










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.43 n.1-2 Córdoba ene./jul. 2008
Desarrollo foliar y caulinar de las unidades de alargamiento de Nothofagus dombeyi (Nothofagaceae) en condiciones de alta y baja luminosidad
Cecilia Calabria1 y Javier Puntieri1, 2
1Departamento de Botánica, Centro Regional Universitario Bariloche, Universidad Nacional del Comahue, Quintral 1250, 8400 Bariloche. cgcalabria@netlynx.com.ve
2Carrera del Investigador Científico y Tecnológico, CONICET. jpuntier@crub.uncoma.edu.ar
Resumen: Se analizaron el tamaño del tallo y las hojas que componen las unidades de alargamiento (UA) del tronco, las ramas principales y las ramas cortas de ejemplares juveniles de Nothofagus dombeyi desarrollados en comunidades de áreas abiertas (bordes de camino) y de sotobosque de Patagonia. Se evaluaron el área y la forma (ancho/longitud) de las hojas a lo largo de las UA, el área foliar específica (AFE) y las relaciones área foliar/volumen del tallo, longitud del tallo/peso del tallo y volumen del tallo/peso del tallo. Para igual posición en la UA, las hojas del tronco son mayores que las de las ramas principales y éstas mayores que las de las ramas cortas. En las UA del tronco el desarrollo caulinar es proporcionalmente mayor al de las hojas en comparación con las UA de las ramas principales y, en especial, de las ramas cortas. La restricción en el desarrollo caulinar y el aumento del AFE en ejemplares de N. dombeyi creciendo a la sombra permitiría su supervivencia pero con limitado desarrollo en altura y, en consecuencia, baja probabilidad de alcanzar el dosel.
Palabras clave: Área foliar; Categoría de ejes; Coihue; Nothofagaceae; Nothofagus; Unidad de alargamiento.
Summary: Leaf and stem development in extension units of Nothofagus dombeyi (Nothofagaceae) under high and low light conditions. The size of stem and leaves was studied for trunk, main branch and short branch extension units (EU) of young Nothofagus dombeyi trees growing in open areas (road verges) and understory communities in Patagonia. The surface area and outline (width/length ratio) of single leaves along EU, the specific leaf area, and the ratios leaf area/stem volume, stem length/stem mass and stem volume/stem mass were assessed. Trunk leaves were larger than main-branch leaves and main-branch leaves larger than short-branch leaves in similar positions on the EU. Stem developed proportionally more than leaves in trunk EU than in main-branch EU and, especially, short-branch EUs. Restrictions in stem development and increases in specific leaf area would allow N. dombeyi trees to survive under shaded conditions though limiting their height growth and, consequently, their chances to reach the canopy.
Key words: Axis categories; Coihue; Extension unit; Leaf area; Nothofagaceae; Nothofagus.
Introducción
Las hojas son unidades funcionales de la fotosíntesis y la transpiración de las plantas vasculares. El tamaño foliar y su relación con el tamaño del tallo son relevantes para el crecimiento de una planta. Desde hace varias décadas se ha buscado explicar las variaciones inter- e intraespecíficas en el tamaño y la forma de las hojas y sus implicancias funcionales (Schaffalitzky de Muckadell, 1959; Critchfield, 1960; Parkhurst & Loucks, 1972; Givnish& Vermeij, 1976; Björkman, 1981; Givnish, 1982, 1988; Kozlowski & Pallardy, 1997; Barthélémy & Caraglio, 2007), pero pocos estudios han considerado a las hojas en el contexto del desarrollo caulinar del cual dependen (e.g. Nicolini & Chanson, 1999; Puntieri et al., 2001). Dados el grado de amificación que puede alcanzar el sistema caulinar en las especies arbóreas y la diferenciación morfológica entre los ejes componentes (Puntieri et al., 2003), podrían predecirse variaciones intraespecíficas en la estructura foliar paralelas con esa diferenciación. La aplicación de los conceptos de gradiente ontogenético y de categoría de ejes, estimulados a partir de los estudios de síntesis sobre la arquitectura de las plantas (Hallé et al., 1978; Tourn et al., 1999; Barthélémy & Caraglio, 2007), permite una aproximación sistemática y repetible a esta temática.
En numerosas especies leñosas, cada eje constituyente del sistema caulinar suele alargarse en forma rítmica, de modo que alternan períodos de alargamiento de los ejes y períodos en los cuales la longitud de los mismos se mantiene constante, por lo menos a nivel macroscópico. En general, esto permite identificar las porciones de un eje correspondientes a sucesivos períodos de alargamiento, las que en adelante se denominarán «unidades de alargamiento» (UA). La UA puede ser considerada una unidad estructural y funcional de un eje, dada la relativa simultaneidad con que se desarrollan el tallo y las hojas que la componen (Barthélémy & Caraglio, 2007). El análisis del área foliar total de una UA y su relación con el tamaño del tallo del cual deriva puede permitir una mejor comprensión del funcionamiento de las UA de cada categoría de ejes de un árbol.
El género Nothofagus Blume cuenta con 35 especies, nueve de las cuales habitan en los bosques subantárticos de Sudamérica. Este género se considera relevante no solo en la dinámica de los bosques sino también en la comprensión de las relaciones evolutivas entre las especies vegetales actuales (Veblen et al., 1996). Nothofagus dombeyi (Mirb.) Oersted es una especie leñosa perennifolia que habita a ambos lados de la cordillera de los Andes entre los 34° 30' S y 47° S en Chile y Argentina (Donoso et al., 2006). Forma bosques mixtos o puros, cuya dinámica se vincula estrechamente con la ocurrencia de disturbios (Veblen, 1989). Es una especie pionera considerada intolerante a la sombra (Donoso, 1992; Donoso et al., 2006). Sin embargo, por años pueden permanecer individuos con crecimiento limitado bajo el dosel de árboles adultos de esta especie (Stecconi, 2006).
