Rollinia emarginata, Salix humboldtiana, Sambucus australis, Schinus longifolia var. longifolia, Senna corymbosa, Sesbania punicea, Syagrus romanzoffiana, and Terminalia australis (Figs. 4C, 5D, 5M, 8D, 9G, and 9H).
Anticlinal epidermal cell walls are straight, curved or undulate to sinuate (Figs. 2J, 4C, 5D, 5G, 5M, 6J, 7K, 8D, 8J, 8M, 9C, 9G, and 9N). The amount of undulation is greater on the abaxial epidermis than on the adaxial one. According to Roth (1984), this is because the abaxial epidermis is exposed to conditions of greater shadow and humidity. Frequently, the anticlinal cell walls are undulate or sinuate, however, sometimes are straigh, e.g. Nectandra spp. (Lauraceae). The predominance of waived wall correlates with moderate environmental conditions of the area in which the species grow. Results are in agreement with Stace (1965) and Roth (1984) who reported that mesomorphic plants have straight or wavy anticlinal walls.
Some species present hypodermis. Phytolacca dioica (Phytolaccaceae) shows hypodermis under both epidermis, while Myrceugenia glaucescens (Myrtaceae) has only one below the adaxial epidermis (Figs. 3L, and 4F). Metcalfe & Chalk (1979) called it aqueous tissue. This water-storing tissue was cited by Roth (1995) for Myrtaceae and was reported by Gattuso (1996) in Phytolacca dioica . The hypodermal layers also are present in Syagrus romanzoffiana which belongs to the Monocotyledons. In fact, the presence of hypodermis has taxonomic implications at species level. This found is in agreement with the concept of Roth (1984, 1995) who reported that the hypodermal layers appeared in only few families and less frequently under the abaxial epidermis. This water-storing layer is more important on the adaxial surface because its function is to protect the leaf under unfavourable climatic conditions (Roth, 1995), specially the palisade parenchyma from desiccation and excessive insolation. The species of Lauraceae that are studied here do not show hypodermal tissue as was cited by Metcalfe (1987), although he also reported studies that demonstrated the lack of the hypodermis in trees growing in upper mountain rain-forests.
Stomata: The majority of the species have anomocytic stomata type (stomata without subsidiary cells). The guard-cells are oval-shaped and variable in size. They are randomly oriented as in most of Dicotyledons. However, Ligaria cuneifolia has its stomata arranged into parallel rows but transverse to the leaf blade longitudinal axis. This fact is in accordance with the description made by Varela & Gurni (2003). Syagrus romanzoffiana (Arecaceae), exhibits tetracytic stomata surrounded by four subsidiary cells, two of them parallel to the guard cells (Fig. 2J). It belongs to the Monocotyledons and its stomata are ordered parallel to the leaf blade longitudinal axis. Anisocytic stomata apparatous (with three subsidiary cells, one of them is distinctly larger or smaller than the other two) is found in Cestrum, Hibiscus, Lantana, Lippia, Monteiroa, Nicotiana, Pavonia, Poiretia, Solanum, and Sesbania . Paracytic type (with two subsidiary cells lying parallel to the guard cells) are found as the predominant stomata type in Blepharocalyx, Calliandra, Casearia, Croton, Erythrina, Eugenia, Guarea, Heteropterys, Hibiscus, Inga, Ligaria, Myrsine, Nectandra, Ocotea, Phyllanthus, Rollinia, Salix, Sapium, Senna, and Symplocos (Figs. 4C, 5M, 6J, 8J, 8M, and 9C). Less frequent, cyclocytic type (with subsidiary cells forming one or two narrow rings around the guard cells) is found in Baccharis spp. It was previously reported by Pertusi (1987), and Freire et al. (2005). Diacytic type (with two subsidiary cells lying transversely to the guard cells) is present in Lippia alba . Frequently, species have more than one stomata type. This fact was previously mentioned for the family Solanaceae by Bernardello (1982) studying Lycium and Grabowskia . Cosa de Gastiazoro (1991) established the presence of diverse stomata types in most of the genera of Solanaceae. Colares et al . (1999) found two types in Nicotiana glauca . Recently, Rigonatto et al. (2005) mentioned anisocytic and anomocytic in Solanum argentinum Bitter & Lillo. Freire et al. (2005) found anisocytic and anomocytic in Nicotiana glauca and Solanum spp . Liscovsky & Cosa (2005) reported for the genus Cestrum (Solanaceae) the presence of anomocytic, paracytic and anisocytic stomata types. The presence of various stomata types in one leaf was also found by Stenglein et al . (2003, 2005) in species of Fabaceae, e.g. anisocytic and anomocytic or paracytic and anomocytic types. In the present study, we observed that some Myrtaceae have anomocytic stomata, while others have paracytic ones, and in Eugenia uniflora are presents these two stomata types, simultaneously. These two stomata types were independently reported on Myrtaceae by Lorca et al . (1995) and Cortadi et al . (1996). Recently, Guantay (2004) found anomocytic and paracytic stomata types in Blepharocalyx salicifolius . However, we noted predominance of paracytic stomata apparatous in Myrtaceae as in the families Annonaceae, Euphorbiaceae, and Lauraceae.
In spite of the fact that according to our observations most taxa have the pair of guard cells at level with the epidermal tissue (Fig. 2D), in some cases they are slightly sunken among pappillaes (e.g. Enterolobium contortisiliquum ). Sometimes, they are slightly elevated above the rest of epidermal cells (e.g. Eupatorium inulifolium and Buddleja brasiliensis ), and they are conspicuously elevated in others (e.g. Aloysia gratissima var. gratissima, Lantana camara, and Lippia alba ). According to Bonzani et al. (1997) the closely related genera Aloysia, Lantana, and Lippia have different stomata positions: slightly elevated over the rest of epidermal cells, stomata in crypt, and stomata raised on columns which are developed with different degree of complexity. Our results agree with the observations of Bonzani et al. (1997), though we were not able to see stomata in crypts. To our knowledge the stomata apparatous raised on columns also is present in Cordia curassavica (Jacq.) Roem. & Schult. (Boraginaceae) [Arambarri et al. (2006), ined.]. This stomatal structure that was cited for the genus Cordia by Metcalfe and Chalk (1979) appears as a diagnostic character that establish some similarity among the families Verbenaceae and Boraginaceae. Species of these two families show combined hygromorphic traits (e.g. stomata elevated above surface) with xeromorphic features (e.g. high trichome density and reduced leaves). Bonzani et al. (2003) reported that species of Aloysia and Lippia have adaptations to xerophytic conditions in their stems and leaves.
Most of the species examined have hypostomatic leaves, however, amphistomatic leaves with a distribution fairly homogeneous on both surfaces are present in Baccharis coridifolia, Eupatorium ceratophyllum, E. laevigatum,, Hibiscus striatus, Ligaria cuneifolia, Nicotiana glauca, Salix humboldtiana, Solanum granulosum-leprosum, and Syagrus romanzoffiana. Other species like Cestrum parqui, Croton urucurana, Lippia alba, Monteiroa glomerata, Phyllanthus sellowianus, Poiretia tetraphylla , Sapium haematospermum, Schinus longifolia var . longifolia, and Sesbania punicea exhibit a few number of adaxial stomata representing an intermediate situation between amphistomatic and hypostomatic leaves. The presence of stomata on the adaxial surface and situated in the areolae or a few along the vecinity of midvein is a typical quality of mesomorphic leaves (Stace, 1965; Metcalfe & Chalk, 1989). According to Roth (1984) this feature is more frequent in hygromorphic shade leaves which do not have restriction in water availability.
Most of the taxa with amphistomatic leaves have lower density of stomata in their adaxial face than the abaxial one, however, in some cases is similar or higher in the adaxial rather than in the abaxial epidermis, e.g. Solanum granulosum-leprosum (Solanaceae). Bernardello (1982) reported that the species of Lycium (Solanaceae) have predominantly amphistomatic leaves, while Cosa de Gastiazoro (1991) concluded that most genera of Solanaceae are amphistomatic. Rigonatto et al . (2005) studying Solanum argentinum , also found amphistomatic leaves. Martín et al . (2001) studying Cestrum strigillatum Ruíz et Pav. (Solanaceae) observed hypostomatic leaves. Liscovsky & Cosa (2005) found predominantly hypostomatic leaves studying thirteen species of Cestrum. One exception was cited by Liscovsky & Cosa (2005) for Cestrum parqui with amphistomatic leaves. We studied this species from fresh and dry specimens inhabiting in the area of La Plata, Buenos Aires province. The leaves were found hypostomatic, though we found some stomata adaxially on the leaf blade tip. However, when we studied specimen from Formosa province, we found stomata on the adaxial surface, and distributed in middle part of the blade. It is known that certain micromorphological leaf-epidermal features such as stomatal frequency vary with environment conditions (Stace, 1965). Some reasons that may explain the epidermal variation are: the age of the leaf or rather its degree of maturity, the position of the leaf on the plant and the environment in which the plant was grown (Ashby, 1948; Stace, 1965; Sinclair & Sharma, 1971; Poole et al ., 1996).
Nevertheless, stomata indices of 15% or higher are found in Malvaceae, Myrtaceae, Solanaceae, Verbenaceae and some taxa of Fabaceae ( Inga, Poiretia , Senna ), and Malpighiaceae ( Heteropterys angustifolia ). Our results show that amphistomatic leaves and high stomatal indices are predominantly present in shrubs and small trees. We can speculate that these traits are structural adaptations to humid and shaded habitats. On the variability of stomata density, Roth (1984) reported that apparently there is a trend to increase stomata density in higher forest levels to adapt leaf structure to stronger insolation and drought. Woodward & Bazzaz (1988) mentioned that stomatal density responds to the partial pressure of CO 2 , and they found that it is directly related with the altitude (m a.s.l.). Bravo & Grau (1992) found stomatal density positively related with higher precipitations (high humidity) independently of altitude. Stenglein et al . (2005) reported an indirect relationship between altitude and stomatal density and index. The difference between all these findings and ours, might be attributed to the different factors, such as partial pressure of atmospheric CO 2 , altitudinal situation (m a.s.l.), temperature, levels of irradiance, precipitations, environmental humidity, and different reactions of each species with all these factors.
Trichomes: Leaf surfaces are glabrous or more or less densely covered by different types of glandular and non-glandular trichomes, forming an indumentum of variable texture and density. Among the 62 examined taxa, 19 are glabrous or glabrescent, and the rest are pubescent with one, two or more trichome types. The presence of cystolith-like structures in trichomes which are found on the adaxial surface of Aloysia gratissima var. gratissima (Verbenaceae) is diagnostic, because it is restricted to a limited number of species and families (Fig. 7O). Metcalfe & Chalk (1950, 1979) and Barboza et al . (2001) mentioned this microcharacter, although the former did not mention it for the genus Aloysia . In Verbenaceae we found glandular and non-glandular trichomes. The non-glandular trichomes are uniseriate with verrucose surface and they are located over more or less pronounced pedestals (emergences). This trait was exhaustively detailed by Bonzani et al. (1997). Moreover, we found this trichome organization in Cordia curassavica (Boraginaceae) [Arambarri et al. (2006), ined.], adding another similarity between these families.
