










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.43 n.1-2 Córdoba ene./jul. 2008
Comunidades de malezas en cultivos hortícolas en la Provincia de Neuquén (Argentina)
María Beatriz Cerazo1 y Luisa Conticello1
1Depto. de Biología Aplicada. Facultad de Ciencias Agrarias. Universidad Nacional del Comahue. Casilla de Correos 85. C.P. 8303.Cinco Saltos, Río Negro. Argentina.
Resumen: La gran mayoría de los cultivos hortícolas son muy sensibles a la competencia de malezas, las cuales inciden en los rendimientos y calidad de los mismos y en ocasiones determinan la pérdida total del cultivo. El presente trabajo tuvo como objetivo conocer la composición florística de las comunidades de malezas y su análisis sintaxonómico, ya que ambos constituyen un buen parámetro para medir el estado de los cultivos. La vegetación fue muestreada siguiendo la metodología de Braun Blanquet, en explotaciones de seis localidades de la provincia. A partir de esta información se determinaron ocho asociaciones, de las cuales seis están comprendidas en la clase Stellarietea mediae (Br. Bl. 1931) Tx., Prsg. et Lohm, 1950 y las dos restantes en Artemisietea vulgaris Lohm, Prsg et Tüxen 1950. El deficiente estado del cultivo está señalado por el desarrollo de malezas perennes de difícil control, incluidas en su mayoría en la clase Artemisietea vulgaris. Se inventariaron ochenta y nueve especies, para los periodos otoño- invierno y primavera - verano. Existe un claro predominio de Dicotiledóneas (79 %) sobre Monocotiledóneas y de malezas anuales (66 %) sobre perennes.
Palabras claves: Malezas; Sintaxonomía; Hortalizas; Neuquén.
Summary: Weed's communities in vegetable crops in Neuquen Province, (Argentina). Most vegetable crops are very sensitive to the weed competition, which affects the yields and quality and sometimes the total lost of the crop. The objective of this study was to evaluate the floristic composition of the weed communities and to analyse its sintaxonomy, because both are effective parameters for measuring the state of the crop. The vegetation was sampled following the Braun-Blanquet methodology on farms in six localities in the province. We determined eight associations from this information: six are included in the class Stellarietea mediae (Br. Bl. 1931) Tx., Prsg. et Lohm, 1950 and the other two in Artemisietea vulgaris Lohm, Prsg and Tüxen 1950. The deficient state of the crop resulted from the development of perennial and difficult to control weeds, most of them included in the class Artemisietea vulgaris. Eighty nine species were tacked stock for the autumn -winter and spring - summer periods. There was a clear predominance of Dicotyledonous ( 79 %) over Monocotyledonous and a predominance of annual weeds (66%) over perennial weeds.
Key words: Weeds; Syntaxonomy; Vegetables; Neuquén.
Introducción
El laboreo continuo del suelo para la producción hortícola, ha favorecido la selección de genotipos de malezas oportunistas y muy competitivas que actúan sobre la calidad y rendimiento de estos cultivos. Entre la especie cultivada y las malezas, se establece una activa competencia por agua, luz, nutrientes y espacio, que afecta en forma decisiva su potencial productivo. Además existe la posibilidad que muchas de ellas sean hospedantes de organismos patógenos entre los que se destacan los virus y sus insectos transmisores (Nobile et al. 1989).
En Europa el estudio de las comunidades de malezas ha alcanzado gran desarrollo, se establecieron correlaciones entre estas comunidades y determinadas practicas culturales (Thomas & Dale 1991, Thomas et al. 1996, Colbach et al. 2000) como también con algunas características de los suelos (Andreasen & Striebig 1991). Existen estudios que permiten predecir la abundancia de malezas basados en el análisis isoclimático (Panetta & Mitchell 1991) y otros estudios sobre su dinámica y sucesión (Pysek& Pysek 1991, Cousens & Mortimer 1995, Gardener et al 1999). En Argentina existen numerosos antecedentes sobre el estudio de comunidades de malezas en distintos cultivos (Méndez 1983, 1984, 1990, 2002, Suarez et al 1997, Ahumada & Alvarez 2001, Medina et al 2004).
La producción hortícola en Argentina ha tenido un gran crecimiento frente a otros rubros productivos tradicionales. La provincia de Neuquén no es ajena a esta tendencia, ya que se verifica un aumento en la superficie provincial que pasa de 371 ha (Censo Frutihortícola de la Provincia del Neuquén 1994) a 1.518 ha de hortalizas(Censo Nacional Agropecuario 2002).
Debido a la importancia de las malezas en los cultivos hortícolas se continuó con estudios florísticos, fitosociológicos y sintaxonómicos de la vegetación arvense (Conticelllo & Gandullo 1991, Conticello et al. 1998, 2001, 2002; Cerazo et al. 2000).
El presente trabajo formó parte de un Proyecto de Investigación cuyo objetivo principal fue el estudio de las malezas presentes en cultivos hortícolas en la provincia de Neuquén y su relación con algunas practicas culturales.
Material y Método
Las localidades de Añelo y San Patricio del Chañar pertenecen al departamento de Añelo; Centenario, Plottier y Senillosa al de Confluencia y Picún Leufú al del mismo nombre. Todos estos departamentos están incluidos en el Area Fisiográfica Oriental.
Dicha área se caracteriza por la presencia de extensos pedimentos que suavemente se inclinan hacia el río Limay. Los suelos están desarrollados sobre rocas sedimentarias, cubiertas por rodados o gravas de pedimento cementados por carbonato de calcio. Son predominantemente pardos grises desérticos. La cantidad de humus es mínima por lo que los cultivos son factibles solo en los lugares en que la textura es fina, como ocurre en las llanuras aluviales de los ríos Neuquén y Limay.
