










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.42 n.3-4 Córdoba ago./dic. 2007
La vegetación de los Altos Andes II. Las Vegas del flanco oriental del Cordón del Plata (Mendoza, Argentina)
Eduardo Méndez1
1UID Botánica y fitosociología. IADIZA-CRICYT-CONICET. Avda Dr. Adrián Ruiz Leal, s/nº|. Parque General San Martín, CC 507, (5500) Ciudad, Mendoza, Argentina. E-Mail: emendez@lab.cricyt.edu.ar
Resumen: Para completar estudios de vegetación de la vertiente oriental del Cordón del Plata, se analizaron las comunidades vegetales de sitios húmedos de vegas. Se determinaron dos tipos de vegas: de baja altura, entre los 1200- 2000 m s.m. y de altura, por encima de los 2000 m s.m. representadas por 9 y 11 comunidades respectivamente. Estas se describieron fisonómica, florística y ecológicamente. Se establecieron relaciones de distribuciones de las vegas entre altitudes y temperaturas, involucrando también a procesos de congelamiento de los suelos. Además, se consideraron aspectos biológicos, de origen y corológica de las especies. En coincidencia con los pisos de vegetación del área sobre las vegas de altura se reconocen tentativamente una de mediana altura distribuída entre los 2000 a 3200 m s. m. y una de tundra que no fue considerada en este trabajo , por encima de 3200 m s.m.. Se elaboró un mapa de localización de las vegas del área de estudio.
Palabras clave: Altos Andes; Cordón del Plata; Vegetación de las vegas; Pisos de vegetación; Fitogeografía.
Summary: Vegetation of the high Andes II. The meadows on the eastern slope of the Cordón del Plata (Mendoza, Argentina). In this study I present a description of the plant communities of humid sites of vegas or mallines of the eastern slope of the Cordón del Plata were analyzed. Two types of vegas were identified: 1) low altitude vegas between 1200- 2000 m a.s.l. represented by 9 communities; and 2) high altitude vegas above 2000 m a.s.l. represented by 11 communities. These communities were described through their physiognomy, floristic composition, and ecology. In order to analize the distribution of the vegas, relationships were drawn among altitude, temperature, and soil freezing processes. In addition, biological and chorological characteristics of the species, as well as their origins were considered. With respect to the vegetation distribution, high-altitude vegas occur at a intermediate altitudes between 2000 and 3,200 meters above sea level, and tundra (which was not considered as part of the study) occurs above 3,200 meter above sea level. A map was produced of the location of the area's wetlands.
Key words: High Andes; Cordón del Plata; Vegetation of the vegas; Vegetation belts; Phytogeography.
INTRODUCCIÓN
Estudios de flora y vegetación de sitios húmedos en las zonas montañosas de Sudamérica han sido considerados en ambientes de páramos (Cleff, 1981), puneños (Gutte, 1980; Martínez Carretero, 1997; Rivas Martínez & Tovar, 1982; Ruthsatz, 1995, 2000) patagónicos ( Gandullo & Faggi, 2003, 2006; Méndez, 2005; Roig et al, 1985) y, más raramente, andino central (Hauman, 1918; Boecher et al., 1963,1968,1972; González Loyarte & Peralta, 2004; Méndez, 1986, 2004; Méndez et al. 2006). A pesar de la valiosa información que se tiene de la vegetación de los sitios húmedos en la zona andina central, donde regionalmente se los identifica como vegas (o mallines), éstos no son suficientemente conocidos. En estos sitios húmedos factores ambientales como la temperatura jugarían un papel importante en el establecimiento y distribución de las especies de plantas y sus comunidades. En efecto, la temperatura podría delimitar muy bien no sólo el ascenso o descenso altitudinal de las mismas, sino también su distribución latitudinal. Por esta razón, y para complementar estudios anteriores de vegetación del área del Cordón del Plata (Andes Centrales) (Méndez, 2004), aquí se pretende abordarlos en su desarrollo altitudinal. Según hipótesis, se hallarían, en el desarrollo altitudinal, a distintas comunidades vegetales de vegas que podrían agruparse en diferentes tipos o clases a modo de pisos altitudinales. Se esperaría encontrar, en estas clases o tipos de vegas, a especies y comunidades vegetales afines a las halladas en estudios de otras áreas húmedas.
Los objetivos de este trabajo fueron 1: clasificar y describir las comunidades vegetales de vegas que se desarrollan en un gradiente altitudinal en el Cordón del Plata (Andes Centrales), 2: relacionarlas con factores ecológicos como la temperatura que pueden influir en su distribución.
MATERIALES y MÉTODOS
1. El área estudiada
Se localiza al oeste de la ciudad de Mendoza, entre 69º 10' y 69º 24' W y 32º 54' a 33º 04' S y entre los 1200 y 5000 m s.n.m. aproximadamente (Fig. 1). La superficie total del área estudiada es de aproximadamente 30.000 ha . Las características climáticas, orográficas, geológicas, hidrográficas y de uso del área ya han sido consideradas en un trabajo anterior (Méndez, 2004). Las diferencias de altitudes, pendientes y exposiciones del relieve determinan fuertes variaciones en las temperaturas y precipitaciones. Mientras los sectores mas bajos presentan condiciones de mayor aridez, con temperaturas más altas, y sequías, los de mayor altitud son más fríos y húmedos. Geológicamente existen formaciones precámbricas, paleozoicas, pérmicas, terciarias y cuaternarias que afloran en sectores de los relieves geomorfológicos de la Cordillera Frontal , Precordillera y niveles de glacis (Caminos, 1965, 1979, Mikkan, 1998, Viers, 1964). Las texturas de los suelos varían desde muy gruesas en alta montaña, medianas en los niveles de glacís y cerrilladas, hasta muy finas en los embanques aluvionales de los ríos.

Fig. 1. Localización de las vegas en el flanco oriental del Cordón del Plata, Luján de Cuyo, Mendoza, Argentina. Sitios de vegas de baja altura ( 1-8 ): 1: ciénagas de Potrerillos, 2: Médanos de Cacheuta, 3: Cerros Colorados, 4: Quebrada del 60 y del Medio, 5: Cerro Médanos, 6: Quebrada de la Manga , 7: El Mal Paso, 8: Arroyo Blanco; de altura (9-20): 9: Agua de Juancho, 10: Quebrada de la Manga-El Salto , 11: Quebrada del Chacay, 12: Quebrada de los Manantiales, 13: Quebrada del Andresito, 14: Morenas Coloradas, 15: Rincón de Vallecitos, 16: Rincón de los Colorados, 17: Cerro Negro, 18: Ciénagas del Bayo, 19: Quebrada de los Potrerillos y 20: Quebrada de la Hoyada. El mapa muestra aspectos geomorfológicos del área (Viers, 1964, Mikkan, 1988). Las letras minúsculas ( a-j ) indican las localidades con datos meteorológicos.