Estudios recientes han analizado las características arquitecturales de N. dombeyi y clasificado los ejes de individuos juveniles en tres categorías principales, las cuales difieren en la morfología y, presumiblemente, en la fisiología: (a) el eje principal o tronco, generalmente único, formado por brotes largos de desarrollo vertical, (b) las ramas principales, que derivan del tronco y crecen en forma horizontal u oblicua, y (c) las ramas cortas, que crecen horizontalmente a partir del tronco y de las ramas principales (Puntieri et al., 2003). Mientras que el tronco y las ramas principales son ejes de larga vida, las ramas cortas permanecen por pocos años (Stecconi, 2006). Hasta el momento se desconoce, para esta especie, la medida en la cual la morfología foliar se vincula con factores endógenos (como la categoría de ejes) y factores exógenos (como la luminosidad del ambiente donde crece el árbol).
El presente estudio evalúa las siguientes hipótesis: (1) la morfología de las hojas de cada UA depende de la categoría de ejes a la que ésta pertenece; (2) la posición sobre una UA del nudo de origen de una hoja determina en buena medida la forma y tamaño de la misma; (3) el área y la biomasa de las hojas de una UA y sus relaciones con el tamaño del tallo varían de acuerdo a la categoría de ejes a la que pertenece la UA y a la disponibilidad de luz en el ambiente.
Materiales y Método
Sitios y poblaciones de muestreo
Se realizaron muestreos en tres comunidades de bosque mixto y de bosque puro de N. dombeyi ubicadas en el Parque Nacional Nahuel Huapi. Las mismas fueron escogidas por su accesibilidad y alto número de ejemplares coetáneos de N. dombeyi. La precipitación anual media en esta zona es de 8001000 mm, y la temperatura media anual es de 8-9° C (Conti, 1998). El suelo en la región deriva de cenizas y sedimentos volcánicos y piroclásticos y es rico en materia orgánica superficial (Scoppa, 1998). Las tres comunidades pueden describirse como bosque maduro dominado por coihue, con abundante regeneración en claros producidos por disturbios naturales o antrópicos. En cada uno de los sitios de muestreo es posible encontrar dos tipos de condiciones lumínicas extremas para los renovales: un ambiente relativamente sombrío, bajo el dosel de los árboles dominantes (cobertura de entre 67% y 87% estimada fotográficamente), y un ambiente con incidencia solar notablemente mayor (0% de cobertura), donde el dosel arbóreo ha sido drásticamente interrumpido por la construcción de un camino. Estos dos ambientes serán referidos en este trabajo como ambiente sombreado o sotobosque y ambiente abierto, respectivamente. Puede considerarse que la incidencia de luz fue el factor ambiental que más difirió entre estos ambientes en cada sitio de muestreo.
Población Cerro López: se ubica en un área de alrededor de 0,5 ha en la base de la ladera norte del Cerro López, a una altitud aproximada de 800 msnm (41° 05' 33'' S, 71° 32' 50'' O) y a una distancia de 30 km hacia el oeste del centro de la ciudad de San Carlos de Bariloche. Se trata de un bosque mixto cuyo estrato arbóreo está dominado por individuos de N. dombeyi de entre 10 y 41 m de altura y 1,06 ± 0,067 m de diámetro a la altura del pecho (DAP; media ± error estándar, EE; N = 6), y Austrocedrus chilensis (D. Don) Pic. Serm. & Bizzarri de entre 9 y 15 m de altura y 0,27 ± 0,035 m (N = 3) de DAP. Individuos de N. dombeyi de 15 a 25 años de edad dominan en el sotobosque, donde también hay ejemplares dispersos de Lomatia hirsuta (Lam.) J.F. Macbr., Maytenus chubutensis (Speg.) Lourtig, O'Donell & Sleumer, Berberis darwinii Hook. y Schinus patagonicus (Phil.) Cabrera.
Población Mascardi: ocupa alrededor de 1 ha sobre la margen oeste del lago Mascardi, a una altitud de 830 msnm (41° 21' 43'' S, 71° 33' 54'' O) y a 59 km al sur del centro de San Carlos de Bariloche. La población de muestreo se encuentra en un bosque puro de N. dombeyi con ejemplares de entre 26 y 44 m de altura y 0,21 ± 0,029 m (N = 6) de DAP. El sotobosque está dominado por individuos jóvenes de coihue de entre 12 y 21 años de edad. En los estratos arbóreo bajo y arbustivo también se encuentran ejemplares aislados de Lomatia hirsuta, Embothrium coccineum J.R. Forst. & G. Forst., Nothofagus antarctica (G. Forst) Oerst., Aristotelia chilensis (Molina) Stuntz y Schinus patagonicus, entre otras especies.
Población Angostura: ocupa alrededor de 1,5 ha a 200 m al este del arroyo Pedregoso, sobre la margen noreste del lago Nahuel Huapi, a 770 msnm (40° 59' 10'' S, 71° 27' 34'' O). Se ubica a 12 km al este de la localidad de Villa La Angostura, bordeando la ruta 231. Se trata de un bosque mixto de N. dombeyi y Austrocedrus chilensis. El dosel está conformado por individuos de coihue de entre 34 y 36 m de altura y DAP de 0,41 ± 0,063 m (N = 3). En el sotobosque predominan ejemplares dispersos de N. dombeyi, Aristotelia chilensis, Schinus patagonicus, Lomatia hirsuta, Maytenus chubutensis, Berberis darwinii y Rosa rubuginosa L., entre otras especies.
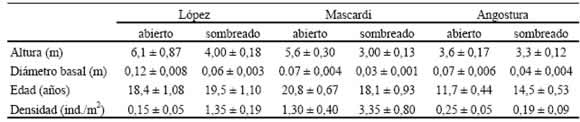
Para cada uno de los sitios de muestreo y cada uno de los dos ambientes, soleado y abierto, se seleccionaron diez ejemplares de una cohorte de individuos jóvenes o renovales de N. dombeyi. Cada cohorte presentaba poca variación de edad, la que fue estimada en forma no destructiva a partir del reconocimiento de límites entre UA sucesivas (Puntieri et al., 1998, 2000; Stecconi, 2006) y tamaño entre individuos (Tabla 1). Se excluyeron aquellos ejemplares con daño evidente sobre el tronco y las ramas principales. Se midieron la altura y el diámetro basal de cada individuo seleccionado y la densidad de individuos de cada cohorte, muestreada en parcelas de entre 4 y 25 m2, según la densidad de la población (Tabla 1).