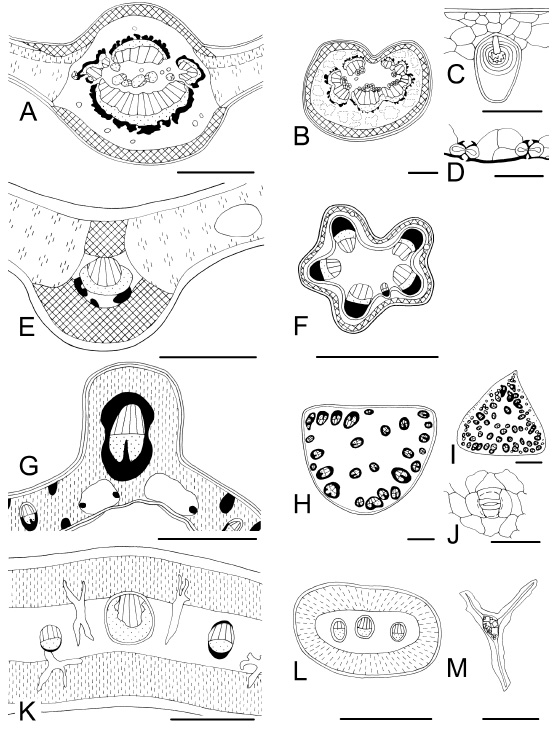
Fig. 2 . Leaf characteristics in transection and surface view. A-D : Ficus luschnathiana . A, midvein; B, aerenchymatic petiole; C, adaxial multilayered epidermis with cystoliths; D, transection of stomata on abaxial epidermis. E, F : Poiretia tetraphylla. E, midvein and indifferentiated mesophyll with secretory cavities; F, petiole outline pentalobed. G-J : Syagrus romanzoffiana . G, midvein and indifferentiated mesophyll; H, petiole (atactostele); I, rachis; J, tetracytic stoma. K-M: Ligaria cuneifolia . K, midvein, isobilateral mesophyll, and idioblastic sclereids; L, petiole, M, sclereid with crystals inside. Bars: A, B, F, I, L = 1000 µm; C, M = 100 µm; D, J = 50 µm; E, G, K = 300 µm; H = 500 µm.
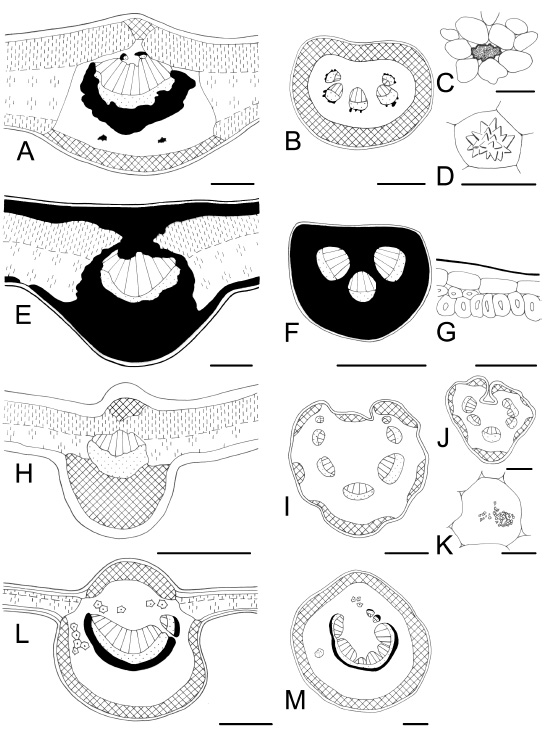
Fig. 3 . Leaf characteristics in transection. A-D : Sapium haematospermum . A, midvein and isobilateral mesophyll; B, petiole; C, a laticifer tube; D, a druse. E-G : Berberis ruscifolia. E, midvein, abundant sclerenchyma; F, petiole, fully sclerenchymatic; G, detail of subepidermal sclerechymatic cells. H-K : Sambucus australis . H, midvein, without sclerenchyma; I, bilobed petiole; J, rachis; K, crystal sand in a parenchymatic cell. L, M: Phytolacca dioica . L, midvein with idioblastic sclereids, and hypodermis on adaxial and abaxial sides; M, petiole, subcircular. Bars: A = 200 µm; B, I, J, L, M = 500 µm; C, G, K = 50 µm; D = 25 µm; E = 100 µm; F, H = 300 µm.
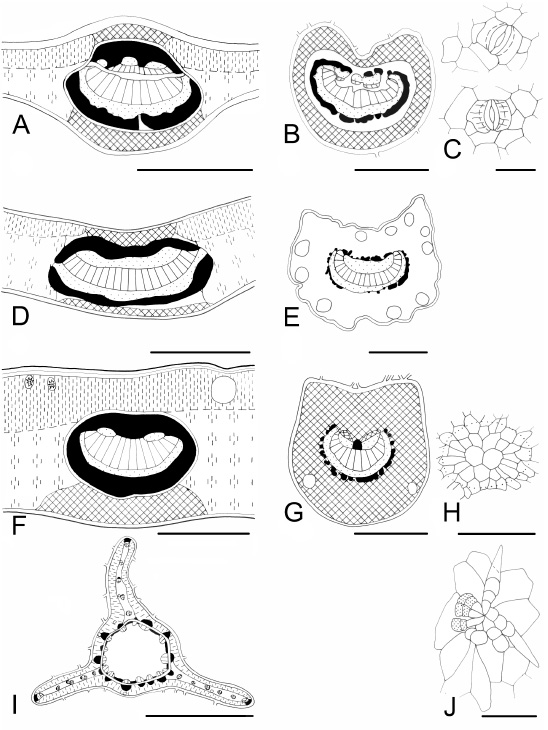
Fig. 4 . Leaf characteristics in transection and surface view. A-C : Salix humboldtiana . A, midvein, abaxial vascular bundles and inverse adaxial vascular bundles with endoscopic xylem; B, petiole; C, stomata on adaxial and abaxial epidermis, the abaxial with cuticular ornamentation. D, E : Eugenia pitanga. D, midvein, collenchyma on both sides; E, petiole, numerous secretory cavities. F-H : Myrceugenia glaucescens . F, midvein, hypodermis present and collenchyma absent on the adaxial side; G, petiole; H, a secretory cavity in surface view. I, J: Baccharis microcephala . I, stem three winged, ducts associated to vascular bundles; J, pilose nest. Bars: A, B, D-F = 300 µm; C, J = 50 µm; G = 500 µm; H = 100 µm; I = 1000 µm.
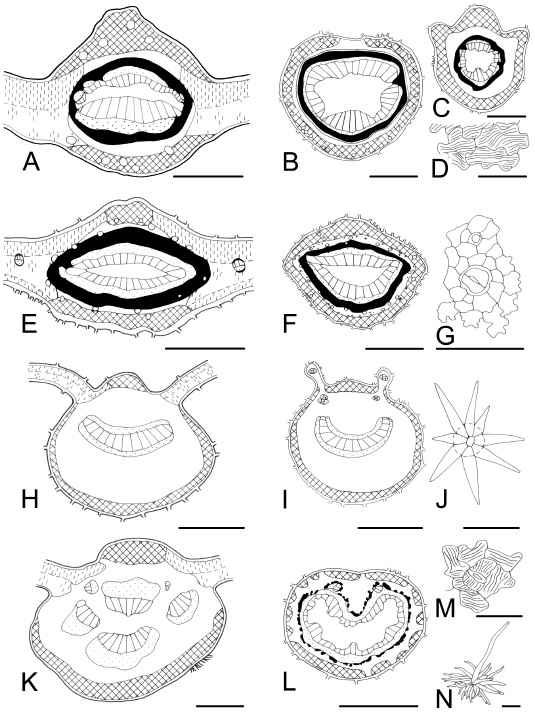
Fig. 5 . Leaf characteristics in transection and surface view. A-D : Allophylus edulis . A, adaxial keel-shaped midvein; B, petiole with ducts; C, petiolule; D, cuticular ornamentation on epidermal cells. E-G : Pouteria salicifolia. E, adaxial keel-shaped midvein; F, petiole with convex adaxial side; G, epidermis with a stoma showing peristomatal rim, and a trichome base. H-J : Solanum granulosum-leprosum . H, midvein, bicollateral vascular bundle; I, bilobed petiole; J, stellate multiangulate trichome. K-N: Croton urucurana . K, midvein with separated vascular bundles, and without sclerenchyma; L, petiole; M, cuticular ornamentation on epidermal cells; N, porrect stellate trichome. Bars: A = 200 µm; B, C, E, K = 300 µm; D, G, M = 50 µm; F, H = 500 µm; I, L = 1000 µm; J = 25 µm; N = 100 µm.
Following Metcalfe & Chalk's (1979) classification of stellate trichomes, we found these types: ( i ) stellate multiangulate sessile (Fig. 5J) and stalked over the surface are found in Solanum granulosum-leprosum (Solanaceae); ( ii ) porrect stellate (Fig. 5N) is observed in Croton urucurana (Euphorbiaceae); ( iii ) rotate (Fig. 6D) in Fagara rhoifolia (Rutaceae), Hibiscus, Monteiroa, and Pavonia species (Malvaceae), and Luehea divaricata (Tiliaceae). In addition, the family Malvaceae has two to five-armed trichomes, while Luehea has long tangled trichomes on the abaxial epidermis, and ( iv ) stellate candelabraform type (Fig. 9K) characterizes Buddleja spp. (Buddlejaceae). Within two to five-armed trichomes we observed: ( i ) two-armed T-shaped (malpighiaceous) (Fig. 8N) in Pouteria spp. (Sapotaceae) and Heteropterys angustifolia (Malpighiaceae). Within scales trichome types, peltate scales (Fig. 8G) lying in depressions of the epidermis, are present in Fagara hyemalis , F. rhoifolia (Rutaceae), and Myrsine laetevirens (Myrsinaceae). This feature agrees with Metcalfe & Chalk (1979), Roth (1995), Bruno et al. (1999), Spegazzini et al . (2002), and Wagner & Ponessa (2004).
Fagara spp. (Rutaceae) and Erythrina crista-galli (Fabaceae) have aculeate leaves with prickles on the petiole, leaf blade, and rachis. A combination of non-glandular and glandular trichomes forming pilose nests (Fig. 4J), characterize the genus Baccharis (Asteraceae) as was described by Ariza Espinar (1973), Pertusi (1987), Hellwig (1992), and Freire et al . (2005). Pilose nests formed only by glandular trichomes are observed in Eupatorium laevigatum (Asteraceae). All these trichome types were previously cited by Metcalfe & Chalk (1950, 1979) and Roth (1995), and they are highly diagnostic because of their restriction to a limited number of species.
Glandular indumentum consists in uniseriate or biseriate glandular trichomes, capitate or clavate, with a head 1-celled, 2-4-celled or multicellular. Some glandular trichomes have 1-celled and spherical head (campanulate) e.g. Aloysia gratissima var . gratissima, Lantana camara, Lippia alba , though there are with elongated and multicellular head, like in Schinus longifolia var. longifolia . Glandular trichomes are found in Aloysia, Allophylus, Baccharis, Buddleja, Calliandra, Cestrum, Croton, Eupatorium, Fagara, Guarea, Hibiscus, Inga, Lantana, Lippia, Luehea, Monteiroa, Myrsine, Pavonia, Sambucus, Schinus, Solanum, Tessaria, and Vernonia (Figs. 4J, 6C, 8G, 9G, 9J, 9O).
Some families are characterized by the presence of indumentum. Trichome morphology is an important taxonomic feature, whereas the density is a variable character (Metcalfe & Chalk, 1979). Density of trichomes does not appear associated with neither the density and size of stomata, nor with the thickness of blades and/or the outer wall of epidermal cells (Selvi & Bigazzi, 2001). We thought that trichome density is associated with irradiance and precipitation of the place where the species grow. Roth (1984) reported that trees of humid tropical forests have predominantly smooth surfaces in their leaves, mainly in their adaxial epidermis, which facilitate drainage of leaves after a heavy rain fall, and represents a barrier against microorganism infections.