Se han formado suelos azonales, sin diferenciación de horizontes, de texturas gruesas, baja retención de humedad, baja fertilidad y erosionables. Se destaca la presencia de regosoles, con presencia de carbonatos en todo el perfil, condicionando el pH algo alcalino (7,5-8,5) (Atlas de la provincia de Neuquén 1982).
La aridez es el factor que define el área, la cual esta limitada al occidente por la isohieta de 150 mm. Según la clasificación de Thornthwaite (1948) el clima de la región es árido, mesotermal con un déficit hídrico que cual oscila entre los 500-550 mm anuales (Papadakis, 1980). La temperatura media anual es de 14°C, con una máxima absoluta que sobrepasa los 40°C y una mínima absoluta de -13°C. La temperatura media del mes más caluroso (enero) es de 22°C y la del mes más frío (julio), de 5,5°C. La frecuencia media anual de heladas varía entre 52 a 74 días, ocurriendo desde abril a octubre, siendo el período libre de heladas de aproximadamente 184 días al año. La zona se caracteriza por ser muy ventosa sobre todo en primavera, con vientos del sector Oeste - Sudoeste. (Servicio Meteorológico Nacional, 1992).
Las observaciones se realizaron en dieciséis establecimientos dedicados a cultivos hortícolas tales como cebolla, ajo, lechuga, tomate, entre otros, conducidos bajo condiciones de riego artificial. Los campos seleccionados corresponden a pequeños y medianos productores, con una extensión entre 1 y 10 hectáreas. Los mismos se distribuyen en seis localidades de la provincia de Neuquén: Añelo, Centenario, San Patricio del Chañar, Picún Leufú, Senillosa y Plottier (Figura 1). El trabajo de campo se inició en enero de 2004 hasta enero de 2005. Se realizaron 12 campañas, una cada 30 días. El total de relevamientos fue de 190, siguiendo la metodología de Braun Blanquet (1979) actualizada por Géhu & Rivas Martinez (1982). Los relevamientos se realizaron en parcelas de 20 m2 y para cada especie relevada se consideró su cobertura y presencia. La información obtenida se volcó en tablas iniciales para los períodos otoño - invierno y primavera - verano, a fin de registrar las variaciones estacionales en la composición florística y parámetros considerados. Las formas de vida se determinaron según Raunkiaer (1934) y con ellas se elaboró el espectro biológico, basado en la abundancia y cobertura de las especies. La comparación de las tendencias seguidas por esos tipos biológicos permitió evaluar el desarrollo de las comunidades.
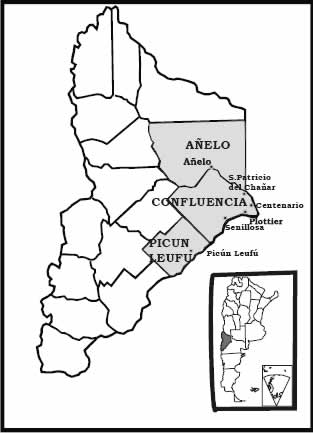
Fig. 1. Ubicación relativa del área de estudio en Neuquén, Argentina.
Se confeccionaron tablas comparativas sintéticas de relevamientos, para otoño - invierno (86 censos) y primavera - verano (102 censos) con los valores de la escala de presencia y en base a ellas se realizó el análisis sintaxonómico de las mismas. El menor número de censos en el primer periodo del año es consecuencia del anegamiento de algunos sitios que impidió la ejecución de los mismos.
Para la normalización de los sintaxones se siguió el Código Internacional de Nomenclatura Fitosociológica (Barckman et al. 1986, Weber et al. 2000).
Los datos de cobertura de las especies fueron analizados usando el programa P-Cord (Mc Cune & Mefford, 1999) para lo cual se trabajó con una matriz que incluyó los censos correspondientes a las periodos otoño - invierno y primavera - verano respectivamente. Los resultados preliminares de estos dendrogarmas permiten detectar grupos florísticos. Estas matrices se someten a un análisis de clasificación numérico mediante el método de la mínima varianza, usando la distancia relativa euclídea como parámetro de similitud (Ward 1963, Anderberg 1973, Orloci 1978).
Resultados y Discusión
Se inventariaron 89 especies, distribuidas en 24 familias y 74 géneros. Las familias más importantes por el número de especies fueron Asteráceas y Poáceas, tal como ocurre con las floras exóticas a nivel mundial (Pauchard & Alaback 2004). Existe un claro predominio de Dicotiledóneas (79 %) sobre Monocotiledóneas y de malezas anuales (66 %) sobre perennes (Tabla 1). El alto porcentaje de especies exóticas (76 %) es indicador de la presión antrópica a que están sometidos los sitios destinados a cultivos hortícolas (Hauenstein et al. 2002). Las especies comunes a todos las localidades estudiadas fueron: Amaranthus quitensis, Chenopodium album, Cichorium intybus, Capsella bursa - pastoris y Sonchus oleraceus, lo que concuerda con la gran amplitud ecológica que las caracteriza.