Las vegas del área cubren pequeñas superficies de diferentes formas y tamaños, localizadas preferentemente en los bordes de los cauces con agua permanente y en las surgencias naturales de las laderas. Prácticamente todas están sometidas en mayor o menor grado a la actividad ganadera extensiva con caprinos, vacunos y equinos.
2. Análisis de la vegetación
Se utilizó el método fitosociológico de la escuela de Zürich-Montpellier (Braun-Blanquet, 1979). Los relevamientos fueron obtenidos durante los años 1986 a 1992 utilizando en cada comunidad parcelas de 0,50 a 25 m2 ubicadas en sitios florística, fisonómica y ecológicamente homogéneos, tratando de cubrir la totalidad de las comunidades. Todos los relevamientos obtenidos (102) fueron volcados a una tabla general donde las especies se ordenaron por los valores de cobertura o abundancia-dominancia de la escala de Braun- Blanquet. A través de las especies diferenciales se determinaron las comunidades vegetales. Paralelamente se analizaron, sobre la base de la lista florística, las formas de vida y espectros biológicos según Raunkiaer (1934), y se identificó la corología y el origen de cada especie utilizando la información disponible sobre la mismas de Boecher et al (1963, 1968, 1972), Cabrera (1976), Hauman (1918), Roig (1998), Zuloaga et al. (1994) y Zuloaga & Morrone (1996, 1999a, 1999b). De la tabla general de relevamientos se obtuvo una tabla sintética de las comunidades expresando los valores de cada especie por sus grados de constancia o presencia ( números romanos en Tabla 1). También para la descripción de cada comunidad se elaboraron tablas parciales solo con los valores de abundancia o cobertura (números arábigos en Tablas 2 a 9). No se trató la sintaxonomía de las comunidades por no contar con suficiente información de otras áreas o de regiones más amplias a ésta.
El análisis ecológico involucró las relaciones entre la distribución de las vegas en altitud y las temperaturas en el sector analizado utilizando los valores de temperaturas medias anuales, de los meses mas frío (julio ) y más cálido (enero) para localidades dentro del área o próximas a ella (De Fina et al., 1964). Con ellos se obtuvo una línea de regresión entre esos valores y la altimetría. Este procedimiento (Roig et al., 2007), permite obtener ideas de las condiciones de temperaturas que existen en cada una de las clases de vegas y hasta poder relacionarlas con las áreas parageocriogénicas y geocriogénicas (Corte, 1983a,b; Minetti & Corte, 1984.).
Con los valores de temperaturas obtenidos en cada caso , y considerando un gradiente medio de -0.6 ºC por cada 100 m de ascenso (Buk, 1983) se determinaron los valores de temperaturas de los límites inferior y superior de las vegas.
Para la identificación correcta de los nombres de las especies fueron usados los catálogos de las plantas vasculares de Zuloaga et al. (1994) y Zuloaga & Morrone (1996, 1999a, 1999b).
Para la labor cartográfica de localización de las vegas se contó con el apoyo de las fotografías aéreas de pares y mosaicos a escala 1:50.000 de la Dirección General de Fabricaciones Militares, más el de imágenes satelitarias Landsat disponibles a escala 1:250.000 aproximadamente. Con el análisis de la vegetación de las vegas en las tablas comparativas y el control permanente del terreno, se representaron perfiles transversales de las comunidades vegetales.
RESULTADOS y DISCUSIÓN
La vegetación
La vegetación del área se distribuye altitudinalmente en pisos representados por los subdistritos de Larrea cuneifolia (1200- 1750 m s.n.m.), Larrea divaricata (1300- 1950 m s.n.m.), Junellia scoparia- Colliguaja integerrima (1500- 2250 m s.n.m.), Adesmia horrida (2100- 3200 m s.n.m.), Adesmia subterranea-Azorella monantha (2750- 3700 m s.n.m), y Nassauvia cumingii - Senecio chrithmoides ( 3500- 4200 m s.n.m.), cada uno caracterizado por su fisonomía, composición florística y condiciones ecológicas (Méndez, 2004).
Por análisis de las especies características y diferenciales se han determinado dos tipos de vegas: de baja altura ( 1000 a 2000 m s.n.m.) y de altura (> 2200 m s.n.m.) que están representadas por 9 y 11 comunidades respectivamente y que revelan la heterogeneidad y la presencia de biotopos distintos. En el piso superior no se ha contemplado, por el momento, un tercer tipo de vega que se correspondería con la tundra andina por encima de los 3200 m s.n.m. Es posible que un examen mas profundo de las vegas en estas y otras áreas permitan ajustar mejor sus límites con los de los principales pisos vegetales.
Descripción de las comunidades
I. Vegas de baja altura (grupo 1 a 9 en Tabla 1)
Tabla 1. Comunidades vegetales de las vegas del flanco oriental del Cordón del Plata, Mendoza, Argentina. Números romanos y arábigos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80). (Ver tabla)
Comprende aquellas vegas originadas en los cursos de agua temporarios a mas permanentes entre los 1200 a 2000 m s.n.m. aproximadamente. Se localiza en los pisos más cálidos y secos de los jarillales de Larrea spp. Por lo general son salinas, como las situadas en las vertientes de los cauces huayquerianos de los relieves Terciarios y Triásicos. Si bien son numerosas, por lo común ocupan pequeñas superficies solo incrementadas en márgenes del río principal.
En la Tabla 1 se ordenan las comunidades, de ambos tipos de vegas, en un gradiente de xericidad y salinidad en aumento hacia las últimas columnas. En la Figura 2 se da un esquema de la distribución de las comunidades vegetales de las vegas de baja altura.

Fig. 2. Distribución esquemática de las vegas de baja altura. Zonificación de las comunidades vegetales: 1: Azolla filiculoides , 2: Typha domingensis , 3: Phragmites australis , 4: Veronica anagallis-aquactica , 5: Rorippa nasturtium aquacticum , 6: Schoenoplectus pungens , 7: Juncus articus , 8: Cortaderia rudiuscula y 9: Tamarix gallica . Ciénagas de Potrerillos, Río Mendoza y Las Vegas, Luján de Cuyo, Mendoza, Argentina.
1- Comunidad de Azolla filiculoides (Tabla 1, Tabla 2).
Tabla 2. Comunidades de Azolla filiculoides ,de Typha domingensis y de Phragmites australis . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Se presenta en sitios con agua con escaso movimiento o casi estancada. Allí domina en densas colonias Azolla filiculoides y con frecuencia la acompañan Lemna gibba y Myriophyllum elatinoides que en conjunto pueden llegar a ocultarla. Esta comunidad de flotantes se contacta con la de Typha domingensis.