Leaf blade: Midvein transection is vaulted (convex) or flattened (plane or concave) on the ventral or adaxial side, whereas it is convex on the dorsal or abaxial side. Approximately 50% of the studied species exhibit the midvein projecting to both sides (adaxial and abaxial), i.e. biconvex, whereas the rest show the midvein plane or concave on the adaxial side, i.e. plane-convex or concave-convex. One exception is Calliandra parvifolia which has its midvein transection plane on both sides. Allophylus edulis (Sapindaceae) and Pouteria salicifolia (Sapotaceae) have keel-shaped midvein projecting on the adaxial face (Figs. 5A and 5E). This is a character with diagnostic value previously reported for Allophylus by Roth (1995). Midvein vascular system : the number of vascular bundles, their distribution and relation to mechanical tissue are variable. Veins are typically collateral, with the xylem located adaxially and strands of phloem abaxially. Bicollateral veins were observed in Myrtaceae and Solanaceae as Metcalfe & Chalk (1950, 1979) and Roth (1995) did. However, the vascular system is frequently more complicated, formed by one or more abaxial vascular bundles and a variable number of inverse vascular bundles with endoscopic xylem on the adaxial side. For that reason, it appears in many cases as a bicollateral vascular bundle or sometimes forming a circle with vascular strands fusioned and surrounded by a ring of sclerenchymatic tissue (Figs. 5A, 5E, 6L, 8E, and 9E). Midveins with one vascular bundle, a row or an arc of separated vascular bundles are observed within the genera Baccharis , Berberis, Calliandra, Cassearia, Enterolobium, Eupatorium, Heteropterys, Hibiscus, Ligaria, Monteiroa, Pavonia, Phyllanthus, Phytolacca, Poiretia, Rollinia, Sambucus, Sesbania, Tessaria, and Vernonia (Figs. 2E, 2G, 2K, 3E, 3H, 6A, 7E, and 8K).
Frequently, the midvein mechanical tissue is sclerenchymatic, forming a sheat around the xylem and phloem, or a bundle cap. However, some species have sclerenchymatic tissue reduced or absent, e.g. Aloysia gratissima var. gratissima, Baccharis microcephala, Buddleja spp., Cestrum spp., Croton urucurana, Enterolobium contortisiliquum, Eupatorium ceratophyllum, E. inulifolium, Hibiscus striatus, Lantana spp ., Lippia alba, Nicotiana glauca, Sambucus australis, Schinus longifolia var . longifolia, Sesbania punicea, Solanum granulosum-leprosum, and Vernonia scorpioides (Figs. 2K, 3H, 5H, 5K, 7M, 9I, and 9L). Plants without sclerenchymatic tissue in their leaves are adapted to humid regions ( sensu Roth, 1995), and precisely these are the environmental conditions in which these taxa grow. On one hand, our results show that shrubs and small trees have not lignified mechanical tissue or at least it is not well developed. These species frequently have amphistomatic leaves and stomatal index > 15%. They seem to be adapted to less exposed, shadow and humid situations. Similar features were reported by Roth (1990) for low forests cloud stratum (1,30-5 m). On the other hand, the leaf structure of big trees increase the amount of lignified mechanical tissue, coincidentally they are more exposed to rigurose conditions. They show xeromorphic sun leaf structure of Roth (1984), and accord well with leaf characteristics mentioned by Roth (1990) for the higher stratum of cloud forests.
Predominantly, mechanical tissue of collenchymatic nature is located under both epidermis. However, is absent in Berberis ruscifolia (Berberidaceae) which is characterized by having sclerenchymatic tissue located around vascular bundles and layered under both epidermis (Fig. 3E-G). This characteristic agrees with data reported by Metcalfe & Chalk (1950) for this family. This taxon shows sclerophyllous characteristics, perhaps as one adaptation to plain areas and stronger insolation situations (Metcalfe & Chalk, 1989) or perhaps to the sandy and pedregous soils where this species grows (Hurrell & Bazzano, 2003). Collenchymatic tissue is also absent in leaves of Calliandra parvifolia (Fabaceae) and Ligaria cuneifolia (Loranthaceae). Both species have vascular bundles with phloematic fibres and in Ligaria there is crystalliferous sclerenchyma (i.e. branched stone cells containing crystals in their cavities) in the mesophyll (Fig. 2K-M). This diagnostic feature observed in Ligaria cuneifolia was previously reported by Varela & Gurni (2003).
Idioblastic sclereids appear solitary or in clusters, and they are distributed in an apparently haphazard manner in the parenchyma of the leaf blade and/or petiole for which they provide support. They constitute a diagnostic character in several species like Guarea macrophylla (Meliaceae) , Inga verna subsp. affinis (Fabaceae) , Ligaria cuneifolia (Loranthaceae), Phytolacca dioica (Phytolaccaceae), and Rollinia emarginata (Annonaceae) (Figs. 2K-M, 3L, 3M, 6L-O, 8I, 9E, and 9F) . The presence of idioblastic sclereids was mentioned by Metcalfe & Chalk (1979) and Metcalfe (1987) as a diagnostic trait for Annonaceae, Loranthaceae, Malvaceae, and Meliaceae. Now, we add to this list the families Fabaceae and Phytolaccaceae, and note that it is an important diagnostic trait mainly at specific level. As stated by Roth (1984) this microcharacter seems to be and adaptation to exposed situations with high light (sun leaf type), and frequently appears in xeromorphic leaf forms.
The majority of the taxa under study have dorsiventral mesophyll or bifacial leaves with adaxial photosyntetic tissue, usually formed by one or two layers of typical palisade cells (rarely more) e.g. Buddleja spp. Frequently, the palisade parenchyma is discontinuous above midvein, however, the chlorenchyma is well developed on the adaxial side of the midvein in Calliandra parvifolia, Enterolobium contortisiliquum, Eugenia uruguayensis, Fagara hyemalis, Ligaria cuneifolia, Myrceugenia glaucescens, Nicotiana glauca, and Schinus longifolia var . longifolia (Figs. 2K and 4F) . We agree with Roth (1995) on the fact that it is a peculiar characteristic which seldom occurs.
Most of the taxa show a palisade tissue closely packed with elongated cells, along with a well developed and predominant spongy parenchyma rich in intercellular spaces. These leaf structural traits suggest an adaptation to localities with humid conditions (Metcalfe & Chalk, 1979, 1989; Roth, 1984). A small group of taxones give evidence of having xeromorphic features and/or sun leaf characteristics (e.g. multilayered palisade parenchyma ( Buddleja, Ficus ), mesophyll indifferentiated with palisade cells ( Bauhinia, Syagrus ), and sclerenchymatic tissue around vascular tissue ( Allophylus, Berberis, Fagara, Guarea, Inga, Luehea, Myrceugenia glaucescens, Myrsine, Nectandra, Ocotea, Pouteria., Rollinia, Syagrus, Terminalia ) and under both epidermis ( Berberis ).
Some species also show an incipient isobilateral structure with abaxial photosynthetic tissue constituted by one-layered palisade parenchyma of short cells, like in Baccharis articulata, B. microcephala, Blepharocalyx salicifolius, Ficus luschnathiana, Ligaria cuneifolia, Nicotiana glauca, Pouteria gardneriana , P. salicifolia , Sapium haematospermum, and Tessaria integrifolia var. integrifolia (Figs. 2K, 3A, 4I ,5E, and 7E). Our result of isobilateral structure in Blepharocalyx salicifolius is in agreement with Cortadi et al . (1996). However, Guantay (2004) found dorsiventral mesophyll for this taxon. A similar situation may be mentioned for Salix humboldtiana and Nicotiana glauca . The latter was studied by Colares et al . (1999) who observed a dorsiventral structure, though they found an incipient isobilateral structure. This is in agreement with data reported by Cosa de Gastiazoro (1991). We may suggest that these structural differences might be related to the height of the tree, altitude (m a.s.l.), environmental humidity, degree of insolation, and possibly, soil characteristics.
An undifferentiated mesophyll is characteristic of Poiretia tetraphylla and Syagrus romanzoffiana (Fig. 2E, and 2G). In the palm Syagrus there is a homogeneous indifferentiated parenchyma in young leaves, while there is a mesophyll formed exclusively by palisade parenchyma in adult leaves. The latter also is present in Bauhinia forficata subsp. pruinosa, mesophyll structure that was previously described by Barboza et al . (2001). These results are in agreement with Roth (1984), who reported that young leaves show mesomorphic or hygromorphic characters (shade leaf type), while usually adult leaves show xeromorphic-like characters (sun leaf type). Bernardello (1982) mentioned that plants growing at great altitude or in dry environmental conditions develop a homogeneous mesophyll that is a characteristic of xerophytic habitats. In Bauhinia the presence may be attributed to insufficient water availability or special characteristics of soils.
Crystals: Appart from Asteraceae ( Baccharis spp. , Eupatorium spp. , and Tessaria integrifolia var. integrifolia ), leaves of all species present crystals, mostly solitary crystals, maclas and druses of different size (Figs. 3D, 6K, 7D, and 7L). Besides, there are: ( i ) crystal sand in Cestrum spp., Nicotiana glauca, Solanum granulosum-leprosum (Solanaceae), and Sambucus australis (Caprifoliaceae) (Fig. 3K); ( ii ) acicular crystals (styloids and raphides) in Ocotea acutifolia (Lauraceae) , Phyllanthus sellowianus (Euphorbiaceae), and Phytolacca dioica (Phytolaccaceae). All these traits correlate with Metcalfe & Chalk (1979), Cosa de Gastiazoro (1991); Colares et al . (1999), Stenglein (2001), and Freire et al . (2005).
Secretory tissue: Tannin cells are very common in all studied taxa as was cited by Metcalfe & Chalk (1979). According to Roth (1995) the plants growing at a great altitude (m a.s.l.) are exposed to strong winds, insolation and UV radiation. Tannin and resiniferous contents protect effectively, particularly against radiation. Secretory cells are usual in Allophylus edulis, Luehea divaricata, Myrsine laetevirens, Nectandra angustifolia , Ocotea acutifolia, Rollinia emarginata, and Salix humboldtiana (Fig. 7C). Secretory cavities (glands and sacs) are present in Baccharis spp., Blepharocalyx salicifolius, Casearia sylvestris, Erythrina crista-galli, Eugenia spp ., Eupatorium spp., Fagara spp. , Myrceugenia glaucescens, Poiretia tetraphylla, and Rollinia emarginata (Figs. 2E, 2F, 4D-I, 6H, 6I, 8A-C, and 8H) . Secretory canals (ducts) are found in Allophylus edulis, Croton urucurana , Ficus luschnathiana, Hibiscus striatus, Monteiroa glomerata, Myrsine laetevirens, Nectandra angustifolia, Pavonia sepium subsp . sepium, Phyllanthus sellowianus, Pouteria spp. , Rollinia emarginata, Sapium haematospermum, and Schinus longifolia var. longifolia (Figs. 2A, 2B, 3A-C, 5A, 5B, 5E, 5F, 5K, 5L, 6A, 6B, 6E, 6F, 8E, 8F, and 8H). These secretory structure were mentioned by Metcalfe & Chalk (1979) and Metcalfe (1987). The secretory tissue associated with veins in Baccharis and Eupatorium has been described as oil filled schizogenous secretory cavities by Lersten & Curtis (1986) or ducts (Cortadi & Gattuso, 1994). In Schinus longifolia var. longifolia the ducts present in the phloem have been described as resin filled schizogenous ducts by Perrota & Arambarri (2004). Blepharocalyx, Eugenia , and Myrceugenia have large secretory cavities in the leaf mesophyll and over these cavities the epidermis shows two (or three) heteromorphic cells (Fig. 4H). These characters were mentioned by Lorca et al . (1995), and are diagnostic for Myrtaceae. Some species present mucilaginiferous epidermal cells, e.g. Luehea divaricata . We may suggest that the function of these cells, like hypodermis, is to store water. On one hand, Malvaceae and Tiliaceae exhibit mucilaginiferous cavities, while on the other hand, Annonaceae and Lauraceae have oil cells. The relationships between these two last families were reported by Metcalfe (1987). The distribution of laticifer tubes in the plant kingdom is not associated to the taxonomy (Metcalfe & Chalk, 1950, 1979). However, characterizes families, e.g. Euphorbiaceae and Moraceae.