Tabla 1. Especies de malezas inventariadas en el área de estudio.
| Especies | Familia | Cobertura | F V | Especies | Familia | Cobertura | F V | ||
|---|---|---|---|---|---|---|---|---|---|
| O-I | P-V | O-I | P-V | ||||||
| Amaranthus quitensis | Amarantáceas | 244,5 | 656 | Te | Lamium amplexicaule | Lamiáceas | 143 | 47 | Te |
| Ambrosia tenuifolia | Asteráceas | 1,5 | 0 | Cr | Lolium multiflorum | Poáceas | 6 | 0 | Te |
| Anoda cristata | Malváceas | 1,5 | 0 | Te | Malva parviflora | Malváceas | 9,5 | 0 | Te |
| Anthemis cotula | Asteráceas | 14 | 11 | Te | Marrubium vulgare | Lamiáceas | 2 | 0 | He |
| Artemisia verlotorum | Asteráceas | 10,5 | 35,5 | Cr | Medicago lupulina | Fabáceas | 1,5 | 3,5 | Te |
| Brassica nigra | Brasicáceas | 0,5 | 0 | Te | Melilotus albus | Fabáceas | 24,5 | 8,5 | Te |
| Bromus brevis | Poáceas | 0 | 10 | Te | Mentha piperita | Lamiáceas | 7 | 0 | Cr |
| Bromus tectorum | Poáceas | 0 | 6 | Te | Oenothera stricta | Onagráceas | 0,5 | 0 | Te |
| Bromus catharticus | Poáceas | 13 | 24 | Te | Oxalis corniculata | Oxalidáceas | 0,5 | 0 | Te |
| Calystegia sepium | Convolvuláceas | 0 | 40 | Cr | Panicum capillare | Poáceas | 30 | 331 | Te |
| Capsella bursa-pastoris | Brasicáceas | 174 | 77 | Te | Phyla canescens | Verbenáceas | 0,5 | 0 | He |
| Cardaria draba | Brasicáceas | 204 | 294 | Cr | Plantago lanceolata | Plantagináceas | 32 | 37 | He |
| Cenchrus pauciflorus | Poáceas | 14 | 0 | Te | Plantago major | Plantagináceas | 2 | 0 | He |
| Centaurea calcitrapa | Asteráceas | 4,5 | 5 | Te | Polygonum aviculare | Poligonáceas | 80,5 | 194 | Te |
| Centaurea melitensis | Asteráceas | 50,5 | 155 | Te | Polygonum persicaria | Poligonáceas | 3 | 40 | Te |
| Chenopodium album | Quenopodiáceas | 255 | 625 | Te | Portulaca oleracea | Portulacáceas | 12,5 | 247,5 | Te |
| Chenopodium ambrosioides | Quenopodiáceas | 26 | 11 | Te | Raphanus sativus | Brasicáceas | 5 | 0 | Te |
| Chenopodium cordobense | Quenopodiáceas | 4 | 103 | Te | Rapistrum rugosum | Brasicáceas | 64,5 | 33 | Te |
| Chenopodium hircinum | Quenopodiáceas | 0 | 49 | Te | Rumex crispus | Poligonáceas | 6 | 16 | He |
| Chenopodium multifidum | Quenopodiáceas | 1,5 | 0 | He | Schismus barbatus | Poáceas | 1 | 0 | Te |
| Cichorium intybus | Asteráceas | 52,5 | 53,5 | He | Senecio vulgaris | Asteráceas | 33 | 9 | Te |
| Cirsium vulgare | Asteráceas | 6 | 13 | Te | Setaria parviflora | Poáceas | 126,5 | 163 | Cr |
| Conium maculatum | Apiáceas | 23,5 | 33 | Te | Setaria verticillata | Poáceas | 32,5 | 130 | Te |
| Convolvulus arvensis | Convolvuláceas | 223,5 | 395 | Cr | Sisymbrium irio | Brasicáceas | 111 | 147 | Te |
| Cuscuta indecora | Cuscutáceas | 1,5 | 0 | Te | Solidago chilensis | Asteráceas | 1 | 0 | Cr |
| Cynodon dactylon | Poáceas | 181,5 | 253 | Cr | Sonchus asper | Asteráceas | 8 | 0 | Te |
| Cyperus rotundus | Ciperáceas | 11 | 0 | Cr | Sonchus oleraceus | Asteráceas | 47 | 24,5 | Te |
| Datura ferox | Solanáceas | 2,5 | 0 | Te | Sorghum halepense | Poáceas | 1 | 40 | Cr |
| Descurainia argentina | Brasicáceas | 0,5 | 0 | Te | Spergula ramosa | Cariofiláceas | 3,5 | 7,5 | Te |
| Digitaria sanguinalis | Poáceas | 171,5 | 180,5 | Te | Sphaeralcea mendocina | Malváceas | 0,5 | 0 | He |
| Diplotaxis tenuifolia | Brasicáceas | 1 | 4 | He | Stellaria media | Cariofiláceas | 36,5 | 75 | Te |
| Distichlis spicata | Poáceas | 29,5 | 15 | Cr | Tagetes minuta | Asteráceas | 5 | 0 | Te |
| Echinochloa colonum | Poáceas | 72 | 186 | Te | Taraxacum officinale | Asteráceas | 5,5 | 14 | He |
| Echinochloa crusgalli | Poáceas | 26,5 | 74 | Te | Tessaria absinthioides | Asteráceas | 4 | 5,5 | Cr |
| Eragrostis lugens | Poáceas | 42 | 60,5 | He | Tribulus terrestris | Zigofiláceas | 0,5 | 0 | Te |
| Eragrostis virescens | Poáceas | 8 | 245 | Te | Trifolium repens | Fabáceas | 3 | 0 | Cr |
| Erodium cicutarium | Geraniáceas | 3 | 0 | Te | Urtica urens | Urticáceas | 175 | 25,5 | Te |
| Eruca sativa | Brasicáceas | 40 | 0 | Te | Verbascum thapsus | Escrofulariáceas | 0,5 | 0 | Te |
| Galinsoga parviflora | Asteráceas | 22 | 78 | Te | Verbascum virgatum | Escrofulariáceas | 0,5 | 0 | Te |
| Glycyrrhiza astragalina | Fabáceas | 30 | 184 | Cr | Verbena bonariensis | Verbenáceas | 1 | 0 | He |
| Gnaphalium montevidense | Asteráceas | 0,5 | 0 | He | Veronica didyma | Escrofulariáceas | 134 | 78 | Te |
| Hoffmannseggia erecta | Fabáceas | 38,5 | 2,5 | Cr | Wedelia glauca | Asteráceas | 1 | 0 | Cr |
| Hordeum murinum | Poáceas | 2 | 33,5 | Te | Xanthium cavanillesii | Asteráceas | 3,5 | 24 | Te |
| Kochia scoparia | Quenopodiáceas | 1,5 | 52 | Te | Xanthium spinosum | Asteráceas | 2,5 | 13 | Te |
| Lactuca serriola | Asteráceas | 10 | 22 | Te | |||||
Referencias: Cobertura: suma de los porcentajes de coberturas estimadas para cada especie en el total de los relevamientos; O-I: otoño - invierno; P-V: primavera - verano; F-V: formas de vida; Cr: criptófitas; He: hemicriptófitas; Te: terófitas.