2- Comunidad de Typha domingensis (Tabla 1, Tabla 2)
Esta unidad se registra no solo en el lecho del río Mendoza donde adquiere la mayor expresión vegetativa sino también en pequeños sectores de los cauces en el relieve huayqueriano de las Crestas del Toscal, donde cubre áreas pantanosas o con muy lento desplazamiento de las aguas, poco profundas. En uno de estos sectores se halla Habenaria paucifolia , orquidea de pantano, de fuerte presencia en colonias al pie de los totorales. También se la encuentra en habitats similares, en Potrerillos, sobre las terrazas aluviales del río Mendoza.
La fisonomía de la unidad es la de un totoral de hasta 2 m de alto con máximas coberturas.
Además de la especie dominante se comportan como características diferenciales Habenaria paucifolia , Xanthium cavanilliesii , Salix alba var. alba. En este ambiente salino, con agua estancada y con suelos fangosos y agua en superficie dominan el estrato inferior Schoenoplectus pungens, Hydrocotyle bonariensis , y Eleocharis melanomphala . Typha domingensis es cosmopolita con un amplio espectro geográfico.
En el área esta comunidad ha sido muy castigada y en algunos sectores se ha mantenido hasta el endicamiento del dique Potrerillos.
3- Comunidad de Phragmites australis (Tabla 1, Tabla 2).
Se destaca no solo en las áreas pantanosas de Typha , en el lecho del río Mendoza, sino también en las vertientes de laderas próximas a la estación Guido, en contacto frecuente con el matorral -pajonal de Cortaderia rudiuscula . En general ocupa pequeñas superficies de sitios húmedos y salinos. No es raro que figuren con ella elementos de la vegetación flotante y de las comunidades de Schoenoplectus pungens , sobre todo cuando existen claros por su extracción. Esto último ha llevado a la reducción de sus superficies. Tiene como característica diferencial a Fimbristylis autumnalis. Esta comunidad se asemeja a la de la Reserva Provincial Laguna de Llancanelo (Méndez, 2005).
4-Comunidad de Rorippa nasturtium-aquacticum (Tabla 1, Tabla 3).
Tabla 3. Comunidades de Rorippa nasturtium-aquacticum y de Veronica anagallis - aquactica . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Esta unidad aparece como etapa dinámica en los ambientes muy húmedos de los arroyos de modo transitorio en los cauces temporarios o más permanentes, aunque en ellos requiere agua superficial con desplazamiento más o menos lento y figura como cinturón perisférico a ella cuando adquiere mayor velocidad. Convive con Veronica anagallis-aquactica de semejante distribución. En ella se observan elementos de esta y de comunidades flotantes. A medida que se estanca y disminuye el nivel del agua ingresan elementos higrófilos como Mentha aquactica , Hydrocotyle bonariensis , Polypogon monspeliensis , etc.
5- Comunidad de Veronica anagallis-aquatica (Tabla 1, Tabla 3).
Está muy emparentada con la anterior, con la que se contacta con frecuencia; resulta sin embargo más tolerante a la falta de agua superficial. En éstas últimas es posible hallar a Juncus septati , Poa annua , Plantago major, Ranunculus cymbalaria , Sisyrinchium sp., etc. que denuncian las aguas estancadas. Chara fragilis logra formar colonias en sitios con lento movimiento de las aguas.
Rorippa nasturtium-aquacticum y Veronica anagallis-aquatica, de origen holártico, han sido citadas en borde de cauces de acequias y riachuelos en Perú (Galán de Mera et al., 2003).
6- Comunidad de Schoenoplectus pungens (Tabla 1, Tabla 4).
Tabla 4. Comunidades de Schoenoplectus pungens y de Juncus articus . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos y arábigos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Esta comunidad se localiza en sitios semejantes a los anteriores con los que se contacta. Con frecuencia se asocia a suelos arcillosos empapados en agua o con agua salinas en lento movimiento.
Su fisonomía es la de una pradera húmeda de ciperáceas de 20 -30 cm de alto y con máximas coberturas.
Tiene como plantas características a Eleocharis melanomphala , E. albibracteata , Ranunculus cymbalaria , Polypogon monspeliensis , etc. comportándose como preferenciales. Esta unidad permite el ingreso de las especies mas higrófilas y salinas y las más salinas y menos húmedas. Esto sucede con Juncus articus que incorpora con su ingreso a elementos del Cortaderal.
En esta comunidad Schoenoplectus pungens ha sido citado como elemento dominante de vegas cálidas en San Juan (Gonzalez Loyarte & Peralta, 2004).
7- Comunidad de Juncus articus (Tabla 1, Tabla 4).
Se halla muy emparentada con la anterior, a la que desplaza cuando descienden los niveles de agua. Esta unidad no se ha observado por el momento bien desarrollada en la zona.
Es una comunidad de amplia extensión en bañados de Mendoza (Méndez, 2005), en la zona andina (González Loyarte & Peralta, 2004; Méndez et al., 2006) y Patagonia (Gandullo & Faggi, 2006).
8- Comunidad de Cortaderia rudiuscula (Tabla 1, Tabla 5).
Tabla 5. Comunidades de Cortaderia rudiuscula y de Tamarix gallica . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80). (Ver tabla)
Está bien representada en los embanques aluviales del río Mendoza, en ambas márgenes , destacándose en proximidades del Hotel Potrerillos y en fajas mas o menos discontinuas a todo lo largo del río. También de igual modo aparece en los cauces temporarios y salinos de las áreas huayquerianas donde logra ascender por los cursos permanentes de agua como los del arroyo Blanco, de la Manga , y penetrar en el piso de las vegas mas húmedas. En los sectores de la baja montaña vive en los cauces y en las vertientes de los fondos de las quebradas de los relieves triásicos o con areniscas (Ao del Rincón Verde, del Durazno, etc.) y en las formaciones de Mogotes donde su presencia es sinónimo de fuente de agua.
En general prefiere suelos profundos arenosos a franco arenosos y salinos.
La fisonomía es la de un pastizal alto-pajonal, cortaderal, bi a triestratificado con plantas de hasta 3 m o más de alto y máximas coberturas, en las mejores condiciones de humedad y nutrientes minerales.
Se comportan como características Tessaria absinthioides , Distichlis spicata, Baccharis spartioides , Hypochoeris chondrilloides , Muhlenbergia asperifolia , etc. que denuncian las condiciones de salinidad del lugar. Es frecuente que ingresen en la unidad elementos más xéricos de los cauces o de las comunidades vecinas como Senecio subulatus , Eupatorium patens , E. buniifolium , etc.
9- Comunidad de Tamarix gallica (Tabla 1, Tabla 5).
Esta unidad vive en sitios mas o menos salinos de los cursos de agua y se destaca en las márgenes del lecho del río Mendoza, donde logra formar bosquecitos de hasta 3- 4 m de alto. Su distribución en el lecho del río y a lo largo del mismo no es una casualidad, pues tiene alto poder de germinación sobre los embanques aluvionales donde logra formar densos almácigos, comportándose en ellos como pionero, colonizador y constructor de las nuevas unidades. Tiene frecuente contacto con el matorral de Cortaderia rudiuscula y se comportan como características preferenciales, Melilotus albus y Baccharis juncea.