Petiole, rachis and petiolule: Petiole outline is usually semicircular or rounded in transection (Figs. 2-9). However, sometimes it can be conspicuously lobed or winged, having lobes or wings with small vascular bundles or not completely developed bundles. We found lobed or winged petiole in Aloysia, Bauhinia, Calliandra, Cestrum, Enterolobium, Inga, Lantana , Lippia, Myrsine, Poiretia , Sambucus, Sesbania, and Solanum (Figs. 8B, 8F, 8I, 9F, and 9M). The parenchyma of the petiole of two species ( Erythrina crista galli and Ficus luschnathiana ) is differentiated in an aerenchyma (Fig. 2B). This leaf structural trait suggests an adaptation to localities with very wet average conditions, and it accords well with the area where these species grow. Vascular structures of the petiole and rachis are similar to those found in the correspondent leaf midvein. The vascular tissue may be forming an arc or a circle of separated vascular bundles with or without phloematic fibres. Often, vascular bundles are arranged in a circle or in a horseshoe-shaped surrounded by a ring of sclerenchymatic tissue. Inga verna subsp. affinis shows one of the most complicated structures. It has an arc of vascular bundles on the abaxial side, and an arc of vascular bundles inversely disposed on the adaxial side with endoscopic xylem, both forming more or less a circle. In addition, there are supplementary vascular bundles in medular position. They are merged with the adaxial bundles in the petiole (Fig. 6L-N). Results are fully in agreement with Metcalfe & Chalk (1979, 1989) and Roth (1995).
Leaves show differences in their structure and enable us to distinguish them. In fact, using the epidermal characteristics and leaf structure microcharacters we offer the following key to distinguish the 62 studied taxa. In Baccharis articulata and B. microcephala the characters are taken from winged stems because their leaves are bract-like.
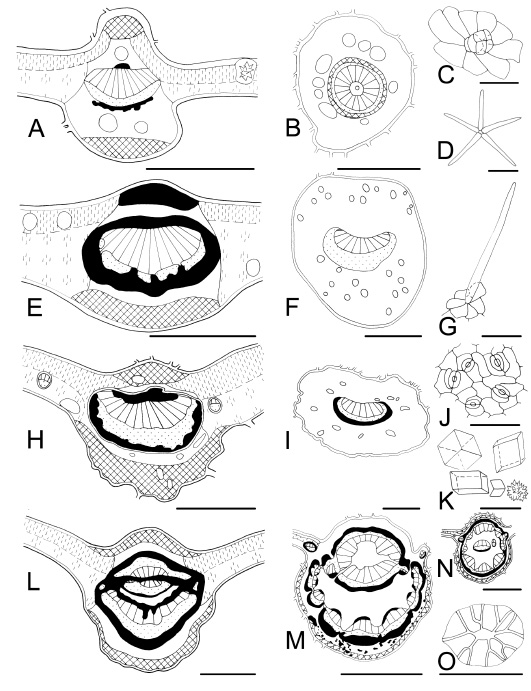
Fig. 6 . Leaf characteristics in transection and surface view. A-D : Pavonia sepium subsp . sepium . A, midvein and mesophyll with an idioblastic druse; B, petiole with large mucilage secretory cavities; C, glandular trichome, multicellular head; D, stellate trichome. E-G : Nectandra angustifolia. E, midvein shows a ring of sclerenchyma and lignified adaxial collenchyma; F, petiole with secretory tissue; G, non-glandular trichome. H-K : Casearia sylvestris . H, midvein showing xylematic and phloematic fibres; I, petiole with secretory tissue; J abaxial epidermis; K, solitary, maclas and druse of different size. L-O: Inga verna subsp. affinis . L, midvein showing a complicated distribution of vascular tissue; M, petiole with petreal cells in the cortex; N, rachis; O, an idioblastic sclereid (petreal cell). Bars: A, E, H, L = 300 µm; D, = 200 µm; B, F, I = 500; C, G, J, O = 50 µm; K = 25 µm; M, N = 1000 µm. (see figure ).
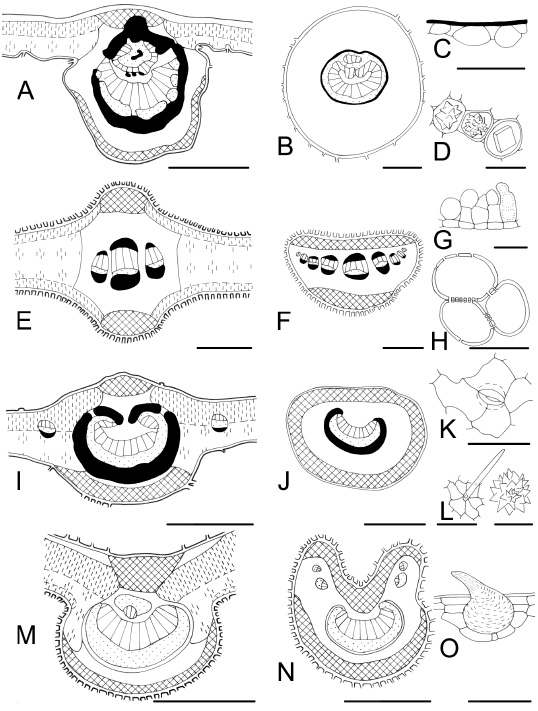
Fig. 7 . Leaf characteristics in transection and surface view. A-D : Luehea divaricata . A, midvein; B, subcircular petiole; C, epidermal mucilage cells; D, crystals. E-H : Tessaria integrifolia var. integrifolia. E, midvein, isobilateral mesophyll; F, petiole with a row of vascular bundles; G, trichomes; H, lignified parenchymatic cells. I-L : Terminalia australis . I, midvein and dorsiventral mesophyll; J, petiole, plane on ventral side; K, anomocytic stoma and anticlinal cell walls curved on the abaxial epidermis; L, trichome and druse. M-O: Aloysia gratissima var. gratissima . M, midvein with one inverse vascular bundle on the adaxial side; N, petiole with vascular bundles towards to lobes; O, cystolith-like structure in a trichome. Bars: A, E, I, M = 300 µm; B, F, J, N = 500 µm; C, O = 100 µm; D, G, H, K, L (druse) = 50 µm; L (trichome) = 200 µm.
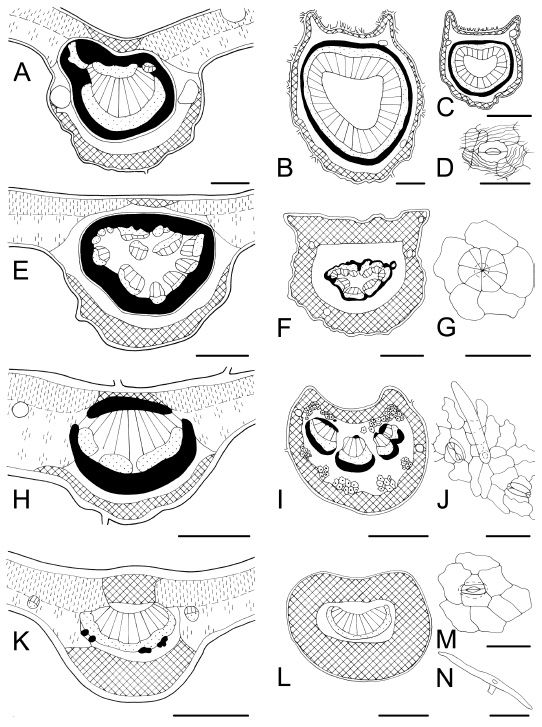
Fig. 8 . Leaf characteristics in transection and surface view. A-D : Fagara rhoifolia . A, midvein, secretory tissue; B, petiole, a circle of vascular tissue and a ring of sclerenchyma; C, rachis; D, stoma and conspicuous cuticular ornamentation. E-G : Myrsine laetevirens. E, midvein with abundant sclerenchyma; F, petiole; G, scale peltate trichome. H-J : Rollinia emarginata . H, midvein with adaxial and abaxial fibres; I, petiole characterized by clusters of idioblastic sclereids; J, abaxial epidermis showing paracytic stomata and non-glandular trichome. K-N: Heteropteris glabra . K, midvein; L, petiole, collenchymatic cortex; M, paracytic stoma; N, malpighiaceous trichome. Bars: A, G = 100 µm; B, E, H, K, N = 200 µm; C, F, I, = 500 µm; D, J, M = 50 µm; L = 300 µm.
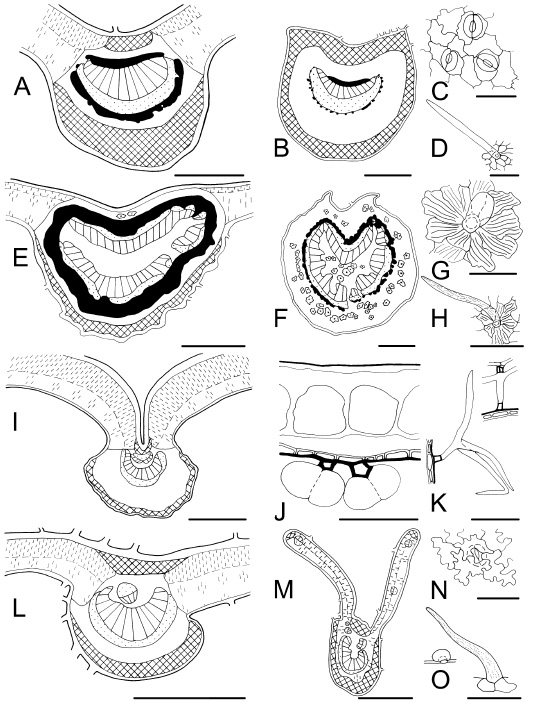
Fig. 9 . Leaf characteristics in transection and surface view. A-D : Symplocos uniflora . A, midvein; B, petiole showing continuous collenchyma; C, abaxial epidermis with paracytic stomata, and undulate anticlinal cell walls; D, non-glandular trichome. E-H : Guarea macrophylla subsp . spicaeflora. E, midvein; F, petiolule showing idioblastic sclereids with cortical and medullar distribution; G, glandular trichome and conspicuous cuticular ornamentation; H, non-glandular trichome covered by waxy papillaes. I-K : Buddleja thyrsoides . I, midvein with concave adaxial face; J, adaxial epidermis with large cells, and abaxial epidermis with short cells and glandular trichomes; K, candelabraform trichomes. L-O: Lantana megapotamica . L, midvein with one adaxial inverse vascular bundle; M, winged petiole; N, stoma and sinuate anticlinal cell walls; O, non-glandular and glandular trichomes. Bars: A, E, I, L = 300 µm; B, F, M = 500 µm; C, G, J, N = 50 µm; D, H, K, O = 100 µm.