En aquellos sitios de muestreo en que las labores culturales fueron incompletas o en aquellos cultivos que fueron abandonados, se relevó el mayor número de especies perennes y agresivas tales como Cyperus rotundus, Distichlis spicata, Cynodon dactylon, Cardaria draba, entre otras. En contraposición, en aquellos cultivos que se desarrollan en suelos profundos, bien drenados, con fertilización nitrogenada y labores culturales adecuadas y oportunas son predominantes Urtica urens, Lolium multiflorum, Amaranthus quitensis, Capsella bursa - pastoris, Portulaca oleracea y Galinsoga parviflora.
El análisis del espectro biológico (Fig. 2) permite establecer que las terófitas fueron las especies más abundantes y de mayor cobertura durante todo el año, en los dieciséis sitios estudiados. Poseen varias germinaciones en el año, característica que sumada a su elevada producción de semillas y plasticidad genética determina que sean las especies que más rápidamente se adaptan a los cambios de ecosistemas inestables y altamente modificados (Shaltout & El Fahar 1991). Las criptófitas y hemicriptófitas caracterizan a los cultivos abandonados en el que estas formas biológicas dominan completamente los sitios en que hay ausencia de labores culturales (Méndez 1983, 1990).
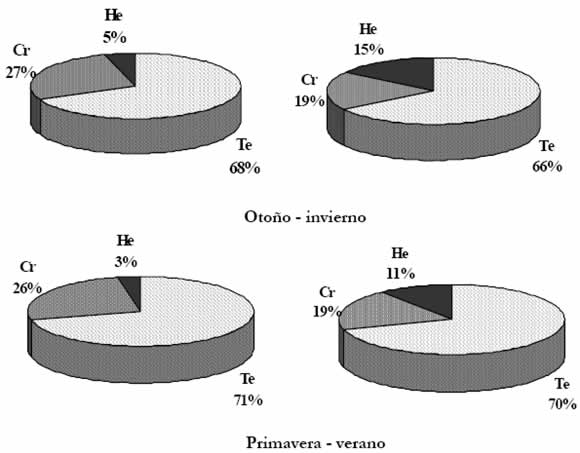
Fig. 2. Espectro biológico de la vegetación estudiada, según número de especies y cobertura. Referencias: Cr, criptófitas; He, hemicriptófitas; Te, terófitas.
Sintaxonomía
Por sus afinidades fisonómicas, florísticas y estructurales las comunidades presentes en los sitios estudiados se organizan de acuerdo al siguiente esquema sintaxonómico (Géhu et al. 1983).
I. Clase Stellarietea mediae (Br. Bl. 1931) Tx. Prsg. et Lohm 1950. 1.Orden Wedeletalia glaucae Méndez 1983
1.1. Alianza Wedelion glaucae Méndez 1983
1.1.1. Asociación Cynodetum dactyli Méndez 1983
2. Orden Chenopodietalia albi Tx. et Lohm 1950
2.1. Alianza Sisymbrion Méndez 1983
2.1.1. Asociación Sisymbrio - Rapistretum Méndez 1983
2.1.2. Comunidad de Urtica urens.
3. Orden Solano-nigri-Polygonetalia convolvuli (Sissingh ex Westhoff, Dijk et Passier 1946) Bolòs 1962
3.1. Alianza Polygono convolvuli - Chenopodion polyspermi Koch ex Sisingh 1946 en Müller et Oberd in Oberd 1983
3.1.1. Asociación Setario - Echinochloetum coloni A. et O. Bolòs ex O. Bolòs 1956
3.1.2. Asociación Setario - Echinochloetum cruris-gallii Peinado, Bartolomé et M. Parras 1985.
4. Orden Sisymbrietalia officinalis J. Tx in Lohm et al. 1962
4.1. Alianza Hordeion leporini Br. Bl. (1931) 1947.
4.1.1. Comunidad de Stellaria media.
II.Clase Artemisietea vulgaris Lohm, Prsg et Tx 1950
Subclase Agropyretalia repentis Oberd, Th, Müller et Görs in Oberd 1967.
5.Orden Convolvulo arvensis - Agropyrion repentis Görs 1966.