Tabla 1. Promedio (± error estándar) de altura, diámetro basal, edad de los individuos y densidad poblacional para renovales de Nothofagus dombeyi en tres sitios y dos ambientes lumínicos (abierto y sombreado). Para las primeras tres filas los promedios corresponden a los 10 individuos de los que se cosecharon brotes.
Muestreos
Se realizaron dos muestreos independientes entre marzo y abril de 2001, luego de finalizado el alargamiento de las UA del período 2000-2001. Para ambos muestreos se tomó en cuenta la siguiente categorización de ejes: (a) tronco, (b) rama principal y (c) rama corta (Barthélémy et al., 1999; Stecconi, 2006). De aquí en adelante, estas categorías serán abreviadas como TR, RP y RC, respectivamente.
Muestra 1. De cada árbol seleccionado se cortaron las UA desarrolladas en las temporadas de crecimiento 1999-2000 y 2000-2001 para un eje de cada una de las categorías de ejes indicadas. Las UA seleccionadas se encontraban por encima de 1,5 m sobre el nivel del suelo. Los límites entre UA se identificaron por la presencia de catafilos o cicatrices de los mismos y la longitud de los entrenudos (mínima en ambos extremos de una unidad de crecimiento; Barthélémy et al., 1999). Para cada UA cosechada se registraron las siguientes características: (1) longitud del tallo (con cinta métrica y 0,1 cm de precisión, desde el límite proximal de la UA hasta el extremo proximal de la yema terminal o el límite con la UA siguiente), (2) diámetro del tallo en su extremo proximal, inmediatamente por encima del límite entre UA sucesivas, (3) diámetro distal del tallo, por debajo del límite mencionado (ambos diámetros medidos con calibre con una precisión de 0,1 mm) y (4) número de nudos. A partir de los diámetros proximal y distal del tallo de una UA se calculó el diámetro promedio del tallo. Las hojas de cada UA fueron numeradas desde el extremo proximal hasta el extremo distal de la UA, correspondiendo la posición uno al catafilo ubicado en el extremo proximal de la UA (hay uno o dos catafilos en la base de cada UA en N. dombeyi; Puntieri et al., 1998). Las hojas de cada UA fueron escaneadas con una definición de 300 dpi y el área, longitud y ancho máximo de cada lámina fueron obtenidos digitalmente (programa Scion Image). A partir del cociente entre el ancho máximo y la longitud de cada lámina se obtuvo una evaluación de su forma. Las imágenes de las hojas con pequeñas porciones faltantes por daño fueron tratadas digitalmente a fin de completar esas porciones. En los casos en que la porción faltante hizo imposible la estimación del tamaño completo de la lámina, la misma fue considerada faltante.
Muestra 2. Se seleccionaron 90 láminas foliares no dañadas de cada una de las dos condiciones lumínicas de cada sitio: 30 de cada categoría de ejes y 15 de cada uno de los períodos de alargamiento 19992000 y 2000-2001. Se escanearon las láminas y se midió el área foliar de cada una. Se cosecharon 60 UA por sitio (10 por categoría de eje y para cada uno de los períodos de alargamiento 1999-2000 y 2000-2001) y se registraron la longitud y los diámetros proximal y distal del tallo. Láminas y tallos fueron secados en estufa a 70° C hasta peso constante (48 horas), y pesados con una precisión de 0,001 g.
Análisis estadístico de los datos
Previamente a los análisis estadísticos, se analizó la homoscedasticidad de las variancias para cada variable con el test F max de Hartley (Sokal & Rohlf, 1981). Las varianzas heterogéneas se homogeneizaron mediante transformación logarítmica para su análisis estadístico. Para todas las comparaciones se adoptó un nivel de significación del 5 %.
Muestra 1. Se evaluaron mediante análisis de varianza (ANDEVA; Sokal & Rohlf, 1981) los efectos de: categoría de ejes (factor fijo), ambiente lumínico (sombreado o abierto; factor fijo), sitio de muestreo (factor aleatorio) y año de alargamiento (factor aleatorio) sobre longitud, número de nudos, área de la lámina de área mayor por UA y posición de la hoja de área mayor relativa al total de nudos de la UA, calculada como: [número del nudo de la hoja mayor contado desde el extremo proximal de la UA] / [número de nudos de la UA].
Se analizaron las variaciones en área foliar y forma de la lámina en función de la posición de la hoja en la UA. Para ello se promediaron las áreas o los cocientes ancho/longitud correspondientes a cada posición, para las UA de igual categoría de ejes, ambiente lumínico y sitio. Este análisis se realizó solamente para las UA desarrolladas en el período 2000-2001 debido al elevado número de hojas faltantes en las UA del período 1999-2000.
Se analizó la relación entre la sumatoria de las áreas foliares de una UA completa y el volumen de su tallo. Este último fue estimado a partir de su longitud y diámetro medio, suponiendo que se trata de un cilindro. El área de cada lámina faltante se estimó promediando las áreas de las hojas en posiciones inmediatamente proximal y distal con respecto a la hoja faltante. En los casos en que dos hojas sucesivas hubieran estado ausentes o muy dañadas para ser reconstruidas, se promediaron las áreas de las hojas inmediatamente proximal y distal con respecto a las dos hojas faltantes o dañadas y se asignó a estas dos últimas el mismo valor de área foliar. Las UA con más de dos hojas sucesivas faltantes se excluyeron del análisis. La relación entre [suma de las áreas foliares] / [volumen del tallo] de una UA fue comparada entre categorías de eje, ambientes lumínicos (factores fijos), sitios de muestreo y año de alargamiento (factores aleatorios) mediante ANDEVA.
Las UA cortadas de todos los sitios fueron ordenadas mediante un análisis de componentes principales (ACP; Sokal & Rohlf, 1981) incorporando las variables: (1) número de nudos, (2) volumen del tallo, (3) área foliar máxima, (4) suma de las áreas foliares, (5) relación [suma de las áreas foliares] / [volumen del tallo], (6) posición relativa de la hoja de área máxima y (7) forma de la hoja de área máxima.