Key to the medicinal species from gallery forests of the Paranaense biogeographic province

CONCLUSIONS
Characters value: Epidermal features and leaf anatomy have high diagnostic value at specific level and/or at family level: (1) Ficus luschnathiana can be distinguished by its multilayered epidermis; (2) a hypodermis is present in some studied species of Angiosperm-Dycots: under both epidermis in Phytolacca dioica and only below the adaxial one in Myrceugenia glaucescens , and in Angiosperm-Monocots: under both epidermis in Syagrus romanzoffiana ; (3) cystolith-like structure in trichomes are present in Aloysia gratissima var. gratissima ; (4) ciclocytic stomata type are present in Baccharis articulata and B. microcephala ; (5) Ligaria cuneifolia has an exclusive arrangement of their stomata, which are in parallel rows but transverse to the leaf blade longitudinal axis; (6) Bauhinia forficata sub sp . pruinosa and adult leaf of Syagrus romanzoffiana are characterized by having the mesophyll formed by palisade parenchyma exclusively; (7) outer periclinal epidermal cell walls are conspicuously ornamented in Lauraceae and Rutaceae; (8) stellate trichomes are diagnostic characters in Buddlejaceae, Euphorbiaceae, Malvaceae, Rutaceae, Solanaceae, and Tiliaceae; (9) bicollateral vascular bundles are diagnostic leaf traits of two families: Myrtaceae and Solanaceae; (10) stomata apparatous patterns and non-glandular and verrucose trichomes on pedestal are diagnostic characters in family Verbenaceae; 11) stomata raised on columns and non-glandular trichomes types confirm the relationships among Boraginaceae and Verbenaceae families; and 12) two species have aerenchyma in their petioles: Erythrina crista-galli and Ficus luschnathiana.Ecological implications (Table 1).
Some characters analyzed here, such as epidermal cells, stomata, mesophyll, and midvein structure correspond to mesophytic plants. For example: (1) anticlinal cell walls frequently are curved on the adaxial face and undulate on the abaxial side; (2) the majority of studied species have hypostomatic leaves, and the stomata lie at epidermis level; (3) forty six of the total sixty two analyzed taxa have dorsiventral leaves; (4) the palisade parenchyma is 1(-2)-layered in most part of species, and all species exhibit a predominant well developed spongy parenchyma rich in intercellular spaces; (5) twenty two studied species have not sclerenchymatic tissue in the midvein or it is not well developed. There are not an abrupt change from hygromorphic, mesomorphic and xeromorphic leaf structure. Some taxa show sclerophytes features, such as a little thick and/or ornamented cuticle, e.g. Ficus luschnathiana (Moraceae), Fagara spp. (Rutaceae) or with abundant sclerenchyma that surrounds the midvein, e.g. Allophylus edulis (Sapindaceae), Berberis ruscifolia (Berberidaceae), Fagara spp. (Rutaceae), Guarea macrophylla subsp . spicaeflora (Meliaceae), Inga verna subsp. affinis (Fabaceae), Luehea divaricata (Tiliaceae), Myrsine laetevirens (Myrsinaceae), Nectandra angustifolia, Ocotea acutifolia (Lauraceae), and Pouteria spp. (Sapotaceae). Scleromorphic plants are not confined to dry habitats. They also grow in humid habitats. Their xeromorphic traits may be interpreted as an adaptation to hot summers, strong insolation areas, dry periods or it may be related to the persistence of leaves or soil characteristics.
Table 1. List of studied species and ecological leaf characteristics (1).
Appendix 1. List of studied species with specimen vouchers and ethnobotany use.
Aloysia gratissima (Gillies ex Hook.) Tronc. var. gratissima (Verbenaceae). Buenos Aires, La Plata, 1-XI-1993, Chamorro s.n. (LPAG). Leaves: diaphoretic (1); digestive (2). Flowers and leaves: cardiotonic, stomachache, carminative, diaphoretic (3). Care must be take because is also mentioned as toxic for cattle (4).
Allophylus edulis (A. St.-Hil. A. Juss. & Cambess.) Radlk. (Sapindaceae). Misiones, Parque Nacional Iguazú, 21-X-1970, Dimitri & Amorín s.n. (LPAG 1227). Leaves: digestive (1); hepatic (5); cholagogue, diuretic (6); analgesic, antiseptic, febrifugal (7). Fruits: antirheumatic, depurative, to lower fevers, tonic (8-10).
Baccharis articulata (Lam.) Pers. (Asteraceae). Misiones, Apóstoles, 19-IX-1961, Fabris & Hunziker 7353 (LP). Stem: digestive, cicatrizant, (1); antirheumatic (11, 12); antiseptic, colagogue, digestive, hepatic (3, 5, 12); antidiarrhea, cardiotonic, diuretic, febrifugal (9).
Baccharis coridifolia DC. (Asteraceae). Buenos Aires, San Nicolás, 6-III-1941, Cabrera 7161 (LP). Misiones, San Ignacio, IV-1948, Schwarz 5822 (LP). Fresh plant: recommended to external use: curative of boils, eczema, herpes and others inflammatory skin process (13).
Baccharis microcephala (Lers.) DC. (Asteraceae). Misiones, Posadas, 26-I-1908, Ekman 213 (LP). Stem: hepatic (14); antidiarrhea, cardiotonic, diuretic, to lower fever (9).
Bauhinia forficata Link subsp . pruinosa (Vogel) Fortunato & Wunderlin (Fabaceae). Buenos Aires, La Plata, 15-I-1949, Burkart s.n. (LPAG 1881 ); 11-I-1966, Amorín s. n. (LPAG 1883 ); 16-XI-2004, Arambarri 250 (LPAG). Leaves: antidiabetic, astringent, diuretic (3, 11- 13, 15); expectorant (2, 15); antinephritic, antitusive. Flowers: antihemorrhoidal, digestive. External use against dandruff (9, 10).
Berberis ruscifolia Lam. (Berberidaceae). Entre Ríos, Gualeguaychú, XII-1973, Burkart et al. 30249 (SI); Federación, 25-XI-1976, Troncoso 1423 (SI). Leaves and stem: hepatic, antiinflammatory, antimalarial (3). Root: antimalarial, febrifugal (2). Root: febrifugal, antimalarial (3). Fruits: food, and tinctorial (9, 16). Toxic (3).
Blepharocalyx salicifolius (Kunth.) O. Berg. (Myrtaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, 8-II-1966, Amorín 1 (LPAG). Leaves: antiasthmatic, antidiarrhea, antirheumatic, antiseptic, astringent, bronchitis, digestive, expectorant, tonic (7-9, 12, 15, 17-19). External use: vulnerary (10).
Buddleja brasiliensis Jacq. ex Spr. (Buddlejaceae). Corrientes, Esquina, 23-X-1977 , Ahumada et al. 1427 (LP). Entre Ríos, islas, IX-1957, Ferreira 1984 (LP). Leaves: astringent; antihemorrohidal; emollient (2, 20) antiasthmatic, antitusive (3); antiasthmatic, anticatarrhal (9). Leaves and flowers: increases sweating (7).
Buddleja thyrsoides Lam. (Buddlejaceae). Entre Ríos, Delta del Paraná, 22-XI-1931, Burkart 1974 (LP). Leaves: antiasthmatic, anticatarrhal (9, 13).
Calliandra parvifolia (Hook. f. & Arn.) Speg. (Fabaceae). Buenos Aires, La Plata, Facultad de Agronomía, X-1949, Arzuaga 194 (LPAG); 13-III-1950, Arzuaga s.n. (LPAG). Potential medicinal use (21).
Casearia sylvestris Sw. (Flacourtiaceae). Corrientes, 8-VIII-1973, Schinini, Tressens & Benítez 6905 (LP). Leaves and bark: antiinflammatory, cicatrizant, febrifugal (2). External use: vulnerary, insect repellent, antidote of bite viper; antidiarrheic, depurative, antirrheumatic (22).
Cestrum euanthes Schltdl. (Solanaceae). Buenos Aires, Magdalena, 23-IV-1995 , Bayón 283a (LPAG). Bark: cathartic, increases sweating, tempering (2); febrifugal (7).
Cestrum parqui L´Her. (Solanaceae). Buenos Aires, La Plata, 21-II-1995, Bayón 123, 128; 19-III-1995, Bayón 199, 200a (LPAG). Entre Ríos, Concordia, 20-XII-1972, Volkart s.n. (LPAG). Formosa, Bermejo, 14-XII-2004, Moreno & Bayón 947 (LPAG). Recommendation: external use. It is a toxic species. Leaves: have properties as analgesic, antihemorrhoidal (11); eczema, fungal infections, sudorific. Fruits: febrifugal, madurative (2); emmollient, desinflamatory (9). Root: drastic, sudorific (9).
Croton urucurana Baill. (Euphorbiaceae). Corrientes, Esquina, 20-VII-2004, Bayón 785, 788 (LPAG). Misiones, Posadas, Rodríguez 4 (SI). Leaves: astringent, resolutive (2). Latex of seeds and bark: curative of mouth ulcers in infants (7).
Enterolobium contortisiliquum (Vell.) Morong (Fabaceae). Buenos Aires, La Plata, XI-2004, Montes de Oca s.n. (LPAG); X-2004, Arambarri 248 (LPAG). Misiones, Puerto Iguazú, XI-1970, Dimitri ( 3156 LPAG). It is cited as toxic for fish and cattle (8, 9). Medicinal properties. Leaves: astringent (2); vulnerary (8, 9). Bark: analgesic. Ripe fruits: capillary tonic and anti-dandruff. Root, bark, and fruits: anti-blennorrheic, expectorant (10).
Erythrina crista-galli L. (Fabaceae). Buenos Aires, La Plata, 26-XI-1998, Alberto & Aguiar 52 (LPAG); La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, 3-XI-2004, Arambarri s.n. (LPAG). Leaves, bark and flowers: sedative, antiseptic, narcotic (2, 11, 14). Leaves: antihemorroidal (2, 9, 11). Bark: cicatrizant (13), antiasthmatic, astringent (9, 12). Flowers: antitusive, expectorant, hypnotic (9).
Eugenia involucrata DC. (Myrtaceae). Misiones, Parque Nacional Iguazú, 14-IX-1970, Dimitri & Amorín s.n. (LPAG). Potential use stomachache (10).
Eugenia pitanga (O. Berg.) Kiaresk. (Myrtaceae). Misiones, Candelaria, 8-IX-1945, Montes 2522 (LP). Leaves: antirheumatic, aromatic, balsamic (2); antihypertensive, digestive, hepatic (22). Root: astringent (2).
Eugenia uniflora L. (Myrtaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales. 25-VI-1966, Amorín s.n. (LPAG). Leaves: aromatic, diuretic (18, 25); antidiarrhea, carminative, digestive, emmenagogue (9, 10). Leaves and stem: antyhypertensive (24). Bark: astringent, diuretic (11); emolient (9, 10).
Eugenia uruguayensis Cambess. (Myrtaceae). Buenos Aires, Isla Martín García, 20-XII-1991, Hurrell et al. 1100 (LP). Potential use as E. uniflora (8).
Eupatorium ceratophyllum Hook. & Arn. (Asteraceae). Entre Ríos, Paraná, s. f., Salellas 31 (LP). Córdoba, s. f., Kuntze s. n. (LP). Digestive, febrifugal, hepatic (26).
Eupatorium inulifolium Kunth (Asteraceae). Misiones, Posadas, I-1907, Spegazzini 13650/13700 (LP). Anticonception; emenagogue (2, 3, 27).
Eupatorium laevigatum Lam. (Asteraceae). Misiones, Yabebyry, 23-IV-1945, Montes 830 (LP); Santa Ana, s. f. , Ekman s. n . (LP). Anticephalalgia, drastic (2). Antibacterial and antifungal activities (28).
Fagara hyemalis (A. St. Hil.) Engl. (Rutaceae). Buenos Aires, Isla Martín García, 12-XI-1994, Hurrell 2103 (LP). Corrientes, Santa Teresa, 16-X-1954, Cabrera 11804 (LP). Bark: emmetic (29). External use: analgesic (earache; teeth pains) (2). Some people think that the decoction of the thorns of the trunk stops alcoholism (7, 8). Bark and leaves: antirheumatic, diuretic, sudorific, tonic (9).