5.1.1. Asociación Cardario drabae - Agropyretum repentis Th, Müller et Görs 1969 Subclase Onopordenea acanthii Rivas Mart., Báscones, Díaz, Fernández González et Loidi 1991
6. Orden Carthametalia lanati Brullo in Brullo et Marcenó 1985
6.1.1 Comunidad de Centaurea melitensis
Descripción de las comunidades
El análisis sintaxonómico permitió establecer ocho asociaciones, de las cuales seis están comprendidas en la clase Stellarietea mediae (Br. Bl. 1931) Tx., Prsg. et Lohm, 1950 y las dos restantes en Artemisietea vulgaris Lohm, Prsg et Tx 1951.
La primera clase está constituida por vegetación herbácea y ruderal de origen antrópico, con especies anuales, bienales y algunas perennes. Sus asociaciones son características de suelos ricos en nitrógeno y están ampliamente distribuidas en nuestro país, ya que pueden desarrollarse tanto en suelos sueltos como compactados, con escaso o abundante contenido de materia orgánica.
La Clase Artemisietea vulgaris comprende vegetación nitrófila, formada por plantas herbáceas, vivaces, que crecen en suelos alterados, generalmente profundos y con buena humedad. Algunas de las asociaciones pertenecientes a esta clase son abundantes en hemicriptófitos y criptófitos y señalan con su desarrollo a cultivos en los que las labores culturales no fueron las adecuadas y terminaron por ser abandonados.
Las comunidades presentes en la zona de estudio se ordenaron en tablas comparativas sintéticas (Tablas 2 y 3) según grados crecientes de abandono de los cultivos. La eficiencia en la realización de las prácticas culturales determina no solamente el rendimiento del cultivo, sino también el tipo de comunidades de malezas y principalmente su composición florística (Mortimer 1996).
Tabla 2. Cuadro comparativo sintético de relevamientos para las comunidades de malezas del período otoño-invierno. Se expresan los grados de constancia: V, presente en mas del 81 % de los censos; IV, 61-80 %; III, 41-60%; II, 21-40%; I, menos del 20% (Braun Blanquet).

Especies accidentales Sphaeralcea mendocina en G5; Sorghum halepense, Oenothera stricta, Gnaphalium montevidense, Verbascum thapsus,Verbascum virgatum en G9; Brassica nigra en G1; Oxalis corniculata en G4; Tribulus terrestris y Descurainia argentina en G2; Phyla canescens en G7.
Tabla 3. Cuadro comparativo sintético de relevamientos para las comunidades de malezas del período primavera-verano. Se expresan los grados de constancia:V, presente en mas del 81 % de los censos; IV, 61-80 %; III, 41-60%; II, 21-40%; I, menos del 20% (Braun Blanquet).

Las tres primeras comunidades que figuran en la Tabla 2: Sisymbrio - Rapistretum, comunidad de Stellaria media y comunidad de Urtica urens, permiten establecer que los cultivos en los cuales se desarrollan se caracterizan porque sus labores fueron adecuadas y oportunas (Méndez 1983, Conticello et al, en prensa).
La asociación Sisymbrio - Rapistretum está formada por especies anuales con altos valores de constancia. Es muy frecuente en todas las áreas cultivadas del país y de la zona; los terófitos son muy abundantes pero cada especie por sí sola presenta escasa cobertura. Las especies características y acompañantes tales como Sonchus oleraceus, Amaranthus quitensis, Chenopodium album y Capsella bursa - pastoris, entre otras actúan como pioneras de sitios altamente disturbados como los cultivos hortícolas que requieren de labores culturales continuas.
La comunidad de Stellaria media no se la incluyó en el Sisymbrio - Rapistretum por presentar como especie diferencial a Senecio vulgaris con altos valores de constancia. No se le dio rango sintaxonómico por considerar que requiere de estudios posteriores más detallados, al igual que la comunidad de Urtica urens. Los altos valores de cobertura de esta especie son indicadores de cultivos ricamente abonados ya que es muy exigente en nitrógeno, lo cual es confirmado por la presencia de Malva parviflora, Chenopodium multifidum y Lolium multiflorum (Rivas - Martínez 1978, Méndez 1990, Conticello et al., en prensa).
Las tres comunidades descriptas son de ciclo estival y a pesar de ello persisten durante el periodo otoño invernal. La explicación de esto se fundamenta en las particulares características climáticas del periodo 2004- 2005, en el que las temperaturas invernales no fueron tan rigurosas asociado a una mayor ocurrencia de precipitaciones.
Las asociaciones Setario - Echinochloetum coloni y Setario - Echinochloetum cruris gallii están formadas por especies cuya altura varía entre 0,60 a 1m. Son típicas de cultivos bajo riego y su plasticidad les permite adaptarse a suelos de naturaleza química muy diversa. Las especies características y diferenciales poseen mayores valores de cobertura ya que en su mayoría presentan porte decumbente o bien forman matas de gran diámetro. Ambas asociaciones se comportan como vicariantes, ya que Setario - Echinochloetum cruris gallii reemplaza a Setario - Echinochloetum coloni cuando disminuye la humedad del suelo (Méndez 1990, Cerazo & Conticello 2006).
Marcan con su desarrollo el estado medio de los cultivos como consecuencia de que las labores culturales no fueron completas. Esta situación se ve agravada por tratarse de sitios con problemas de drenaje como lo denuncia la aparición de Plantago lanceolata, Cichoriun intybus y Polygonum aviculare. El predominio de terófitas y algunas hemicriptófitas en ambas asociaciones permite su fácil control con lo que se logra reducir la incidencia de las malezas en los cultivos.