También se la encuentra esporádicamente en los cauces temporarios y salinos de los relieves huayquerianos de la Cuesta del Toscal.
II. Vegas de altura (grupo 10 a 20 en Tabla 1).
Son vegas de montaña que en su ascenso entre los 2000 m s.n.m. hasta los 3200 m s.n.m. por lo general muestran un incremento de la reacción ácida. Por lo común son arenosas a franco arenosas con limos y arcillas y altos contenidos de materia orgánica. Las vegas de mayor altura aparecen al pie de las Morenas glaciarias (vegas del Rincón, de la Angostura , etc. sobre hoyadas mas o menos planas o deprimidas de las vertientes, conos y a lo largo de los cursos de aguas permanentes (Morteritos). A menor altitud y dentro del piso de Adesmia horrida se las ve en similares relieves (vegas del Bayo, de las Vacas, Potrerillos, Manantiales, etc). Muchas de ellas están gran parte del año cubiertas por la nieve, sobre todos las de mayores alturas, y solo quedan disponibles para el pastoreo en los meses estivales y, a veces, por unos pocos días del mes. Por razones climáticas existen suelos permanentemente congelados (permafrosts) localizados en las altas cumbres o bien con congelamiento estacional y estados intermedios en los glacís y faldeos inferiores de la montaña. Allí son frecuentes e intensos los procesos de crioturbación, crioclastismos, soligelifluxión, etc. A estas altitudes los fenómenos de movilización de los suelos (coladas, canaletas, etc.) son comunes e impiden la franca estabilidad de arraigamiento de las cubiertas vegetales.
En la Tabla 1 se ordena la vegetación actual conforme a la disponibilidad de agua, desde lo más húmedo a lo más seco. En la Figura 3 se da un esquema de la distribución de las comunidades vegetales de las vegas de altura.

Fig. 3 . Distribución esquemática de las vegas de altura. Zonificación de las comunidades vegetales: 1: Sciaromium sp., 2: Deyeuxia vellutina , 3: Deschampsia caespitosa , 4: Mimulus luteus , 5: Calceolaria luxurians , 6: Werneria pygmaea , 7: Plantago uniglumis , 8: Senecio bonariensis , 9: Eleocharis albibracteata, 10: Carex gayana , 11: Trifolium repens . Quebrada de los Manantiales, Ciénagas del Bayo, Quebrada del Andresito y del Rincón, Luján de Cuyo, Mendoza, Argentina.
10- Comunidad de Sciaromium sp. con algas (Tabla 1, Tabla 6).
Tabla 6. Comunidades de Sciaromium , de Deyeuxia vellutina y de Deschampsia caespitosa con Poa planifolia . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos y arábigos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Se presenta alrededor de los 2600 m s.n.m. sobre las piedras de los lechos de los arroyos y en donde el agua escurre con fuerte desplazamiento. Esta comunidad de musgos con algas es pionera y colonizadora del cauce y edificadora para las comunidades siguientes. Con preferencia aparece en los saltos o escalones de las rocas o rodados grandes que frenan la violenta corriente del agua. Una de las especies dominantes de musgo es Sciaromium sp.
Esta comunidad se ha registrado de modo fragmentario en los cauces de los valles altoandinos de la Cordillera Principal (Méndez et al., 2006).
11- Comunidad de Deyeuxia vellutina (Tabla 1, Tabla 6).
Crece en sitios con fuertes desplazamientos de las aguas donde arraiga en las grietas de los bloques dispuestos transversalmente a lo largo de los arroyos. En estas condiciones, con agua en movimiento, actúa como edificadora e incorpora a otros acompañantes tales como Festuca hieronymi , Mimulus luteus y Gentianella multicaulis . También Deyeuxia vellutina prospera con alta constancia en pantanos turbosos de altura donde forma pastizales de fuerte color amarillo oro.
Ha sido registrada en sitios de vegas mas frías altoandinas a mas de 3000- 3200 m s.n.m. (González Loyarte & Peralta, 2004; Méndez et al., 2006). También ha sido señalada en vegas puneñas a más de 4000 m s.n.m. (Martínez Carretero, 1997).
12- Comunidad de Deschampsia caespitosa (Tabla 1, Tabla 6).
Es una notable comunidad que vive en el centro del cauce sobre las comunidades anteriores a las que desplaza. Es un pastizal de 20 a 50 cm de alto y máximas coberturas en las vegas del Bayo y en los cauces cordilleranos muy húmedos y con agua en superficie sobre rocas bañadas por ellas.
13- Comunidad de Mimulus luteus (Tabla 1, Tabla 7).
Tabla 7. Comunidades de Mimulus luteus y de Calceolaria luxurians . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). Números romanos y arábigos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Se registra entre los 2400 a 3020 m s.n.m. en los arroyos cordilleranos. Ocupa el centro del cauce con agua con poco movimiento o bien lo hace como primer cinturón con aguas en fuerte movimiento. Las mayores disponibilidades de suelos orgánicos determinan la presencia de Juncus scheuzerioides , Senecio breviscapus , Rorippa nasturtium-aquacticum , Epilobium glaucum , etc. Tiene especies de comunidades anteriores y de comunidades menos húmedas, como Veronica anagallis-aquactica , Ranunculu s spp., etc.
Su presencia se destaca fácilmente durante su floración por el colorido de sus flores que señalan su distribución a lo largo de los cauces. Las plantas de Mimulus alcanzan los 40 cm de alto y posee densas coberturas, tanto que a veces ocultan el desplazamiento de las aguas.
Esta comunidad puede asemejarse a la Gunnero magellanicae- Calthetum sagittatae Gand y Faggi 03 (Gandullo & Faggi, 2006) a través de las especies de Mimulus luteus , Epilobium nivale que parecen tener amplia distribución latitudinal.
14- Comunidad de Calceolaria luxurians (Tabla 1, Tabla 7).
Se sitúa con frecuencia en las nacientes de las vertientes de laderas, con humedad permanente y salida lenta de las aguas donde logra formar densas colonias. Esta máxima cobertura impide la escasa presencia de elementos que acompañan a la especie dominante y que prosperan en los claros abiertos en ella. Esto ocurre con Valeriana aquactica , Juncus scheuzerioides , Mimulus luteus , etc. Se presentan como especies diferenciales, entre otras, Marchanthia sp, con semejante patrón que Calceolaria , Ranunculus spp., etc.
No es rara su presencia en valles cordilleranos en los bordes de cauces con corrientes de agua en lento movimiento (Méndez et al., 2006).
15- Comunidad de Werneria pygmaea (Tabla 1, Tabla 8).
Tabla 8. Comunidades de Werneria pygmaea y de Plantago uniglumis . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). . Números romanos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80).