Muestra 2. Se calcularon las relaciones [longitud del tallo (cm)] / [peso seco del tallo (g)] y [volumen (cm3) / peso seco del tallo (g)], y área foliar específica (AFE = [área foliar (cm2)] / [peso seco de la lámina (g)]). Estas relaciones fueron comparadas entre categorías de ejes, ambientes lumínicos (factores fijos), sitios de muestreo y años de producción de las UA (factores aleatorios) mediante ANDEVA.
Resultados
Muestra 1. Tanto la longitud como el número de nudos de las UA variaron significativamente con la categoría de ejes, el ambiente lumínico, el sitio de muestreo y el período de alargamiento (Tabla 2). Los promedios para ambas variables fueron máximos para el TR, intermedios para las RP y mínimos para las RC. Dentro de cada categoría de ejes, las UA de ambiente abierto presentaron promedios de longitud y número de nudos mayores que las de ambiente sombreado (Fig. 1 A y B). El área de la lámina mayor por UA varió significativamente entre categorías de ejes (TR > RP > RC), pero no entre ambientes lumínicos (Tabla 2; Fig. 1 C); los efectos de sitio y de período de alargamiento de la UA fueron significativos (Tabla 2). La hoja de lámina mayor por UA se desarrolló en posición relativamente más distal en las RC que en las RP y más proximal en el TR que en las RP y RC, y en posición relativamente más distal en ambiente sombreado que en ambiente abierto (Fig. 1 D). Tanto el sitio como el período de alargamiento afectaron significativamente la posición de la hoja de mayor área foliar (Tabla 2).
Tabla 2. Estadístico F de Fisher y grado de significación del mismo (p) para los factores fijos categoría de eje (Cat. eje) y disponibilidad de luz (Luz), y los factores aleatorios período de alargamiento (Período) y sitio de muestreo (Sitio), sobre lassiguientes variables descriptoras de las UA: longitud del tallo (Long.), número de nudos (Nudos), área foliar mayor (Áream), posición de la hoja de área mayor relativa al número de nudos de la UA (Pos.), relación [sumatoria del área foliar] / [volumen del tallo] (Σ Área/vol.), área foliar específica (AFE), relación [longitud] / [peso seco del tallo] (Long./peso) y relación [volumen] / [peso seco del tallo] (Vol./peso). g.l.: grados de libertad totales. *** p < 0,001, ** p < 0,01, * p < 0,05, ns p > 0,05.


Fig. 1. A: Longitud. B: Número de nudos. C: Área de la hoja mayor. D: Posición de la hoja mayor en relación al número de nudos de la UA (posición relativa; media ± límite de confianza del 95 %). A-D para tres categorías de ejes (TR: tronco, RP: rama principal, RC: rama corta) de plantas de Nothofagus dombeyi desarrolladas en ambiente abierto (símbolos blancos) y en ambiente sombreado (símbolos negros).
Para ambas condiciones lumínicas y todas las categorías de ejes, el área de las hojas se incrementó en las primeras cuatro posiciones más proximales de las UA (Fig. 2). Hacia posiciones más distales, el área foliar disminuyó notablemente en todos los casos, aunque en las UA de TR y RP de ambiente abierto, tendió a aumentar nuevamente entre los nudos 20 y 25, no representados en los otros grupos de UA, aunque con elevada variación de área foliar entre UA (Fig. 2 A). La relación ancho/longitud de la lámina tendió a disminuir desde el extremo proximal al extremo distal de las UA excepto en el caso de las UA de TR y RP de ambiente abierto, en las cuales se incrementó, aunque con elevada variación, en posiciones intermedias (Fig. 3). Para cada posición, la relación ancho/longitud de las láminas fue superior en TR que en RP y RC y mayor en RP que en RC (Fig. 3).

Fig. 2. Área de las hojas según su posición en la unidad de alargamiento (UA) para el tronco, las ramas principales y las ramas cortas de árboles de Nothofagus dombeyi, desarrollados en ambiente abierto (A) y en ambiente sombreado (B).

Fig. 3. Relación ancho/longitud de las hojas según su posición a lo largo de la unidad de alargamiento (UA) para el tronco, las ramas principales y las ramas cortas de árboles de Nothofagus dombeyi, desarrollados en ambiente abierto (A) y en ambiente sombreado (B).
La relación [sumatoria de área foliar] / [volumen del tallo] fue afectada significativamente por la categoría de ejes, el ambiente lumínico, el sitio de muestreo y el período de desarrollo de la UA (Tabla 2). Esta relación fue mayor en RC que en RP y mayor en éstas que en TR (Fig. 4 A). Para cada categoría de ejes esta relación fue mayor en el ambiente sombreado que en el ambiente abierto.

Fig. 4. A: Relación [área foliar] / [volumen del tallo]. B: Área foliar específica (AFE). C: Relación [longitud del tallo] / [peso del tallo]. D: Relación [volumen del tallo] / [peso del tallo] (media ± límite de confianza del 95%). A-D para tres categorías de ejes (TR: tronco, RP: rama principal, RC: rama corta) de plantas de Nothofagus dombeyi, desarrolladas en ambiente abierto (símbolos blancos) y en ambiente sombreado (símbolos negros).
El primer eje de ordenamiento de las UA obtenido mediante ACP explicó un 57 % de la variación total, separando a las UA del TR de las de las RC y ubicando las UA de las RP en posiciones intermedias (Fig. 5). Las variables de mayor influencia sobre este componente fueron aquellas relativas al tamaño de las UA (valores altos de número de nudos, volumen del tallo y área foliar para UA del TR), la relación [sumatoria de área foliar] / [volumen del tallo] y la posición relativa de la hoja de mayor área (valores altos para UA de RC). El segundo componente explicó el 14 % de la variación y la variable de mayor influencia sobre el mismo fue el coeficiente de forma (ancho/ longitud) de la hoja mayor (Fig. 5).

Fig. 5. Ordenamiento de las UA de Nothofagus dombeyi a lo largo de los dos primeros ejes obtenidos mediante análisis de componentes principales. Se indican con diferentes símbolos las UA del tronco (círculos negros), de las ramas principales (cuadrados blancos) y de las ramas cortas (cruces). Las flechas indican los efectos proporcionales de las variables incorporadas en el análisis sobre los ejes de ordenamiento graficados.