Las dos últimas asociaciones de la Tabla 2, Cynodetum dactyli y Cardario drabae -Agropyretum repentis denotan con su desarrollo el deficiente estado del cultivo, ya que se encuentran invadidos por malezas rizomatosas o con raíces gemíferas que determinan la perdida total de la cosecha (Labrada 1996).
Cynodetum dactyli forma un pastizal de 0,20 0,30 m de altura con altos valores de cobertura. La presencia de Conium maculatum, Xanthium cavanillesii y Verbena bonariensis marcan una mayor disponibilidad de agua y nitrógeno, localizándose en la periferia del gran manchón que forma este tipo de comunidad (Loidi et al 1995, Matthei 1995). Plantago major, Cyperus rotundus y Solidago chilensis señalan con su presencia que se trata de suelos compactados, con poca aireación (Méndez 1983, 1990). Es casi imposible mantener un cultivo en situaciones como las descriptas por lo que se produce la perdida del mismo.
Igual resultado se produjo en los campos invadidos por malezas de la asociación Cardario drabae - Agropyretum repentis formada por especies herbáceas, de ciclo perenne, hemicriptófitas y criptófitas. Glycyrrhiza astragalina, Tessaria absinthioides y Distichlis spicata denotan el carácter salino de los suelos (Conticello et al. 1997)
En la tabla de primavera - verano (Tabla 3) se mantienen seis de las siete asociaciones descriptas para otoño invierno, a la cual se agrega la comunidad de Centaurea melitensis.
Los indicadores del buen estado del cultivo siguen siendo las asociaciones dominadas por terófitos tales como Sysimbrio - Rapistretum y la comunidad de Stellaria media, que durante el periodo estival incluye a Urtica urens.
Setario - Echinochloetum coloni y Setario -Echinochloetum cruris gallii en este periodo presentan la mayor cobertura dado que adquieren el máximo desarrollo vegetativo por incluir a especies de ciclo estival. En los cultivos donde aparecen ambas asociaciones muestran un estado intermedio, por lo que deberían realizarse labores culturales accesorias.
Cynodetum dactyli y Cardario drabae- Agropyretum repentis fueron ya descriptas para el periodo anterior. La nueva comunidad de Centaurea melitensis que se diferencia en este ciclo, lo hace en cultivos abandonados y suelos poco productivos. Esta situación está corroborada por sus especies características, tales como Centaurea melitensis, C. calcitrapa y Xanthium spinosum que crecen en suelos difíciles de trabajar (Conticello et al. en prensa). El desarrollo de Distichlis spicata marca un probable incremento de la salinidad y compactación del suelo, lo que demandaría de mayores esfuerzos para que dichos campos puedan recuperarse.
Análisis de conglomerados
Los dendrogramas (Fig. 3 y 4) para los periodos otoño - invierno y primavera - verano complementan y permiten establecer una correspondencia con los grupos florísticos de las Tablas 2 y 3.
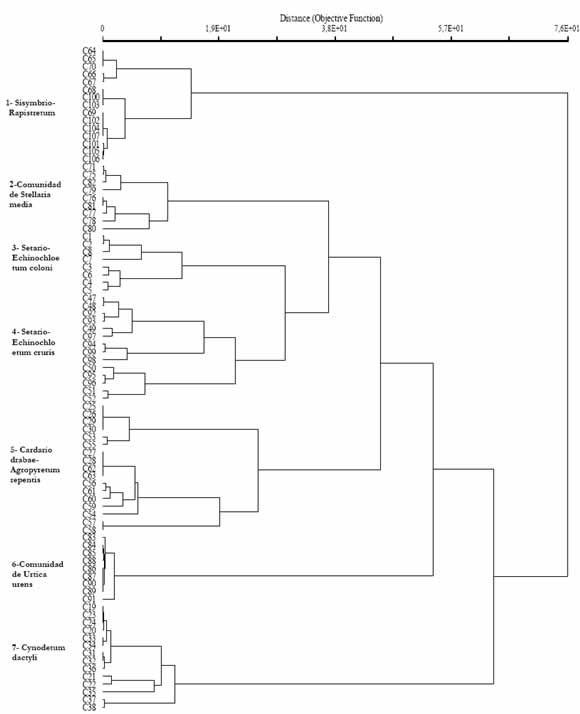
Fig. 3. Dendrograma de similitud florística de las comunidades de malezas en cultivos hortícolas, para el periodo otoño invierno (86 censos), mediante el uso de la distancia relativa euclídea.
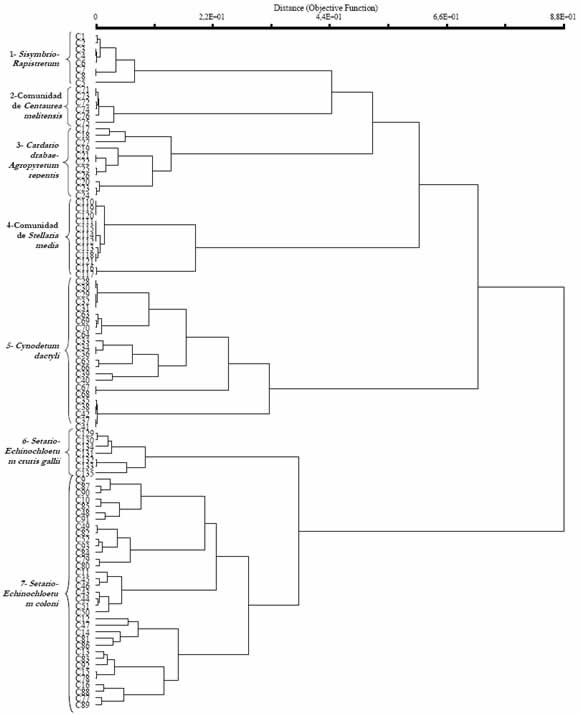
Fig. 4. Dendrograma de similitud florística de las comunidades de malezas en cultivos hortícolas, para el periodo primaveraverano (102 censos), mediante el uso de la distancia relativa euclídea.