Fagara rhoifolia (Lam.) Engl. (Rutaceae). Corrientes, Santo Tomé, 11-II-1972, Krapovickas, Cristóbal, Maruñak, Moroginelli, Pire & Pueyo s.n. (LP). Entre Ríos, Villaguay, XII-1942, Devoto s. n. (LP). Leaves: antirheumatic (7). Bark: analgesic (earache; teeth pains), tonic (13); diaphoretic, diuretic, sialagogue (2); thorns of the trunk stops alcoholism (7).
Ficus luschnathiana (Miq.) Miq. (Moraceae). Buenos Aires, La Plata, X-2004, Santander ( 3158 LPAG). Leaves: purgative (8). Latex: analgesic (8). Fruits: sedative (10). External use: vulnerary (10).
Guarea macrophylla Vahl subsp. spicaeflora (A. Juss.) T.D. Penn. (Meliaceae). Misiones, Iguazú. X-1975, Volkart s. n. (LPAG). Bark: antirheumatic, astringent (2).
Heteropterys glabra (Griseb.) Anderson (Malpighiaceae). Corrientes, Empedrado, 25-II-1957, Pedersen 4667 (LP). Leaves and flowers: sedative (5, 14).
Hibiscus striatus Cav. (Malvaceae). Buenos Aires, Isla Martín García, 30-XII-1999, Hurrell et al. 4148 (LP). Flowers: emollient, pectoral (2).
Inga verna Willd. subsp. affinis (DC.) T.D. Penn. (Fabaceae) . Misiones, Parque Nacional Iguazú, 12-XI-1969, Volkart s.n. (LPAG); Cataratas de Iguazú, 15-XI-1971, Dimitri s.n. (LPAG). Leaves and bark: astringent, expectorant antineuralgic, antiseptic (9, 10). Resin of fruits: analgesic (2, 8).
Lantana camara L. (Verbenaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Ciencias Agrarias, VIII-2004, Arambarri s.n. (LPAG). Misiones, Parque Nacional de Iguazú,16-IX-1970, Dimitri & Amorín s.n . (LPAG). Care must be taken, plant and inmature fruits are toxic (8). Aromatic, digestive, sedative (13). Leaves: antirrheumatic, anticatarrhal, tonic (15). Stem: antispasmodic, carminative (2). Diuretic, febrifugal (9). Flowers: pectoral (15).
Lantana megapotamica (Spreng.) Tronc. (Verbenaceae). Buenos Aires, Magdalena, 23-IV-1995, Bayón 273 (LPAG). Antirrheumatic, diuretic, febrifugal (3, 9).
Lantana montevidensis (Spreng.) Briq. (Verbenaceae). Entre Ríos, Villaguay, 7-IV-1999, Muñoz 5184 (ERA). Leaves: analgesic, febrifugal (leafy stem) (8); anticatarrhal, antirrheumatic, antispasmodic, carminative, digestive, sudorific (2).
Ligaria cuneifolia (Ruiz & Pav.) Tiegh. (Loranthaceae). Buenos Aires, Isla Martín García, 20-II-1998, Hurrell et al. 3884 (LP); Isla Martín García, 16-III-2000, Delucchi 2383 (LP). Leaves, flowers and fruits: control fracture pains, cicatrizant (9, 15). Leaves and stem: hypertensile and inmunology (30).
Lippia alba (Mill.) N.E. Br. (Verbenaceae). Buenos Aires, Barrancas del Sur, 2-V-1902, Venturi 117 (SI); La Plata Estación Experimental J. Hirschhorn, XI-2003, Martínez & Arambarri s.n. (LPAG). Leaves: antihemorrhoidal (external use), digestive, antispasmodic, diuretic, emmenagogue (2, 11, 15). Leaves and stem: catarrh (5).
Luehea divaricata Mart. (Tiliaceae) . Buenos Aires, La Plata, Jardín Botánico, Agronomía, 2-V-1973, Volkart s.n. (LPAG); 15-XI-1966, Capital Federal, Amorin s. n. (LPAG). Leaves and flowers: digestive, tonic. Flowers: sedative (13, 15). Leaves: antiinflammatory, diuretic (2, 15). Bark: antidiarrhea, astringent, digestive, febrifugal, hepatic, tonic (9, 15).
Monteiroa glomerata (Hook. & Arn.) Krapov. (Malvaceae). Buenos Aires, Punta Lara, 16-X-1979, Freire s.n . (LPAG). Diuretic, emolient (2).
Myrceugenia glaucescens (Cambes.) D. Legrand & Kausel (Myrtaceae). Buenos Aires, Isla Martín García, 17-XII-1991, Hurrell et al. 1000 (LP). Leaves: astringent (13); antidiarrhea, digestive (10).
Myrsine laetevirens (Mez.) Arrechav. (Myrsinaceae). Misiones, San Ignacio, 14-II-1946, Montes 1924 (LP). Medicinal (18). Leaves are used in adulterating yerba mate ( Ilex paraguariensis St. Hil.), but are toxic due to the presence of saponin and quinonic pigments (8, 10).
Nectandra angustifolia (Schrad.) Nees & Mart. ex Nees (Lauraceae). Entre Ríos, Concordia, 12-X-1968, Cabrera & Sagastegui 19331 (LP). Bark: purgative (2); digestive (23).
Nicotiana glauca Graham (Solanaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, 6-XII-1996, Bayón 371b (LPAG); 30-X-1997, Colares 6 (LPAG). Formosa, Bermejo, 17-20-V-2004, Moreno & Bayón 742 (LPAG). Recommendation: external use because it has narcotic properties. External use. Leaves: analgesic (12, 13); antiinflamatory (1, 12); antirrheumatic antihemorrhoidal (3, 11, 12). Antipruriginosa, madurative, (2). Skin infections (8). Cicatrizant, antihemorrhoidal, antirheumatic (9, 12, 15).
Ocotea acutifolia (Nees) Mez. (Lauraceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, X-2004, Monti s. n. (LPAG). Potential use, cortex as astringent. It contains essential oils, aromatic resins, saponins and oxidases on leafy stems.
Pavonia sepium A. St.-Hil. subsp. sepium (Malvaceae). Buenos Aires, Pereyra, 24-III-1966, . Amorín s.n. (LPAG). Leaves: emollient, pectoral (2).
Phyllanthus sellowianus Müll. Arg. (Euphorbiaceae). Buenos Aires, Punta Lara, La Plata, 18-IV-2004, Bayón 637, 638 (LPAG). Bark, leaves, root, and stem: drastic (13); antihipertensive, antiasthmatic (8, 12). Bark and leaves: antidiabetic, antiseptic, diuretic, drastic (11, 12, 15); antidiarrhoea, antiasthmatic, and vulnerary (9, 12).
Phytolacca dioica L. (Phytolaccaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, 21-X-2004 , Martínez 17 (LPAG). External use. Leaves: antiseptic (15). Leaves, fruits, root: vulnerary; drastic, emetic, febrifugal (2, 11). Root: antirrheumatic. Bark: antiseptic, astringent (9).
Poiretia tetraphylla (Poir.) Burkart (Fabaceae). Corrientes, Empedrado, 30-XI-1954, Pedersen 3013 (LP). Stomachic, sudorific, tonic (31). Leaves: antiphlogistic, antihelminthic, vulnerary, tonic (2). Suspicious of toxicity (7).
Pouteria gardneriana (A. DC.) Radlk. Toursark. (Sapotaceae). Misiones, Loreto, 2-XI-1931, Gruner 1050 (LP). Leaves: vulnerary (2).
Pouteria salicifolia (Spreng.) Radlk. (Sapotaceae). Misiones, San Ignacio, 1-XII-1931, Gruner 1082 (LP). Leaves: vulnerary (2, 8, 9).
Rollinia emarginata Schltdl. (Annonaceae). Buenos Aires, La Plata, Facultad de Ciencias Agrarias y Forestales, XI-2004, Montes de Oca s. n . (LPAG). Leaves: analgesic, antiseptic (23, 29).
Salix humboldtiana Willd. (Salicaceae). Buenos Aires, Isla Martín García, 16-XII-1991, Hurrell et al. 948 (LP); XI-2004, Monti 27 (LPAG). Bark: antiseptic, eczema, febrifugal, tonic (11); astringent (2); analgesic, antirheumatic, digestive, sedative (9). The decoction of bark is a quinine substitute (10).
Sambucus australis Cham. & Schltdl. (Caprifoliaceae). Buenos Aires, Berazategui, 2-V-2004, Bayón 641 (LPAG); V-2005, Bayón 1089 (LPAG). Leaves: antiinflammatory, cicatrizant (2); antihemorrhoidal, laxative, antitusive (3, 9). Bark: catartic (2). Bark rooth and flowers: diuretic (3). Flowers: sudorific (2); digestive (3, 9). Fruits: depurative (9).
Sapium haematospermum Müll. Arg. (Euphorbiaceae). Buenos Aires, La Plata, 18-IV-2004, Bayón 635 ; 27-IV-2004, Bayón 641 ; 5-V-2005, Bayón 1087 (LPAG). Care must taken with the use of this species. Toxic: seed (3). Recommendation external use. Leaves: febrifugal (3). Bark: cicatrizant. Latex: to cure boils, cutaneous bores (8). Vulnerary (3, 9).
Schinus longifolia (Lindl.) Speg. var . longifolia (Anacardiaceae). Buenos Aires, Isla Martín García, 12-XI-1992, Hurrell et al., 1374 (LP). Córdoba, San Jerónimo, 3-II-1948, Job 2598 (LP). Leaves: expectorant, laxative. Resin: in external use to soften hardness skin (9).
Senna corymbosa (Lam.) H.S. Irwin & Barneby (Fabaceae). Buenos Aires, La Plata, Jardín Botánico C. Spegazzini, Facultad de Ciencias Agrarias y Forestales, 12-III-1984, Florez s.n .; 30-IX-1997, Arambarri 242 (LPAG). Leaves, seeds and root: drastic (8); external use: analgesic, emollient (3, 9).
Sesbania punicea (Cav.) Benth. (Fabaceae). Buenos Aires, Punta Lara, III-1979, Freire s.n . (LPAG); City Bell, 18-III-2005, Bayón 1087 (LPAG). Care must be taken, it may be toxic.(8). Bark: astringent (7).
Solanum granulosum-leprosum Dunal (Solanaceae). Misiones, Parque Nacional Iguazú, 17-XI-1969, Hualde 81 (LPAG). Leaves: antiinflammatory (7). Fruits: anestesic (2).
Syagrus romanzoffiana (Cham.) Glassm. (Arecaceae). Misiones, Leandro N. Alem, 9-IV-1971, Maruñak 185 (LP); Buenos Aires, La Plata, VII- 1994, Colares 49 (LPAG). Root: anticonception (2).
Symplocos uniflora (Pohl) Benth. (Symplocaceae). Misiones, Parque Nacional Iguazú, 25-IX-1970, Dimitri & Amorín 159 (LPAG); Buenos Aires, Isla Martín García, 30-X-1999, Hurrell et al. 4163 (LP). Leaves are used in adulterating yerba mate ( Ilex paraguariensis St. Hil.) (8).
Terminalia australis Cambess. (Combretaceae). Buenos Aires, Punta Lara, 11-III-1979, Freire s. n . (LPAG 5544 ). Bark: astringent (2, 8, 9). Leaves and stem: bactericidal activity (32).
Tessaria integrifolia Ruiz et Pav. var. integrifolia (Asteraceae). Buenos Aires, Punta Lara, 18-IV-2004, Bayón 634 (LPAG). Antigonorrheal (2). Leaves and flowers: antiallergic, antiasthmatic, antiinflammatory, diuretic, bactericidal activity (12).