Constituye una etapa más xérica que la de Carex gayana. Vive a orillas del agua pero en sitios de suelos mas compactados, humíferos y secos. Forma en ellos una densa carpeta de 3 a 5 cm de alto. Se enriquece con elementos que viven en la comunidad de Plantago uniglumis.
Werneria pygmaea Gillies ha sido señalada como elemento de amplia distribución y óptimo ecológico y corológico puneño peruano- boliviano (Gutte, 1980, Rivas Martinez & Tovar, 1982), puneño argentino (Ruthsatz, 1977) y hasta en el páramo colombiano (Cleef 1981).
16- Comunidad de Plantago uniglumis (Tabla 1, Tabla 8).
Se encuentra en el piso nival entre los 2700 a 3300 m s.n.m. en terrenos de suaves pendientes, cóncavos y en bordes de cursos de agua permanentes.
Representa a las turberas xéricas de altura con fuerte presencia de elementos típicamente higrófilos nivales como Colobanthus quitensis , Phylloscirpus acaulis , Gentiana sedifolia , entre otros. En ellos los suelos son mas o menos compactados y están congelados la mayor parte del año.
Por la especie dominante, Plantago uniglumis, estaría muy débilmente representada en el Azorello trifoliate- Plantaginetum uniglumatis Gand. y Faggi 03 (Gandullo & Faggi, 2006).
17- Comunidad de Senecio bonariensis (Tabla 1, Tabla 9).
Tabla 9. Comunidades de Senecio bonariensis , de Eleocharis albibracteata , de Carex gayana y de Trifolium repens . Valores de abundancia-dominancia o cobertura (signo + o números arábigos) (Braun-Blanquet, 1979) de las especies en cada comunidad. Valores de coberturas: +(<1), 1(1-10), 2(10-25), 3(25-50), 4(50-75), 5(>75). Números romanos y arábigos son valores de presencia o constancia de las especies (I<20), II(20-40), III(40-60), IV(60-80), V(>80). (Ver tabla).
Se localiza en sitios de las vegas con escaso movimiento de las aguas como en las nacientes de las vegas de los Manantiales, de las Vacas, etc. , a 2300 m s.n.m. aproximadamente. Ocupa pequeñas superficies. En aquellos sectores Senecio bonariensis alcanza alturas de 1 a 2 m y máximas coberturas. Además de Senecio bonariensis son elementos característicos diferenciales Rumex sp. e Hydrocotyle bonariensis. Esta comunidad está emparentada con la de Eleocharis albibracteata y Carex gayana a traves de especies como Veronica anagallis- aquactica y Mimulus luteus.
18- Comunidad de Eleocharis albibracteata (Tabla 1, Tabla 9).
Figura en suelos muy empapados con agua superficial o subsuperficial y a modo de cinturón perisférico con la comunidad de Mimulus luteus. Su fisonomía es la de un cesped de 3- 15 cm de alto con máximas coberturas. Representa una variante de la comunidad de Carex gayana y tiene en Polypogon monspeliensis , Pratia repens , etc., a sus especies características.
Eleocharis albibracteata tiene amplia distribución latitudinal y altitudinal como lo revelan diferentes autores (Gandullo & Faggi, 2003, Méndez, 1986, 1990 a , b, 2005). Ha sido reconocida como elemento característico de pisos altoandinos en las punas húmedas y subhúmedas del Perú, norte de Bolivia (Rivas Martínez & Tovar, 1982). También en los sitios altoandinos del Norte de Argentina (Ruthtsatz, 1977). Una comunidad similar se presenta también en los sectores más altos de las Sierras de Córdoba (Cabido, 1980).
19- Comunidad de Carex gayana (Tabla 1, Tabla 9).
Surge en suelos anegados permanentemente con agua superficial o temporalmente superficial en sitios planos y con agua estancada o con lento movimiento.
Su fisonomía es la de una pradera herbácea dominada por Carex gayana de 10 a 20 cm de alto y máximas coberturas. Del mismo modo que la anterior prospera en sitios más húmedos, se enriquece en elementos de las comunidades de contacto y la de los sitios más secos de sus perisfería. Tiene en Carex fuscula a su especie característica.
Esta comunidad se asemeja a la dada como Carico gayanae-Eleocharietum albibracteate Gand. y Faggi 03 (Gandullo & Faggi, 2006) por contener entre sus elementos a Carex gayana , Eleocharis albibracteata, etc. También ha sido encontrada a más de 4000 m s.m en sitios similares de suelos saturados en agua en las vegas puneñas de Catamarca (Martínez Carretero, 1997).
20- Comunidad de Trifolium repens (Tabla 1, Tabla 9).
Esta unidad representa una etapa de degradación de las vegas a causa de la presión de pastoreo del ganado. Con preferencia se localiza en los bordes de las comunidades anteriores de Eleoccharis , Juncus , etc. En su composición florística participan plantas exóticas como Medicago lupulina y , Poa pratensis , asociadas a suelos mas compactados.
Facies de Taraxacum officinale (Tabla 1, Tabla 9).
De amplia distribución altitudinal y altamente favorecida por su dispersión por el viento y los animales alcanza en algunos sectores notables coberturas entre los 2000 a 3200 m s.n.m. En la zona de las Mesillas de Los Morteritos codomina con el pastizal de Poa holciformis a 2840 m s.n.m.
Taraxacum officinale (achicoria) es una de las malezas más difundidas en el área, ya a fines de marzo adquiere coloración rojiza de sus hojas destacándose de la fisionomía de los pastizales, en las perisferías de las vegas, o en los posaderos de los animales. Es parte integrante de la dieta de los animales.
Espectro biológico
En general las vegas analizadas bajo este punto de vista no tienen un espectro con todas las formas biológicas (Tabla 10). Así, mientras en las vegas de baja altura dominan las geófitas y nanofanerófitas, que señalan condiciones mas cálidas; en las de altura, además de un dominio de terófitas, hemicriptófitas y geófitas, se presentan las caméfitas como otras formas de vida mejor adaptadas a condiciones más frías, denunciando con ello la tendencia hacia un fitoclima general camefítico que permite la existencia de este tipo de vegas de altura.
Tabla 10. Formas de vida de las especies vegetales en las vegas del Cordón del Plata (Luján de Cuyo, Mendoza, Argentina).

Corología
Las diferencias corológicas que existen también permiten separar bajo este punto de vista los tipos de vegas (Tabla 11). En los dos tipos de vegas dominan las especies de Sudamérica y cosmopolitas. Estas últimas son más abundantes en las vegas de baja altura y esto podría atribuirse por contener un mayor número de especies introducidas.
Tabla 11. Distribución corológica de las especies vegetales en las vegas del Cordón del Plata (Luján de Cuyo, Mendoza, Argentina).

Origen fitogeográfico
En general las especies autóctonas dominan, en presencia, las vegas de altura y las alóctonas o introducidas las de baja altura. Este mayor número de especies introducidas en las vegas de baja altura al parecer estarían asociadas a una mayor presión antrópica que allí se produce.