Ambos dendrogramas muestran la presencia de ocho grupos, de los cuales seis se repiten en los dos periodos del año: Sisymbrio - Rapistretum, Cardario drabae - Agropyretum repentis, Cynodetum dactyli, comunidad de Stellaria media, Setario -Echinochloetum coloni y Setario - Echinochloetum cruris gallii. La diferencia entre los agrupamientos debida a riegos excesivos. Cardario drabae está dada por la estacionalidad de las especies que Agropyretum repentis que corresponde al grupo 5 los integran, las invernales alcanzan su máximo del dendrograma, crece en suelos desde arenosos a desarrollo vegetativo en esta estación del año y lo pedregosos, húmedos y salinos. La comunidad de mismo sucede con las del periodo estival. Urtica urens forma el grupo 6 y representa a lugares
En la Fig. 3 el primero y segundo grupos con suelos ricamente abonados, permeables y diferenciados corresponden al Sisymbrio -húmedos. El grupo 7, Cynodetum dactyli, caracteriza Rapistretum y la comunidad de Stellaria media. a suelos pesados, compactados, muy difíciles de Ambos grupos se desarrollan sobre suelos trabajar, con problemas de drenaje y salinidad. profundos, bien abonados y permeables. Los grupos La Fig.4 muestra siete grupos, de los cuales seis 3 y 4 corresponden a las asociaciones Setario -ya fueron descriptos para el periodo anterior. La Echinochloetum coloni y Setario -Echinochloetum comunidad de Centaurea melitensis que se diferencia cruris gallii que indican suelos compactados, pobres solo en el periodo primavero -estival, crece en suelos en nutrientes y con una mayor disponibilidad de agua, húmedos, salitrosos y pobres en materia orgánica.
Conclusiones
Los resultados obtenidos permiten concluir:
Se determinaron ocho asociaciones, de las cuales seis están comprendidas en la clase Stellarietea mediae (Br. Bl. 1931) Tx., Prsg. et Lohm, 1950 y las dos restantes en Artemisietea vulgaris Lohm, Prsg et Tüxen 1950.
Las asociaciones de la clase Stellarietea mediae (Br. Bl. 31) Tx. Prsg. et Lohm 1950 representan los diferentes estados de los cultivos y sus dinamismos. Se inicia con aquellos que presentan un excelente estado en cuyo caso abundan y dominan las especies anuales, únicas malezas que logran adaptarse a sitios altamente disturbados por labores culturales casi continuas como las que requieren las hortalizas.
Cuando las prácticas culturales son deficientes e incompletas favorecen el desarrollo de especies perennes y de difícil control, incluidas en su mayoría en la clase Artemisietea vulgaris Lohm., Prsg y Tx ex V. Rochow 1951.
Bibliografía
1. AHUMADA, O. & M, ALVAREZ. 2001. Malezas de los cultivos hortícolas de la Quebrada de Humahuaca. Pcia. de Jujuy. Argentina. Primera parte. Actas XXIV Congreso Argentino Horticultura. [ Links ]
2. ANDERBERG, M. 1973. Cluster analysis for applications. Academic Press. New York. [ Links ]
3. ANDREASEN, C. & J.C. STRIEBIG. 1991. Soil properties affecting the distribution of 37 weed species in Danish fields. Weed Res. Vol. 31: 181-187. [ Links ]
4. ATLAS DE LA PROVINCIA DE NEUQUEN. 1982. Provincia de Neuquén - U. N. del Comahue, Depto. de Geografía. [ Links ]
5. BARKMAN, J., J. MORAVEC, & S. RAUSCHERT. 1986. Code of phytosociological nomenclature. Vegetatio 67: 145-195. [ Links ]
6. BRAUN BLANQUET, J. 1979. Fitosociología. Bases para el estudio de las comunidades vegetales. Ediciones H. Blume. Madrid. [ Links ]
7. CENSO FRUTIHORTÍCOLA. 1994. Provincia del Neuquén. [ Links ]
8. CENSO NACIONAL AGROPECUARIO. 2002. INDEC. [ Links ]
9. CERAZO, M.B., A.P. BUSTAMANTE; R. GANDULLO, L. CONTICELLO & F. ROIG. 2000. Malezas en cultivos hortícolas de los departamentos Confluencia y Picún Leufú, provincia de Neuquén. Actas XXIII Congreso Argentino Horticultura. [ Links ]
10. CERAZO, M.B. & L. CONTICELLO. 2006. Sintaxomía de malezas presentes en cultivos de cebolla. Revista Horticultura Argentina. 25(29): 10-19. [ Links ]
11. COLBACH, N., F. DESSAINT & F. FORCELLA. 2000. Evaluating field-scale sampling methods for the estimation of mean plant densities of weeds. Weed Res. 40: 411-430. [ Links ]
12. CONTICELLO, L. & R. GANDULLO. 1991. Relevamiento de malezas estivales del Alto Valle de Río Negro y Neuquén. - Revista Malezas y su control. ASAM. 1: 19-26. [ Links ]