Vernonia scorpioides (Lam.) Pers. (Asteraceae). Buenos Aires, Isla Martín García, 22-XII-1991, Hurrell et al. 1126 (LP). Medicinal (26).
(1)Martínez Crovetto (1964); (2) Toursarkissian (1980); (3) Barboza et al. (2006); (4) Ragonese & Milano (1984); (5) Amat & Vajía (1991); (6) Yajía et al . (1999); (7) Rondina et al . (2003); (8) Lahitte & Hurrell (1994); (9) Lahitte et al. (1998); (10) Lahitte & Hurrell (1999); (11) Sorarú & Bandoni (1978); (12) Mandrile (2005); (13) González et al. (1939); (14) Schulz (1997); (15) Alonso & Desmachelier (2005); (16) Hurrell & Bazzano (2003); (17) Lahitte & Hurrell (1995); (18) Cortadi et al. (1996); (19) Mantese & Montaldo (2002); (20) Carrizo & Isasmendi (1994); (21) Kalesnik & Malvárez (1996); (22) Paz et al. (1992); (23) Martínez Crovetto (1981); (24) Lorca et al. (1995); (25) Amat et al. (1999); (26) Freire & Urtubey (1999); (27) Cortadi & Gattuso (1994); (28) Clavin et al . (1999); (29) Martínez Crovetto (1965); (30) Varela & Gurni (2003); (31) Burkart (1952); (32) Castro et al. (2001).
ACKNOWLEDGMENTS
The authors appreciate the fine technical assistance of Santiago M. Martínez in collecting material, and are grateful to the curators of herbaria ERA, LP, LPAG, and SI for loaning the specimens. They also thank to Víctor H. Calvetti who prepared the map, María A. Migoya for inking our pencil original illustrations, and the reviewers for critically reading the manuscript and for their valuable suggestions. Support for this study by the Comisión de Incentivos a los docentes-investigadores, Decreto 2427/93, Secretaría de Política Universitaria del Ministerio de Educación de la Nación Argentina is gratefully acknowledged.
BIBLIOGRAPHY
ALONSO, J & C. DESMACHELIER. 2005. Plantas medicinales autóctonas de la Argentina. Bases científicas para su aplicación en atención primaria de la salud. Ed. L.O.L.A., Buenos Aires. [ Links ]
AMAT, A. G. & M. E. VAJÍA. 1991. Plantas medicinales y etnofarmacología en la provincia de Misiones (Argentina). Acta Farm . Bonaerense 10: 153-159. [ Links ]
AMAT, A. G., G. A. DE BATTISTA & R. F. ULIANA. 1999. Diuretic activity of Eugenia uniflora L. (Myrtaceae) aqueous extract. Acta Hort . 501: 155-158. [ Links ]
ARAMBARRI, A. M., S. E. FREIRE, N. D. BAYÓN, M. N. COLARES, C. MONTI, M. C. NOVOA, S. A. STENGLEIN, S. M. MARTÍNEZ & V. G. PERROTTA. 2006. Anatomía foliar de árboles y arbustos medicinales de Misiones (Argentina). Provincia biogeográfica paranaense (inédito). [ Links ]
ARIZA ESPINAR, L. 1973. Las especies de Baccharis de Argentina Central. Bol . Acad. Nac. Ci., Córdoba 50: 175-305. [ Links ]
ASHBY, E. 1948. Studies in the morphogenesis of leaves. II. The area, cell size and number of leaves of Ipomoea in relation to their position on the shoot. New Phytol. 47: 177-195. [ Links ]
BARBOZA, G. E., N. BONZANI, E. M. FILIPPA, M. C. LUJÁN, R. MORERO, M. BUGATTI, N. DECOLATTI & L. ARIZA ESPINAR. 2001. Atlas histo-morfológico de plantas de interés medicinal de uso corriente en Argentina. Museo Botánico de Córdoba. Serie Especial 1. Argentina. [ Links ]
BARBOZA, G. E., J. J. CANTERO, C. O. NÚÑEZ & L. ARIZA ESPINAR (eds). 2006. Flora medicinal de la provincia de Córdoba (Argentina). Museo Botánico de Córdoba. Gráficamente ediciones, Córdoba. Argentina. [ Links ]
BARTHLOTT, W. & C. NEINHUIS. 1997. Purity of the sacred lotus, or escape from contamination in biological surfaces. Planta 202: 1-8. [ Links ]
BARTHLOTT, W., C. NEINHUIS, D. CUTTLER, F. DITSCH, I. MEUSEL, I. THEISEN, & H. WHILHELMI. 1998. Classification and terminology of plant epicuticular waxes . J. Linn. Soc. Lond. Bot . 126: 237-260. [ Links ]
BERNARDELLO, L. M. 1982. Estudios en Lycium (Solanaceae) I. Anatomía de hoja y tallo, y sus diferencias con Grabowskia . Bol. Soc. Argent. Bot . 21: 153-185. [ Links ]
BONZANI, N. E., E. M. FILIPPA & G. E. BARBOZA. 1997. Particularidades epidérmicas en algunas especies de Verbenaceae. Anales Inst. Biol. Univ. Nac. Autón. México, Ser. Bot. 68: 47-56. [ Links ]
BONZANI, N. E., E. M. FILIPPA & G. E. BARBOZA. 2003. Estudio anatómico comparativo de tallo en algunas especies de Verbenaceae. Anales Inst. Biol. Univ. Nac. Autón. México, Ser. Bot. 74: 31-45. [ Links ]
BRAVO, S. J. & A. GRAU. 1992. Variaciones de la densidad estomática en poblaciones de Alnus acuminata en un gradiente altitudinal. Lilloa 38: 39-45. [ Links ]
BRUNO, G., M. T. COSA & N. DOTTORI. 1999. Ontogenia de tricomas estrellados en Solanum elaeagnifolium (Solanaceae). Kurtziana 27(1): 169-172. [ Links ]
BURKART, A. 1952. Las Leguminosas Argentinas. 2 nd . ed., Buenos Aires. [ Links ]
BURKART, A. 1969. En: A. BURKART (ed.), Flora ilustrada de Entre Ríos. Vol. 3: 182-270. Colección Científica INTA, Buenos Aires. [ Links ]
CABRERA, A. L. 1976. Regiones ftogeográficas argentinas. Enc. Argent. Agricul. y Jard. 2 nd . ed. T 2 (1): 1-85, Acme, Buenos Aires. [ Links ]
CABRERA, A. L. & A. WILLINK. 1973. Biogeografía de América Latina . Programa regional de desarrollo científico y tecnológico. OEA, Washington, D.C. [ Links ]
CARRIZO, J. & S. ISASMENDI. 1994. Buddlejaceae . Flora del valle de Lerma (Novara, ed.). Aportes botánicos de Salta. Ser. Flora, Vol. 2(21): 1-10. Facultad de Ciencias Naturales, Universidad Nacional de Salta, Argentina. [ Links ]
CASTRO, M. T., S. M. CARPANO, E. D. SPEGAZZINI & M. T. NÁJERA. 2001. Leaf micrographic parameters of Terminalia australis Cambessedes (Combretaceae). Boll. Mus. Reg. Sci. Nat. Torino 18: 345-354. [ Links ]
CLAVIN, M., K. LORENZEN, A. MAYER, V. MARTINO & T. ANKE. 1999. Biological activities in medicinal species of Eupatorium . Acta Hort . 501: 277-282. [ Links ]
COLARES, M. N., N. D. BAYÓN, S. A. STENGLEIN & A. M. ARAMBARRI. 1999. Anatomía y etnobotánica de las especies medicinales de la Provincia Pampeana : Solanaceae (excepto Grabowskia y Solanum ). Acta Farm. Bonaerense 18: 171-182. [ Links ]
CORTADI, A. A. & M. A. GATTUSO. 1994. Caracterización anatómica e histoquímica de Eupatorium macrocephalum Less., E. inulaefolium H.B.K. y E. subhastatum Hook. et Arn. (Asteraceae). Dominguezia 11:32-42. [ Links ]
CORTADI, A., O. DI SAPIO & M. GATTUSO. 1996. Caracteres anatómicos de tres especies medicinales de la familia Myrtaceae. Acta Farm. Bonaerense 15: 109-123. [ Links ]
COSA DE GASTIAZORO, M. T. 1991. Estudio morfoanatómico de órganos vegetativos en Cestroideae (Solanaceae) I: tribu Nicotianeae . Kurtziana 21: 111-152. [ Links ]
D'AMBROGIO DE ARGÜESO, A. 1986. Manual de técnicas en histología vegetal . Ed. Hemisferio Sur, Buenos Aires. [ Links ]
DIMITRI, M. J. 1975. Las áreas argentinas de bosques espontáneos . En: D. Cozzo, (dir.). Enc. Argent. Agricul. y Jard. , 2 nd . ed. T 2, fasc. 16: 7-17. [ Links ]
DITSCH, F., H. PATHA & W. BARTHLOTT. 1995. Micromorphology of epicuticular waxes in Fabales s.l. and its systematic significance. Beitr. Biol. Pflanz. 68: 297-310. [ Links ]
DIZEO DE STRITTMATTER, C. 1973. Nueva técnica de diafanización. Bol. Soc. Argent. Bot. 15: 126-129. [ Links ]
FREEMAN, H. E. 1984. Leaf histology-two modern methods. J. Biol. Educ. 18: 271-272. [ Links ]
FREIRE, S. E. & E. URTUBEY. 1999. Compuestas medicinales de la provincia biogeográfica pampeana. Claves para su determinación e iconografías. Parte II. Compuestas con capítulos isomorfos y papus piloso (grupo 3). Acta Farm. Bonaerense 18: 283-294. [ Links ]
FREIRE, S. E., A. M. ARAMBARRI, N. D. BAYÓN, G. SANCHO, E. URTUBEY, C. MONTI, M. C. NOVOA & M. N. COLARES. 2005. Epidermal characteristics of toxic plants for cattle from the Salado River Basin (Buenos Aires, Argentina). Bol. Soc. Argent. Bot . 40: 241-281. [ Links ]
FRYNS-CLAESSENS, E. & W. VAN COTTHEM. 1973. A new classification of the ontogenetic types of stomata. Bot. Rev . 39: 71-138. [ Links ]
GATTUSO, M. A. 1996. Estudio anatómico, ultraestructural y fitoquímico de las Phytolaccaceae de la Argentina. Ph.D. tesis, Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario. Inédito. [ Links ]
GONZÁLEZ, M., A. LOMBARDO & A. J. VALLARINO. 1939 . Plantas de la medicina vulgar de Uruguay . Talleres Gráficos S.A. Cerrito 580, Uruguay. [ Links ]
GUANTAY, M. E. 2004. Morfoanatomía y arquitectura foliar de Blepharocalyx salicifolius (Kunth) O. Berg (Myrtaceae). Lilloa 41: 85-92. [ Links ]
HELWIG, F. H. 1992. Untersuchungen zur Behaarung ausgewählter Astereae (Compositae). Studies on the hairs of some Asteraceae (Compositae). Flora 186: 425-444. [ Links ]
HOLMGREN, P. K., N. H. HOLMGREN & L. C. BARNETT. 1990. Index herbariorum. Part 1: The Herbaria of the World. 8th ed. New York Botanical Garden, Bronx. [ Links ]
HIERONYMUS, J. 1882. Plantae diaphoricas florae argentinae. Bol. Acad. Cienc . Córdoba 4: 199-598. [ Links ]
HURRELL, J. A. & D. H. BAZZANO. 2003. Arbustos 1. Nativos y exóticos . Colección biota rioplatense 8. Ed. L.O.L.A. (Literature of Latin America), Buenos Aires. [ Links ]