Relaciones ecológicas de las vegas o mallines del Cordón del Plata
Si bien el agua es un factor importante de la presencia o ausencia de éstos ambientes húmedos, y de su vegetación, la temperatura también es un fuerte condicionante en sus establecimientos. En el ascenso altitudinal la temperatura disminuye exponiendo al área considerada a un aumento del frío, de procesos de congelamiento y descongelamiento de los suelos , etc., que se traducen en cambios en las composiciones florísticas, formas de vida y condiciones ecológicas de los mismos.
a-Relación entre altitud/temperatura y distribución de las vegas (Fig. 4).
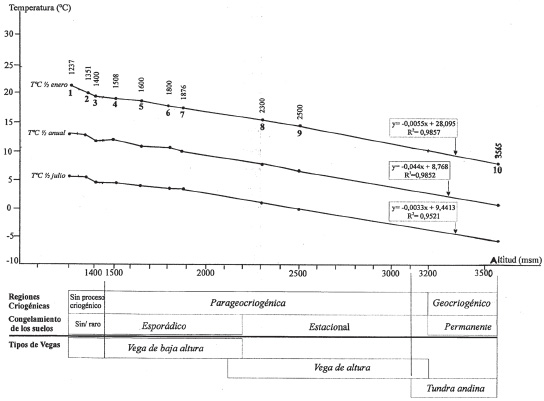
Fig. 4. Relación altura / temperatura en el flanco oriental del Cordón del Plata, Luján de Cuyo, Mendoza, Argentina. Localidades: 1: Cacheuta ( a ), 2: Potrerillos ( b ), 3: Pto Agua La Jarilla ( c ), 4: Estación Guido ( d ), 5: Pto Agua de los Bueyes ( e ), 6: El Salto I ( f ), Estancia El Plata ( g ), 7: La Hoyada ( h ), Vallecitos ( i ) y 10: El Salto II ( j ). Relaciones obtenidas sobre la base de conceptos criogénicos (Corte , 1983 a ,b , Minetti & Corte, 1984) y de vegetación (Roig et al , 2007).
Las vegas de baja altura en el flanco Oriental del Cordón del Plata se presentan en el área estudiada en las zonas sin o con escasos procesos criogénicos y en la parageocriogénica con congelamientos esporádicos, mientras las vegas de altura mas bien se localizan en las zonas parageocriogénica con congelamientos estacional (invierno) y en la región geocriogénica con congelamiento mas permanente. Hay que reconocer que el permafrost se inicia en la base de los glaciares de escombros a 3200 m s.n.m. en El Salto, y alcanza hasta los 4800 m . s.n.m., más arriba empieza el hielo descubierto (Corte 1983 a , b).
Estas relaciones entre comunidades y procesos criogénicos han sido de alguna manera documentada en vegas de la puna del norte (Martínez Carretero, 1997) donde los procesos se inician a los 3900- 4000 m s.n.m.
Las rectas de regresión dan idea de las condiciones de temperaturas existentes en los límites de vegetación para esta área De acuerdo con esto se revela a la vega de baja altura como más cálida que la de altura. En esta última es posible considerar a una de mayor frío que estaría representada por las vegas de la tundra andina por encima de los 3200 m s.n.m. y que en nuestro caso se ha considerado conjuntamente en la figura 4 atento a futuros estudios sobre su vegetación.
Siguiendo a la tipología de las cliseries termoclimáticas (Rivas- Martínez & Tovar, 1982) las vegas de baja altura (10- 13,2 ºC ) pueden ser incluídas en el grupo mesoandino inferior (10-15 º C) y la de altura (<10) dentro del mesoandino superior, (6-10) m, alto andino inferior (3-6) y hasta el altoandino superior (0-3).
b. Relación entre la latitud / temperatura y la distribución de las vegas.
Esta situación de distribución de las comunidades de vegas en altura también se observa con la latitud donde también la temperatura las condiciona Así si bien estas comunidades dadas para el Cordón del Plata están para esa latitud (32º 54' a 33º 04' lat.S) con esos límites altitudinales, estos últimos sufrirían un desplazamiento de descenso o ascenso a mayores o menores latitudes respectivamente. Esto es lo que sucede con las vegas de altura del Plata que en la provincia de Neuquén, a mayor latitud (37º 47' a 37º 55' lat. S), se encuentran con sus límites más bajos. En efecto en relación con los humedales de Neuquén las especies del piso montano inferior de este lugar (Gandullo & Faggi, 2003, 2006) están presentes en las vegas de altura del Cordón del Plata. Entre estas especies están Eleocharis albibracteata, Plantago australis, Mimulus glabratus, Alopecurus magellanicus, Plantago uniglumis, Azorella trifurcata, Deschampsia caespitosa, Carex fuscula. Esto seguramente se condice con las condiciones climáticas en la distribución de las especies y donde la temperatura debe jugar un papel importante. Podemos pensar que estas especies de sitios mas cálidos en el sur o montano inferior deben, en su desplazamiento hacia el norte y en función de sus requerimientos o tolerancias, acondicionarse a regimenes térmicos semejantes, y que en este caso en Mendoza se encontraría en sectores mas alto del flanco occidental del Cordón del Plata . Es decir que mientras los elementos florísticos y sus comunidades del sur penetran en el norte ascendiendo por otro lado los elementos de las vegas de altura de Mendoza llegan al sur descendiendo, buscando en ambos casos acondicionar sus establecimientos a los factores térmicos.
Esto también podría ayudar a explicar el porque en el sector de vegas del Cordón del Plata y aún en Portillo de Tunuyán mas al sur pero siempre en el mismo Cordón de la Cordillera Frontal no se hallan representados el piso montano superior de Neuquén dado por Gandullo y Faggi (2006) y que seguramente puedan estar a esta latitud sobre el Cordón principal o del límite Esta situación se daría posiblemente por encontrar sectores mucho mas fríos y húmedos semejantes al de los valles interiores del Aconcagua sobre el Cordón Principal o del Límite (Méndez et al., 2006 ) donde están localizadas las comunidades del piso superior de Neuquén sobre todo de Patosia o Oxyclöe , y que también se documentara sobre el límite cordillerano con Chile en el Paso el Pehuenche. Estos desplazamientos de las unidades o de las especies y sus acondicionamientos a los factores térmicos o de mayor insolación ha sido reconocido en nuestra zona para otras comunidades (González Loyarte, 1989, Martínez Carretero, 1997, Méndez, 2004, Roig, 1976, Roig & Marone, 1982).
En este análisis de distribución interesa diferenciar que no todas las especies integrantes de las comunidades reaccionan de la misma forma ante las temperaturas. Así por ejemplo existen especies criófilas que solo viven en ambientes muy fríos como por ejemplo Oxychlöe o Patosia y otras megatérmicas de sitios muy cálidos. El aumento de calor o de frío en cada caso son los factores limitantes en sus establecimientos Esto es lo que tal vez suceda con las especies y comunidades de las vegas. En alta montaña esto se ve bien en el desarrollo altitudinal de las composiciones florísticas de las comunidades vegetales y es por esta razón que el trazado de las isotermas y sus relaciones térmicas con las altitudes podrían servir para señalar la presencia o no de estas vegas y o sus composiciones o viceversa señalar el papel bioindicador de las comunidades de vegas para inferir las condiciones térmicas de los sitios de las vegas.