Muestra 2. Tanto el AFE como la relación [longitud del tallo] / [peso del tallo] fueron significativamente afectados por todos los factores incluidos en el análisis (Tabla 2). Ambas variables fueron notablemente mayores en ambiente sombreado que en ambiente abierto y aumentaron siguiendo el gradiente TR < RP < RC (Fig. 4 B y C). La relación [volumen del tallo] / [peso del tallo] fue significativamente afectada por el período de alargamiento de la UA y por la categoría de ejes (máxima para TR), pero no por el sitio de muestreo ni por el ambiente lumínico (Fig. 4 D; Tabla 2).
Discusión
Variaciones en el tamaño y la forma de las hojas en N. dombeyi
La categoría de ejes a la cual pertenece una UA tiene un efecto significativo sobre el área de las hojas de N. dombeyi, tanto en individuos desarrollados en sotobosque como en aquellos que se encuentran en ambiente abierto: UA del tronco desarrollan, en promedio, hojas de mayor área que UA de las ramas principales, y éstas, a su vez, hojas mayores que UA de las ramas cortas. Para las UA de N. dombeyi de tamaño corto a intermedio (con hasta 17 nudos), el área de las hojas aumenta notablemente entre las hojas más proximales y disminuye gradualmente hacia el extremo distal de la UA. En UA más largas se observa un segundo incremento de tamaño foliar (menos definido que el primero) hacia posiciones más distales, seguido de una disminución gradual.
Las diferencias en el área de las hojas entre categorías de ejes y las variaciones en el área de las hojas a lo largo de UA de tamaño corto a intermedio son semejantes a las encontradas para N. pumilio (Puntieri et al., 2001), N. antarctica, ejemplares híbridos entre N. dombeyi y N. antarctica (Stecconi et al., 2004) y N. alpina (Puntieri et al., 2007). Al considerar la posición de las hojas a lo largo de cada UA de N. dombeyi, puede advertirse que, para una posición determinada dentro de una UA, el tronco presenta, en promedio, hojas de mayor tamaño y más ovadas que las ramas principales, y éstas, a su vez, hojas mayores y más ovadas que las ramas cortas. Diferencias en el tamaño y la forma de las hojas en función de la categoría de ejes y de la posición de las hojas sobre las UA fueron encontradas también en Fagus sylvatica (Nicolini & Chanson, 1999), así como en especies de la familia Cupressaceae (Grosfeld, 2002).
Las variaciones tanto en el tamaño como en la forma de las hojas a lo largo de una UA determinada podrían explicarse mediante un argumento adaptativo: la distribución del área foliar en cada UA tendería a maximizar la intercepción de radiación y minimizar el solapamiento entre hojas de la misma UA. Siguiendo esta idea, las hojas de mayor tamaño y mayor relación ancho/longitud deberían corresponder a los entrenudos más largos de una UA y las hojas menores y más estrechas a sus entrenudos más cortos, con lo cual se optimizaría la utilización del espacio entre dos nudos. Los datos cuantitativos referidos a las UA de N. dombeyi o de otras especies de Nothofagus de que se dispone no apoyan esta hipótesis. Por el contrario, observaciones de gran número de UA de estas especies indican que los entrenudos correspondientes a las hojas de mayor tamaño de una UA son más cortos que los de posición más distal, que corresponden a hojas menores (J. Puntieri, datos no publicados). Una interpretación funcional de las tendencias en el tamaño foliar podría sugerir que la forma y el tamaño de una hoja quedarían determinados por el suministro de nutrientes al momento de diferenciarse el primordio correspondiente a partir del meristema. Estudios sobre diferenciación foliar apoyan esta interpretación (Steeves & Sussex, 1989; Van Volkenburgh, 1999). Esta hipótesis debería ser testeada mediante ensayos in vitro en los que se varíe la disponibilidad de recursos para los primordios en formación.
En las más largas UA de N. dombeyi de este estudio, así como en UA largas de otras especies de Nothofagus estudiadas con anterioridad (Puntieri et al., 2001; Stecconi et al., 2004), las variaciones en el tamaño y la forma de las hojas distales según su posición sobre el tallo son más irregulares que las observadas para hojas más proximales de las mismas UA (Fig. 2 y 3). Esta diferencia sería el resultado del carácter preformado de la porción proximal y el carácter neoformado de la porción distal de las UA más largas, lo cual distingue a estas UA de aquellas de tamaño intermedio o corto, enteramente preformadas. Si bien no se dispone de datos cuantitativos de preformación y neoformación para los árboles incluidos en el presente estudio, datos sobre otras poblaciones de árboles de N. dombeyi similares en edad y tamaño a los del presente estudio apoyan esta hipótesis (Puntieri et al., 2000). Mientras que todas las hojas preformadas de una UA resultan de un evento de diferenciación (al final de una estación de crecimiento) y uno de extensión (al inicio de la estación de crecimiento siguiente; García et al., 2006), las hojas neoformadas resultarían de eventos de diferenciación y extensión sucesivos e independientes (Puntieri et al., 2002). Asumiendo esta diferencia, podría esperarse mayor influencia ambiental y, en consecuencia, tamaños y formas foliares más heterogéneos en la neoformación que en la preformación.
El área foliar en el contexto de la unidad de alargamiento
Las hojas son consideradas órganos estrechamente relacionados con la respuesta plástica (no debida a variaciones genéticas) de las plantas a diferentes ambientes (Givnish, 1988). En el caso de los árboles, el estudio de la plasticidad foliar debe tomar en cuenta el contexto arquitectural en el cual es producida cada hoja. Al analizar las características de las hojas en función de las UA y los ejes en los que son producidas, es posible arribar a una mejor comprensión de la relación entre los desarrollos foliar y caulinar de una planta.