KALESNIK, F. A. & A. I. MALVÁREZ. 1996. Uso antrópico potencial de las especies vegetales nativas y naturalizadas del bajo Delta del Río Paraná, Argentina. Vida Silvestre Neotropical 5: 12-21. [ Links ]
LAHITTE, H. B. & J. A. HURRELL. 1994. Los árboles de la Isla Martín García . Comisión de Investigaciones Científicas (CIC). Buenos Aires. [ Links ]
LAHITTE, H. B. & J. A. HURRELL. 1995. Las plantas de la medicina popular de la Isla Martín García (nativas y naturalizadas) . Comisión de Investigaciones Científicas (CIC). Buenos Aires. [ Links ]
LAHITTE, H. B., J. A. HURRELL, M. J. BELGRANO, L. JANKOWSKI, P. HALOUA & K. MEHLTRETER. 1998. Plantas medicinales rioplatenses . Ed. L.O.L.A. (Literature of Latin America), Buenos Aires. [ Links ]
LAHITTE, H. B. & J. A. HURRELL. 1999. Árboles rioplatenses . Ed. L.O.L.A. (Literature of Latin America), Buenos Aires. [ Links ]
LEONARDI, D., O. DI SAPIO, M. GATTUSO & S. GATTUSO. 2002. Caracteres morfoanatómicos de la corteza y hojas de Tabebuia impetiginosa y T. heptaphylla (Bignoniaceae). Bol. Soc. Argent. Bot. 37: 51-61. [ Links ]
LERSTEN, N. R & J. D. CURTIS. 1986. Tubular cavities in white snakeroot, Eupatorium rugosum (Asteraceae). Amer. J. Bot. 73: 1016-1021. [ Links ]
LISCOVSKY, I. J. & M. T. COSA. 2005. Anatomía comparada de hoja y tallo en los representantes de Cestreae G. Don (Solanaceae) de Argentina. Gayana Bot . 62: 33-43. [ Links ]
LORCA, G. G., A. G. AMAT & C. GONZÁLEZ. 1995. Análisis comparativo de caracteres diagnósticos para la identificación de tres especies argentinas de Myrtaceae empleadas en la medicina popular. Acta Farm. Bonaerense 14: 81-86. [ Links ]
MANDRILE, E. L. 2005. Plantas medicinales que se dispensan en Argentina . Tomo 1. (2da edición). Colegio de Farmacéuticos de la Provincia de Buenos Aires. [ Links ]
MANTESE, A. I. & N. H. MONTALDO. 2002. Contribución a la anatomía foliar de Blepharocalyx salicifolius (Myrtaceae). Bol. Soc. Argent. Bot . 37: 167-170. [ Links ]
MARTÍN, C., M. PARRADO & M. ARIAS. 2001. Anatomía de hoja y fruto de Cestrum strigillatum Ruíz et Pav. (Solanaceae). Lilloa 40: 215-222. [ Links ]
MARTÍNEZ CROVETTO, R. 1963. Esquema fitogeográfico de la provincia de Misiones (República Argentina). Bonplandia 1: 171-223. [ Links ]
MARTÍNEZ CROVETTO, R. 1964. Estudios etnobotánicos. I. Nombres de plantas y su utilidad, según los indios Tobas del este del Chaco. Bonplandia 1: 279-333. [ Links ]
MARTÍNEZ CROVETTO, R. 1965. Estudios etnobotánicos. II. Nombres de plantas y su utilidad, según los indios Vilelas del Chaco. Bonplandia 2: 1-23. [ Links ]
MARTÍNEZ CROVETTO, R. 1967.Tintorería Toba. Bonplandia 2: 103-105. [ Links ]
MARTÍNEZ CROVETTO, R. 1981. Las plantas utilizadas en medicina popular en el noroeste de Corrientes. Fundación Miguel Lillo. Miscelánea 69: 69-70. [ Links ]
METCALFE, C. R. 1987. Anatomy of the Dicotyledons . Vol. 3. Clarendon Press, Oxford. [ Links ]
METCALFE, C. R. & L. CHALK. 1950. Anatomy of the Dicotyledons . Vol. 1 and 2. Clarendon Press, Oxford. [ Links ]
METCALFE, C. R. & L. CHALK. 1979. Anatomy of the Dicotyledons . 2 nd . ed. Vol. 1. Clarendon Press, Oxford. [ Links ]
METCALFE, C. R. & L. CHALK. 1989. Anatomy of the Dicotyledons . 2 nd . ed. Vol. 2. Clarendon Press, Oxford. [ Links ]
PAZ, E. A., M. J. BASSAGODA & F. FERREIRA. 1992. Yuyos-uso racional de las plantas medicinales. Fin de siglo, Montevideo. [ Links ]
PERTUSI, L. A. 1987. Caracteres foliares de especies de Baccharis (Compositae) tóxicas para el ganado de la cuenca del arroyo Sauce Corto (Partido de Coronel Suárez, Provincia de Buenos Aires). Revista Mus. La Plata 93: 119-191. [ Links ]
PERROTTA, V. G. & A. M. ARAMBARRI. 2004. Schinus longifolia var. longifolia (Anacardiaceae): anatomía foliar y caulinar. Acta Farm. Bonaerense 23: 142-147. [ Links ]
PONESSA, G. I., A. M. ZENOFF, M. F. PARRADO & H. MORENO. 1998. Morfoanatomía y composición lipídica en hojas de Sapindus saponaria L. (Sapindaceae) y Tipuana tipu (Benth.) O. Kunth. (Leguminosae). Lilloa 39: 137-146. [ Links ]
POOLE, I., J. D. B. WEYERS, T. LAWSON & J. A. RAVEN. 1996. Variations in stomatal density and index: implications for palaeoclimatic reconstructions. Plant, Cell Environ. 19: 705-712. [ Links ]
RAGONESE, A. E. & V. A. MILANO. 1984. Vegetales y substancias tóxicas de la flora argentina. Enc. Argent. Agricul. y Jard. , fasc. 8. Acme, Buenos Aires. [ Links ]
RATERA, E. & M. RATERA. 1980. Plantas de la flora argentina empleadas en medicina popular . Ed. Hemisferio Sur, Buenos Aires. [ Links ]
RIGONATTO, O., N. DOTTORI & M. T. COSA. 2005. Anatomía en Solanum argentinum (Solanaceae). Kurtziana 31(1-2): 21-28. [ Links ]
RONDINA R. V. D., A. L. BANDONI & J. D. COUSSIO (eds.). 2003. Plantas silvestres argentinas con reconocidas propiedades medicinales o tóxicas . Base de Datos, CYTED-OEA. [ Links ]
ROTH, I. 1984. Stratification of tropical forests as seen in leaf structure . Junk Publ., The Hague, Boston Lancaster. [ Links ]
ROTH, I. 1990. Leaf structure of a Venezuela cloud forest (in relation to the microclimate). In : I. ROTH (ed.), Encyclop. Plt. Anat ., Vol 14, Part 1, Gebrüder Borntraeger, Berlin. [ Links ]
ROTH, I. 1995. Leaf structure: montane regions of Venezuela with an excursion into Argentina. In : I. ROTH (ed.), Encyclop. Plt. Anat ., Vol 14, Part 3, Gebrüder Borntraeger, Berlin. [ Links ]
ROTH, I. 1996. Microscopic venation patterns of leaves and their importance in the distinction of (tropical) species. In : I. ROTH (ed.), Encyclop. Plt. Anat ., Vol 14, Part 4, Gebrüder Borntraeger, Berlin. [ Links ]
SALISBURY, E. 1927. On the causes and ecological significance of stomatal frequency with special reference to the woodland flora. Phil. Trans. R. Soc. Lond ., Ser B. 216: 1-65. [ Links ]
SCHULZ, A. G. 1997. Algunas plantas usuales del nordeste argentino. Parodiana 10: 211-241. [ Links ]
SELVI, F. & M. BIGAZZI. 2001. Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomy. Flora 196: 269-285. [ Links ]
SINCLAIR, C. B. & G. K. SHARMA. 1971. Epidermal and cuticular studies of leaves. J. Tenn. Acad. Sci . 46: 2-11. [ Links ]
SORARÚ, S. & A. BANDONI. 1978. Plantas de la medicina popular argentina . Ed. Albatros, Buenos Aires. [ Links ]
SPEGAZZINI, E. D., M. T. CASTRO, J. ROSSI, S. M. CARPANO & M. T. NÁJERA. 2002. Micrografía foliar cuali-cuantitativa de las especies de Pilocarpus (Rutaceae): P. jaborandi Holmes, P. microphyllus Stape ex Wardleworth y P. pennatifolius Lem. Dominguezia 18: 5-16. [ Links ]
STACE, C. A. 1965. Cuticular studies as an aid to plant anatomy. Bull. Br. Mus. (Nat. Hist.) Bot. 4: 1-78. [ Links ]
STENGLEIN, S. A. 2001. Características epidérmicas de la hoja de las especies medicinales del género Solanum L. (Solanaceae) de la provincia biogeográfica pampeana. Acta Farm. Bonaerense 20: 265-274. [ Links ]
STENGLEIN, S. A., A. M. ARAMBARRI, M. N. COLARES, M. C. NOVOA & C. E. VIZCAÍNO. 2003. Leaf epidermal characteristics of Lotus subgenus Acmispon (Fabaceae: Loteae) and a numerical taxonomic evaluation. Can. J. Bot. 81: 933-944. [ Links ]
STENGLEIN, S. A., A. M. ARAMBARRI, M. C. MENÉNDEZ SEVILLANO & P. A. BALATTI. 2005. Leaf epidermal characters related with plant's passive resistance to pathogens vary among accessions of wild beans Phaseolus vulgaris var. aborigineus (Leguminosae-Phaseoleae). Flora 200: 285-295. [ Links ]
SUÁREZ, G. M., A. M. C. RUFFINO, M. E. ARIAS & P. L. ALBORNOZ. 2004. Anatomía de hoja, fruto y semilla de Cupania vernalis (Sapindaceae), especie de importancia en frugivoría. Lilloa 41: 57-69. [ Links ]
TOURSARKISSIAN, A. 1980. Plantas medicinales de la Argentina . Ed. Hemisferio Sur, Buenos Aires. [ Links ]
VARELA, B. G. & A. A. GURNI. 2003. Análisis micrográfico de dos hemiparásitas argentinas usadas en medicina popular y su aplicación en el control de calidad. Acta Farm. Bonaerense 22: 42-552. [ Links ]
VERVOORST, F. 1979. La vegetación del noroeste argentino y su degradación . Serie Conservación de la naturaleza Nº 1, Fundación Miguel Lillo. Tucumán. [ Links ]
WOODWARD, F. I. & F. A. BAZZAZ. 1988. The responses of stomatal density to CO 2 partial pressure . J. Exp. Bot . 39: 1771-1778. [ Links ]
WAGNER, M. L. & G. I. PONESSA. 2004. Morfología foliar y seminal de Fagara coco (Gill.) Engler (Rutaceae). Lilloa 41: 71-84. [ Links ]
YAJÍA, M. E., D. A. MARTÍ, C. J. BIDAU, A. G. AMAT, A. G. RIGLOS & A. SILVESTRONI. 1999. Genotoxicity evaluation of Allophylus edulis (Camb) Radlk. (Sapindaceae) aqueous extract. Acta Hort . 501: 31-35. Belgium. [ Links ]
ZULOAGA, F. & O. MORRONE (eds.). 1999. Catálogo de las plantas vasculares de la República Argentina. II. Dicotyledoneae. Monogr. Syst. Bot. Missouri Bot Gard . 74: 1-1269. [ Links ]
Recibido el 31 de Marzo de 2006,
aceptado el 07 de Noviembre de 2006.