Existe un grupo de plantas como Rorippa nasturtium-aquaticum, Veronica anagallis-aquatica, Juncus septati, Ranunculus sp., Poa annua,, Cotula coronopifolia, Polypogon monspeliensis, Carex gayana, Eleocharis albibracteata, Anemone multifida , que tienen una amplia distribución tanto latitudinal como altitudinal siendo comunes en las dos alturas. Son especies que toleran amplios rangos térmicos.
Por todo esto nos lleva a inferir los límites de las vegas del área a los siguientes pisos altitudinales con un ecotono entre ellos de 100- 200 m.
1. Piso de las vegas de baja altura entre 1200 a 2000 m s.n.m.
2. Piso de las vegas de mediana altura entre 2000 a 3200 m s.n.m.
3. Piso de las vegas de la tundra andina por encima de los 3200 m s.n.m.
Sobre este último debemos seguir trabajando a fin de obtener mayor información y ajustes de los límites en el futuro.
Tabla 12. Datos de temperaturas de las localidades del área analizada (Cordón del Plata, Luján de Cuyo, Mendoza, Argentina). Localidades (a-j): ubicación en el mapa de la figura 1.

Tabla 13. Datos de temperaturas de los límites inferiores y superiores de las vegas del Cordón del Plata (Mendoza, Argentina).

CONCLUSIONES
El estudio de la vegetación de las vegas del flanco oriental del Cordón del Plata releva:
1. La existencia de dos tipos de vegas: de baja altura y de altura, definidas florística, fitosociológica y ecológicamente.
2. En el área estudiada existen 20 comunidades de vegas: 9 en las vegas de baja altura y 11 en las de altura, las que comprenden desde comunidades acuáticas a de terrenos casi secos.
3. Mientras las vegas de altura son dominadas por elementos autóctonos, en la de baja altura dominan los especies alóctonas.
4. No se encontró un espectro biológico con todas las formas de vida.
5. Existen 104 especies en total: 48 exclusivas en las vegas de baja altura y 40 en las de altura, mas 16 especies comunes.
6. Las comunidades, en cada nivel altitudinal, se disponen en un gradiente de humedad desde lo más húmedo a lo más secos. No todos los perfiles estudiados contienen o están completos con las comunidades.
7. Las vegas del área se hallan distribuídas en un gradiente altitudinal dentro de 3 provincias fitogeográficas: del Monte, andina y altoandina. La vegas de baja altura están dentro de los pisos vegetales de Larrea cuneifolia y L. divaricata , para el Monte, y las vegas de altura en los de Junellia scoparia y Colliguaja integerrima , Adesmia horrida, A. pinifolia para la andina y probablemente las vegas de tundra en los de Adesmia subterranea y Azorella monantha en la Altoandina , respectivamente.
AGRADECIMIENTOS
A la Dra. Bárbara Ruthsatz por la bibliografía aportada, la lectura y valiosas sugerencias sobre el trabajo; a los revisores anónimos por sus correcciones; a Judy Boshoven por su ayuda en el resumen en inglés y a Cecilia Scoones por mejorar los dibujos originales.
BIBLIOGRAFÍA
BÖCHER, T. W., J. P. HJERTING & K. RHAN. 1963. Botanical Studies in The Atuel Valley Area, Mendoza, Province Argentina. Dansk Botanisk Arkiv . 22:1 (1-125) Kopenhagen. [ Links ]
BÖCHER, T. W., J. P. HJERTING & K. RHAN. 1968. Botanical Studies in The Atuel Valley Area, Mendoza, Province Argentina. Dansk Botanisk Arkiv . 22: 2 (121- 185). Kopenhagen. [ Links ]
BÖCHER, T. W., J. P. HJERTING & K. RHAN. 1972. Botanical Studies in The Atuel Valley Area, Mendoza, Province Argentina. Dansk Botanisk Arkiv . 22: 3 (191-358). Kopenhagen. [ Links ]
BRAUN -BLANQUET, J. 1979. Fitosociología. Bases para el estudio de las comunidades vegetales . Ed. H. Blume, Madrid 820 pp. [ Links ]
BUK, E. M. 1983. Glaciares de escombros y su significación hidrológica. Actas de la Primera Reunión Grupo Periglacial Argentino, Anales del IANIGLA 5: 22-38. [ Links ]
CABIDO, M. 1985. 1985. Las comunidades vegetales de la Pampa de Achala, Sierras de Córdoba, Argentina. Documents Phytosociologiques . 9:431-456. [ Links ]
CABRERA , A. L. 1976. Regiones Fitogeográficas de la República Argentina. Enciclopedia Arg. Agricult. y Jardinería 2ed. , 2(1): 1-85. ACME, Buenos Aires. [ Links ]
CAMINOS, R. 1965. Geología de la vertiente oriental del Cordón del Plata, Cordillera Frontal de Mendoza. Revista Asoc. Geológ. Argent. 20 (3): 351-392. Bs As. [ Links ]
CAMINOS, R. 1979. Cordillera Frontal. Geología Regional Argentina. Acad. Nac. Cs , Cordoba. [ Links ]
CLEEF, A. M. 1981. The vegetation of the Páramos of the Columbian Cordillera Oriental. Diss. Bot. 61: 1-320. [ Links ]
CORTE, A. 1983. Los conceptos geocriogénico-parageocriogénico y glacial-periglacial en los Andes Centrales de Argentina, latitud 30º. Actas de la Primera Reunión Grupo Periglacial Argentino , Anales del IANIGLA 5: 48-61. [ Links ]
CORTE, A. 1983. Geocriología. El Frío en la Tierra. Mendoza , Ediciones culturales Mendoza, Argentina, 397 pp. [ Links ]
DE FINA , A. L., GIANNETTO, F., RICHARD, A. E. & L. S. SABELLA 1964.Difusión geográfica de los cultivos índices de la provincia de Mendoza y sus causas. INTA. Inst. de Suelos y Agrotec. 83p. [ Links ]
GALÁN DE MERA, A, C. CÁCERES Y A GONZÁLEZ. 2003. La vegetación de la alta montaña andina del sur del Perú. Acta Botánica Malacitana 28: 121-147. [ Links ]