El análisis de las relaciones dimensionales de tallos y hojas de las UA de cada una de las tres categorías de ejes identificadas en el presente estudio (tronco, ramas principales y ramas cortas) permite inferir la función que cumple cada una de estas categorías. El tronco presenta UA con bajas relaciones entre la longitud y el peso del tallo y entre el área foliar y el volumen del tallo. En contraste, las UA de las ramas cortas presentan elevadas relaciones longitud/peso del tallo y área foliar/volumen del tallo. Dados estos resultados, podría concluirse que el tronco estaría más capacitado que las ramas cortas para soportar la carga estática debida a su propio peso y el de sus ramas, así como la carga dinámica debida al viento (Niklas, 1994; Anten et al., 2005). De manera que el tronco sería un eje con función predominantemente exploratoria del espacio vertical, y de localización y soporte en ese espacio de las otras categorías de ejes. Debido a su bajo desarrollo caulinar, las ramas cortas serían ejes principalmente asimilatorios. El bajo número de años de permanencia de cada rama corta en esta especie (Stecconi, 2006) se vincularía con la elevada relación longitud/peso caulinar de las mismas. Las UA de las ramas principales son intermedias entre aquellas del tronco y las de las ramas cortas en términos de desarrollo foliar y caulinar. Esto concuerda con la doble función de las ramas principales que, por un lado, exploran el espacio horizontal y sostienen a las ramas cortas en ese espacio, y por otro despliegan sus hojas en el espacio horizontal que ocupan.
En varias especies arbóreas es posible identificar una categoría de ejes de desarrollo caulinar aún menor que el observado en las ramas cortas de N. dombeyi (Macdonald & Mothersill, 1983; Nicolini & Chanson, 1999). Tal es el caso de las ramas cortas de N. pumilio, cuyas UA suelen no superar los 2-3 mm de longitud (Puntieri et al., 2003; Stecconi, 2006). La ausencia de ejes de tan escaso desarrollo caulinar en N. dombeyi se relacionaría con su carácter perennifolio: cierto grado de alargamiento de entrenudos sería esencial para evitar la superposición de las hojas desarrolladas en UA sucesivas sobre el mismo eje.
Dadas estas diferencias estructurales entre las unidades que componen las diferentes categorías de ejes, podría plantearse la siguiente hipótesis: la traslocación de fotoasimilados en árboles juveniles de N. dombeyi se efectuaría desde las ramas cortas hacia las ramas principales y el tronco, y desde las ramas principales hacia el tronco, pero no en sentido inverso. Actualmente se encuentran en evaluación las propiedades fotosintéticas de hojas correspondientes a diferentes ejes de las especies de Nothofagus, lo cual permitirá progresar en la evaluación de esta hipótesis.
Nothofagus dombeyi en ambientes sombreado y soleado
Estudios ecológicos se refieren a N. dombeyi como una especie de baja a muy baja tolerancia a la sombra (Read & Hill, 1985; Donoso, 1992; Kitzberger, 1994; Donoso et al., 2006). Justifica este calificativo el hecho comprobado de que la regeneración masiva de esta especie a partir de semillas tiene lugar en áreas donde el dosel del bosque ha sido eliminado (Veblen et al., 1996). El clareo del dosel por caídas de ramas o árboles puede permitir el establecimiento de plántulas y renovales de N. dombeyi en el sotobosque, pero el crecimiento de los árboles que circundan el claro suele impedir que las plántulas de esta especie, de crecimiento lento en los primeros años, se establezcan en el dosel. Tal sería el caso de los ejemplares desarrollados a la sombra que se seleccionaron para el presente trabajo.
El desarrollo de N. dombeyi a la sombra afecta a las UA de las diferentes categorías de eje que componen su arquitectura. Si bien es posible categorizar los ejes de los árboles juveniles de N. dombeyi en base a sus características morfológicas, tanto en condiciones de alta como de baja radiación incidente, las diferencias entre estas categorías son más notables en la primera de estas situaciones. Concordantemente, en individuos juveniles de N. pumilio las diferencias morfológicas entre categorías de eje fueron menores para individuos desarrollados en condiciones de sombra que para aquellos desarrollados con alta disponibilidad de luz (Puntieri et al., 1999).
El nivel de radiación incidente sería proporcional al peso, la longitud y el número de hojas de las UA en N. dombeyi, en especial las del tronco y las ramas principales. Pero en cada categoría de ejes, el tallo de una UA es más largo en relación a su peso en ambiente sombreado que en ambiente abierto. Dado que la relación volumen/peso del tallo no es afectada por el ambiente lumínico, puede concluirse que la diferente relación longitud/peso según el ambiente lumínico responde a diferencias en el grosor del tallo y no en la densidad de los tejidos que lo componen.
La diferencia cuantitativamente más notable entre las UA desarrolladas bajo diferentes condiciones lumínicas concierne al área foliar específica (AFE). El AFE es una variable muy afectada por las condiciones en las cuales se desarrolla una hoja (Björkman, 1981; Bartelink, 1997; Wilson et al., 1999). Valores de AFE altos suelen estar asociados con elevadas tasa de fotosíntesis, crecimiento relativo y contenido de nitrógeno (Reich et al., 1998; Ackerly & Reich, 1999; Hanba et al., 1999; Jones, 2001). Si bien el aumento en el AFE en hojas desarrolladas a la sombra en comparación con hojas no sombreadas es una respuesta habitual en las plantas (Björkman, 1981), tal aumento se puede lograr por diferentes mecanismos según la especie. En muchas especies de plantas, bajas intensidades lumínicas inducen un incremento en el área foliar (James & Bell, 2000; Uemura et al., 2000; Niinemets et al., 2004), aunque también se han reportado menores área y biomasa foliares en esas condiciones (Duarte & Dillenburg, 2000). En N. dombeyi, la mayor AFE bajo sombra se alcanza con una menor biomasa foliar pero sin cambios notables en el área foliar. Es probable que la disminución en el peso foliar en respuesta a la sombra se relacione con la disminución del peso del tallo en relación a su longitud y, por lo tanto, con la capacidad de éste de sostener sus hojas en posición horizontal.
De manera que el desarrollo de N. dombeyi bajo condiciones de sombra involucra, para cada categoría de ejes, menor formación de tejidos de sostén en relación a la observada en ejemplares desarrollados en ambiente abierto. El desarrollo del área foliar bajo condiciones de sombra se incrementa pero solo al considerarla en términos relativos a la biomasa de hojas y tallos. Las diferencias estructurales relacionadas con diferentes disponibilidades de luz son similares a aquellas que se observan entre categorías de ejes desarrolladas bajo condiciones de alta disponibilidad de luz. Es decir que una UA del tronco de un ejemplar desarrollado a la sombra se asemeja a una UA de una rama principal de un ejemplar desarrollado en ambiente abierto, y una UA de una rama principal a la sombra a una UA de una rama corta en medio abierto. En vista de las funciones de cada categoría de ejes mencionadas arriba, el desarrollo de un árbol de N. dombeyi a la sombra implicaría cambios estructurales tendientes a aumentar la superficie asimilatoria en desmedro del desarrollo caulinar, con lo cual podría sobrevivir en el sotobosque pero con bajo incremento anual en altura.
La evaluación de la forma y el tamaño de las hojas, así como su funcionamiento en una especie debe tomar en cuenta las variaciones que se producen en los atributos foliares debido a gradientes relacionados con las categorías de ejes que constituyen los individuos y la modalidad de desarrollo de las unidades que componen cada eje.
Agradecimientos
Los autores agradecen a Eduardo Zattara por su colaboración en la digitalización de áreas foliares, a la Universidad Nacional del Comahue y al CONICET por el apoyo económico que hizo posible el desarrollo de este estudio, y a la Administración de Parques Nacionales por la autorización para realizar muestreos en áreas de su jurisdicción.
Bibliografía
1. ACKERLY, D. D. & P. B. REICH. 1999. Convergence and correlations among leaf size and function in seed plants: a comparative test using independent contrasts. Amer. J. Bot. 86: 1272-1281. [ Links ]
2. ANTEN, P. R., R. CASADO-GARCIA & H. NAGASHIMA. 2005. Effects of mechanical stress and plant density on mechanical characteristics, growth, and lifetime reproduction of tobacco plants. Amer. Naturalist 166: 650-660. [ Links ]
3. BARTELINK, H. H. 1997. Allometric relationships for biomass and leaf area of beech (Fagus sylvatica L.). Ann. Sci. Forest. 54: 39-50. [ Links ]
4. BARTHÉLÉMY, D. & Y. CARAGLIO. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann. Bot. 99: 375-407. [ Links ]
5. BARTHÉLÉMY, D., J. G. PUNTIERI, C. BRION, E. RAFFAELE, J. MARINO & P. MARTINEZ. 1999. Morfología de las unidades estructurales y modo de desarrollo básico de especies patagónicas de Nothofagus (Fagaceae). Bol. Soc. Argent. Bot. 34: 29-38. [ Links ]
6. BJÖRKMAN, O. 1981. Responses to different quantum flux densities. In: Lange O. L., P. S. Nobel, C. B. Osmond & H. Ziegler (eds.), Physiological Plant Ecology I. Responses to the Physical Environment, pp. 57-107. Springer-Verlag, Berlin. [ Links ]
7. CONTI, H. A. 1998. Características climáticas de la Patagonia. In: Correa, M. N. (ed.), Flora Patagónica, pp. 31-47. Colección Científica del INTA, Buenos Aires. [ Links ]
8. CRITCHFIELD, W. B. 1960. Leaf dimorphism in Populus trichocarpa. Amer. J. Bot. 47: 699-711. [ Links ]
9. DONOSO, C. 1992. Ecología Forestal. 3ra ed. Editorial Universitaria, Universidad Austral de Chile, Santiago. [ Links ]
10. DONOSO, P., C. DONOSO, C. NAVARRO & B. ESCOBAR. 2006. Nothofagus dombeyi (Mirb.) Oerst. Coihue, Coigüe. In: Donoso, C. (ed.), Las especies arbóreas de los bosques templados de Chile y Argentina. Autoecología, pp. 423-432. Marisa Cuneo Ediciones, Valdivia. [ Links ]
11. DUARTE, L. & L. R. DILLENBURG. 2000. Ecophysiological responses of Araucaria angustifolia (Araucariaceae) seedlings to different irradiance levels. Austral. J. Bot. 48: 531-537. [ Links ]
12. GARCÍA, S., J. G. PUNTIERI & G. VOBIS. 2006. Morfología y anatomía del ápice caulinar de Nothofagus dombeyi (Nothofagaceae) a lo largo de un año. Bol. Soc. Argent. Bot. 41: 13-21. [ Links ]
13. GIVNISH, T. J. 1982. On the adaptive significance of leaf height in forest herbs. Amer. Naturalist 120: 353-381. [ Links ]
14. GIVNISH, T. J. 1988. Adaptation to sun and shade: a whole-plant perspective. Austral. J. Plant Physiol. 15: 63-92. [ Links ]
15. GIVNISH, T. J. & G. J. VERMEIJ. 1976. Sizes and shape of liane leaves. Amer. Naturalist 110: 743-778. [ Links ]
16. GROSFELD, J. 2002. Análisis de la variabilidad morfológica y arquitectural de Austrocedrus chilensis (D. Don) Pic. Serm. et Bizzarri, Fitzroya cupressoides (Molina) I. M. Johnst., Pilgerodendron uviferum (D. Don) Florin y Cupressus sempervirens L. (Cupressaceae). Tesis doctoral, Universidad Nacional del Comahue, Bariloche. [ Links ]
17. HALLÉ, F., R. OLDEMAN & P. TOMLINSON. 1978. Tropical Trees and Forests. Springer Verlag, Berlin. [ Links ]
18. HANBA, Y. T., S. I. MIYAZAMA & I. TERASHIMA. 1999. The influence of leaf thickness on the CO2 transfer conductance and leaf stable carbon isotope ratio for some evergreen tree species in Japanese warm-temperate forests. Funct. Ecol. 13: 632-639. [ Links ]
19. JAMES, S. A. & D. BELL. 2000. Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances. Tree Physiol. 20: 1007-1018. [ Links ]
20. JONES, C. S. 2001. The functional correlates of heteroblastic variation in leaves: Changes in form and ecophysiology with whole plant ontogeny. Bol. Soc. Argent. Bot. 36: 171-182. [ Links ]
21. KITZBERGER, T. 1994. Fire regime variation along a northern Patagonian forest-steppe gradient: stand and landscape response. Tesis Doctoral, University of Colorado, Boulder. [ Links ]
22. KOZLOWSKI, T. T. & S. G. PALLARDY 1997. Physiology of Woody Plants. 2nd ed. Academic Press, San Diego. [ Links ]