GANDULLO, R. & A. M. FAGGI 2003. Análisis fitosociológico de los mallines andinos del Parque Provincial Copahue. Neuquén Argentina. Kurtziana 30: 45-55. [ Links ]
GANDULLO, R. & A.M. FAGGI. 2006. Interpretación sintaxonómica de los humedales del noroeste de la provincia de Neuquén, Argentina. Darviniana 43 (1-4): 10-29. [ Links ]
GONZALEZ LOYARTE, M. M. 1989. Los pastizales de Piedemonte: Los coironales preandinos de Tupungato, Mendoza, Argentina . Detección y Control de la desertificación, Conferencias, Trabajos y resultados del Curso Latinoamericano , 242-253 [ Links ]
GONZALEZ LOYARTE, M. M. & I. E. PERALTA 2004. Flora y Vegetación de la cuenca del río Pachón (Calingasta, San Juan, Argentina). Bol. Soc. Argent. Bot. 39 (3-4): 283-300. [ Links ]
GUTTE, P. 1980. Beitrag zur Kenntnis zentralperuanuischer Pflanzengesellschften II. Die hochandinen Moore und Ihre Kontaktgesellschaften. Feddes Rep. 91 : 327-336. Berlín. [ Links ]
HAUMAN, L. 1918. La végétation des Hautes Cordillères de Mendoza. Anal. Soc. Cient. Argent. 86: 121-188, 255-348. Buenos Aires. [ Links ]
MARTINEZ CARRETERO, E. 1997. The Puna vegetation in the Valley of Rio Cazaderos, Catamarca Province, Argentina. Candollea 52: 497-508. [ Links ]
MÉNDEZ, E. 1986. Dinamismo de la vegetación en el Valle Superior del río Atuel. I. Vegas y conos de deyección . Parodiana 4: 33-350. [ Links ]
MÉNDEZ, E. 1990 a . Dinamismo de la vegetación de la Pampa Amarilla , San Rafael, Mendoza. Multequina 1: 73-81. [ Links ]
MÉNDEZ, E. 1990 b. La vegetación de los embanques aluvionales de las nacientes del Embalse El Carrizal, Provincia de Mendoza y sus dinamismos I. Los bosques espontáneos de Salix. Parodiana 6: 271-288. [ Links ]
MÉNDEZ, E. 2004. La vegetación de los Altos Andes. I Pisos de vegetación del flanco oriental del Cordón del Plata, Mendoza Argentina Bol. Soc. Argent. Bot. 39 (3-4): 227- 253. [ Links ]
MÉNDEZ, E. 2005. La vegetación del Parque Provincial Laguna de Llancanello. Candollea 60 (1): 67-74 [ Links ]
MÉNDEZ, E., E. MARTÍNEZ CARRETERO & I. PERALTA 2006. La vegetación del Parque Provincial Aconcagua. (Altos Andes Centrales de Mendoza, Argentina). Bol. Soc. Argent.. Bot. 41(1-2): 41-69. [ Links ]
MIKKAN, R. A. 1988. Geomorfología de la zona de influencia de la Presa Potrerillos . Bol. Est. Geog. UNC . 25: :91-114. [ Links ]
MINETTI, J. H. & A. E. CORTE. 1984. Zonificación latialtitudinal del clima en la zona andina y su relación con el límite inferior del hielo perenne (LIHP) y del límite inferior geocriogénico (LIG): Anales del IANIGLA : 129-144. [ Links ]
RAUNKIAER, C. 1934 . Life forms and terrestrial plant geography. Clarendon Press, Oxford [ Links ]
RIVAS MARTÍNEZ, S & O. TOVAR. 1982. Vegetatio andinae, I. Datos sobre comunidades vegetales altoandinas de los Andes Centrales del Perú. Lazaroa 4: 167-187. [ Links ]
ROIG, F. 1976. Las comunidades vegetales del Piedemonte de la Precordillera de Mendoza . Ecosur 3: 1- 45. [ Links ]
ROIG, F. A. 1998. La vegetación de la Patagonia . En M. E. Correa (Ed.) Flora Patagonica. Colección Científica del INTA. Tomo VII. (I) 48-170. Buenos Aires. [ Links ]
ROIG, F. A & V. MARONE. 1982. Radiación solar y la distribución de la vegetación en la precordillera de Mendoza. Serie Científica 5(27): 9- 13. [ Links ]
ROIG, F. A., ANCHORENA , J., DOLLENZ, O., FAGGI, A. M. & MÉNDEZ, E. 1985. Las comunidades vegetales de la Transecta Botánica de la Patagonia Austral. Primera parte: Area continental, pp. 350-456 en Boelcke, O., Moore, D. M. , Roig, F. A. (eds.), Transecta Botánica de la Patagonia Austral. Conicet, Royal Society, Instituto de la Patagonia. Buenos Aires [ Links ]
ROIG, F, , ABRAHAN, M. E. & E. MÉNDEZ. 2007. Vegetation belts, cold and soil freezing in the Central Andes of Mendoza, Argentina. Phytocoenologia 37 (1): 99-114. Berlin, Stuttgart. [ Links ]
RUTHSATZ, B. 1977. Pflanzengesellschaften und ibre Lebensdingungen in dem Adinen Halbwüsten Nordwest-Argentiniens Diss. Bot. 39: 1-168. [ Links ]
RUTHSATZ, B. 1995. Vegetation und Ökologie tropischer Hochgebirsgsmoore in den Anden Nord-Chiles. Phytocoenologia 25(2): 185-234. Berlín -Stuttgart. [ Links ]
RUTHSATZ, B. 2000. Die hartpolstermoore der Hochanden und ihre Artenvielfalt. Ber. D. Reinh.-Tüxen-Ges , 12: 351-371. Hannover. [ Links ]
VIERS, G. 1964. La depresión de Potrerillos. Estudio geomorfológico. Bol. Est. Geog. UNC . 43: 73-101. [ Links ]
ZULOAGA, F. O., NICORA, E. G., RÚGOLO DE AGRASAR, Z., MORRONE, O., PENSIERO, J., CIALDELLA, A. M. 1994. Catálogo de la Familia Poaceae en la República Argentina. Monogr. Syst. Bot. Missouri Bot. Gard . 47: 1-178. [ Links ]
ZULOAGA, F. O. & O. MORRONE. 1996. Catálogo de las plantas vasculares de la república Argentina. I. Pteridophyta, Gymnospermae y Angiospermae (Monocotyledoneae). Monogr. Syst. Bot. Missouri Bot. Gard. 60 : 1-323. [ Links ]
ZULOAGA, F. O. & O. MORRONE. 1999a. Catálogo de las plantas vasculares de la república Argentina. II. Acanthaceae-Euphorbiaceae (Dicotyledoneae). Monogr. Syst. Bot. Missouri Bot. Gard . 74: 1-621. [ Links ]
ZULOAGA, F. O. & O. MORRONE. 1999a. Catálogo de las plantas vasculares de la república Argentina. II. Fabaceae-Zygophyllaceae (Dicotyledoneae). Monogr. Syst. Bot. Missouri Bot. Gard. 74: 1-1269. [ Links ]
Recibido el 09 de Octubre de 2006,
aceptado el 10 de Octubre de 2007.