23. MACDONALD, A. D. & D. H. MOTHERSILL. 1983. Shoot development in Betula papyrifera. I. Short-shoot organogenesis. Canad. J. Bot. 61: 3049-3065. [ Links ]
24. NICOLINI, E. & B. CHANSON. 1999. La pousse courte, un indicateur du degré de maturation chez le hêtre (Fagus sylvativa L.). Canad. J. Bot. 77: 1539-1550. [ Links ]
25. NIKLAS, K. J. 1994. Plant allometry. The scaling of form and process. The University of Chicago Press, Chicago & London. [ Links ]
26. NIINEMETS, Ü., A. CESCATTI & R. CHRISTIAN. 2004. Constraints on light interception efficiency due to shoot architecture in broad-leaved Nothofagus species. Tree Physiol. 24: 617-630. [ Links ]
27. PARKHURST, D. F. & O. L. LOUCKS, 1972. Optimal leaf size in relation to environmental. J. Ecol. 60: 505-537. [ Links ]
28. PUNTIERI, J. G., D. BARTHÉLÉMY, P. MARTÍNEZ, E. RAFFAELE & C. BRION. 1998. Annual-shoot growth and branching patterns in Nothofagus dombeyi (Fagaceae). Canad. J. Bot. 76: 673-685. [ Links ]
29. PUNTIERI, J. G., E. RAFFAELE, P. MARTÍNEZ, D. BARTHÉLÉMY & C. BRION. 1999. Morphological and architectural features of young Nothofagus pumilio (Poepp. & Endl.) Krasser (Fagaceae). Bot. J. Linn. Soc. 130: 395-410. [ Links ]
30. PUNTIERI, J. G., M. DAMASCOS & M. S. SOUZA. 2001. Tendencias ontogenéticas en el tamaño y la forma de las hojas de Nothofagus pumilio (Poepp. et Endl.) Krasser (Fagaceae). Ecol. Austral 11: 105-114. [ Links ]
31. PUNTIERI, J. G., M. S. SOUZA, D. BARTHÉLÉMY, C. BRION, M. NÚÑEZ & C. MAZZINI. 2000. Preformation, neoformation and shoot structure in Nothofagus dombeyi (Nothofagaceae). Canad. J. Bot. 78: 1044-1054. [ Links ]
32. PUNTIERI, J. G., D. BARTHÉLÉMY, C. MAZZINI & C. BRION. 2002. Periods of organogenesis in shoots of Nothofagus dombeyi (Mirb.) Oersted (Nothofagaceae). Ann. Bot. 89:115-124. [ Links ]
33. PUNTIERI, J. G., M. S. SOUZA, C. BRION, C. MAZZINI & D. BARTHÉLÉMY. 2003. Axis differentiation in two South American Nothofagus species (Nothofagaceae). Ann. Bot. 92: 589-599. [ Links ]
34. PUNTIERI, J. G., J. E. GROSFELD, M. STECCONI, C. BRION & D. BARTHÉLÉMY. 2007. Bud and growth unit structure in seedlings and saplings of Nothofagus alpina (Nothofagaceae). Amer. J. Bot. 94: 1382-1390. [ Links ]
35. READ J. & R. HILL 1985. Photosynthetic responses to light of Australian and Chilean species of Nothofagus and their relevance to the rainforest dynamics. New Phytol. 101: 731-742. [ Links ]
36. REICH, P. B., D. S. ELLSWORTH & B. WALTERS. 1998. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relation: evidence from within and across species and functional groups. Funct. Ecol. 12: 948-958. [ Links ]
37. SCHAFFALITZKY de MUCKADELL, M. 1959. Juvenile stages in woody plants. Physiol. Pl. 7: 782-796. [ Links ]
38. COPPA, C. O. 1998. Los suelos. In: Correa, M. N. (ed.), Flora Patagónica, pp. 15-30. Colección Científica del INTA, Buenos Aires. [ Links ]
39. SOKAL, R. R. & F. J. ROHLF 1981. Biometry. 2nd ed. Freeman and Co., New York. [ Links ]
40. STECCONI, M., P. MARCHELLI, J. G. PUNTIERI, P. PICCA & L. GALLO. 2004. Natural hybridization between a deciduous (Nothofagus antarctica, Nothofagaceae) and an evergreen (N. dombeyi) forest tree species: evidence from morphological and isoenzymatic traits. Ann. Bot. 94: 75-786. [ Links ]
41. STECCONI, M. 2006. Variabilidad arquitectural de especies nativas de Nothofagus de la Patagonia (N. antarctica, N. pumilio, N. dombeyi). Tesis Doctoral, Universidad Nacional del Comahue, Bariloche. [ Links ]
42. STEEVES, T. A. & I. M. SUSSEX. 1989. Patterns in Plant Development, 2nd edition. Cambridge University Press, Cambridge. [ Links ]
43. TOURN, M., D. BARTHÉLÉMY & J. GROSFELD. 1999. Una aproximación a la arquitectura vegetal: conceptos, objetivos y metodología. Bol. Soc. Argent. Bot. 34: 85-99. [ Links ]
44. UEMURA, A., A. ISHIDA, T. NAKANO, I. TERASHIMA, H. TANABE & Y. MATSUMOTO. 2000. Acclimation of leaf characteristics of Fagus species to previous-year and current-year solar irradiance. Tree Physiol. 20: 945951 [ Links ]
45. VAN VOLKENBURGH E. 1999. Leaf expansion - an integrating plant behavior. Pl. Cell Environ. 22: 14631473. [ Links ]
46. VEBLEN, T. T. 1989. Nothofagus regeneration in treefall gaps in northern Patagonia. Canad. J. For. Res. 19: 365-371. [ Links ]
47. VEBLEN, T. T., C. DONOSO, T. KITZBERGER & A. REBERTUS. 1996. Ecology of Southern Chilean and Argentinean Nothofagus forests. In: Veblen, T. T., R. S. Hill & J. Read (eds.), The ecology and biogeography of Nothofagus forests, pp. 293-353. Yale University Press, New Haven. [ Links ]
48. WILSON, P. J., K. THOMPSON & J. G. HODGSON. 1999. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 143: 155-162. [ Links ]
Recibido el 30 de Mayo de 2007
Aceptado el 14 de Noviembre de 2007.