13. CONTICELLO, L.; A. BUSTAMANTE & C. TARTAGLIA. 1997. El Distichlo-Salicornietum ass. nov. en banquinas del Alto Valle de Río Negro y Neuquén. Actas del XIII Congreso Latinoamericano de Malezas. [ Links ]
14. CONTICELLO, L., R. GANDULLO, A. P. BUSTAMANTE & C. TARTAGLIA. 1998. Estudio fitosociológico de malezas ruderales en rutas nacionales del Alto Valle de Río Negro y Neuquén. Parte I. Parodiana 11: 75-98. [ Links ]
15. CONTICELLO; L., A.P. BUSTAMANTE & M.B. CERAZO. 2001. Ordenamiento sintaxonómico de las comunidades vegetales asociadas a cultivos hortícolas. Actas XXIV Congreso Argentino Horticultura. [ Links ]
16. CONTICELLO, L., M.B. CERAZO & A.P. BUSTAMANTE. 2002. Dinámica de comunidades hidrófilas asociadas a canales de riego en el Alto Valle de Río Negro (Argentina). Gayana Bot. 59: 13-20. [ Links ]
17. CONTICELLO, L., A. P. BUSTAMANTE, A & M. B. CERAZO. En prensa. «Sintaxones ruderales y adventicios en la zona del Alto Valle de Río Negro y Neuquén.» Parodiana 13. [ Links ]
18. COUSENS, R. D. & A.M. MORTIMER 1995. Weed population dynamics. Cambridge University Press. [ Links ]
19. GARDENER M. R., A. TYE & S.R. WILKINSON. 1999. Control of introduced plants on the Galapagos Islands. En: 12th Australian Weeds Conference Papers and Proceedings. Hobart, Tasmania. [ Links ]
20. GÉHU, J. M., J. GÉHU - FRANCK & A. SCOPPOLA. 1983. Schema synsystematique des vegetation nitrophiles et subnitrophiles de la region Nord/pas de Calais. Colloques phytosociologiques. Les vegetations nitrophiles et anthropogenes 12: 567-575. Ed. J. Cramer, Berlin. [ Links ]
21. GÉHU, J.M. & S. RIVAS MARTÍNEZ. 1982. Notions fondamentales de Phytosociologie. Ber. Internat. Symp. Syntaxonomie: 1 -33. Ed. J. Cramer, Vaduz. [ Links ]
22. HAUENSTEIN E., M. GONZALEZ, F. PEÑA-CORTES & A. MUÑOZ PEDREROS. 2002. Clasificación y caracterización de la flora y vegetación de los humedales de la costa de Tolten (IX Región, Chile). Gayana Bot. 59: 87-100. [ Links ]
23. LABRADA, R.. 1996. Gramíneas y Ciperáceas. En: Manejo de malezas para países en desarrollo. Addendum I. Estudio FAO Producción y Protección vegetal 120: 41-99 [ Links ]
24. LOIDI, J., A. BERASTEGUI, I. BIURRUN, I. GARCIA MIJANGOS & M. HERRERA. 1995. Data on Artemisietea vulgaris in the Basque country. Botanica Helvetica 105/2: 165-185 [ Links ]
25. MATTHEI J., O. 1995. Manual de las malezas que crecen en Chile. Alfabeta Impresores. Santiago de Chile. [ Links ]
26. MC CUNE B. & M.J. MEFFORD 1999. «PC- CORD» Multivariate analysis of ecological data, version 4. MJM Software design. Oregon, USA. [ Links ]
27. MEDINA, O., A.I. SANCHEZ & C.A. GONZÁLEZ VERA. 2004. Relevamiento de malezas en el cultivo de pimiento para pimentón (Capsicum annuum L.) en las localidades del Puesto, Las Mojarras y San José Banda. Dpto. Sta. María, Catamarca. Actas XXVII Congreso Argentino de Horticultura. [ Links ].
28. MÉNDEZ, E. 1983. Observaciones sobre la flora adventicia de viñedos en Mendoza. Parodiana 2: 263- 276. [ Links ]
29. MÉNDEZ, E. 1984. Observaciones ecológicas sobre la vegetación adventicia de cauces de riego en Mendoza. Parodiana 3: 185-196. [ Links ]
30. MÉNDEZ, E. 1990. Observaciones fitosociológicas de la vegetación adventicia de cultivos hortícolas en la provincia de Mendoza. Parodiana 6: 197-209. [ Links ]
31. MÉNDEZ, E. 2002. Una comunidad de malezas en cultivos de Mendoza, Argentina: Brassicetum rapae ass. nov. Bol. Soc. Argent. Bot. 37: 93-97. [ Links ]
32. MORTIMER, A.M. 1996. La clasificación y ecología de las malezas. En: Manejo de malezas para países en desarrollo. Addendum I. Estudio FAO Producción y Protección vegetal 120: 13-30. [ Links ]
33. NOBILE, R. A., M. A. DELFINO & M. J. LANTIERI. 1989. El complejo áfidos - malezas en cultivos hortícolas del cinturón verde de la ciudad de Córdoba. Revista de Malezas ASAM. 17: 29 - 40. [ Links ]
34. ORLOCI, L. 1978. Multivariate analysis in vegetation research. 2° edición. Dr. W. Junk, The Hague. [ Links ]
35. PANETTA, F.D. & N.D. MITCHELL.1991. Homoclime analysis and the prediction of weediness. Weed Res. 31:273-284. [ Links ]
36. PAPADAKIS, J. 1980. El clima. Ed. Albatros. Buenos Aires. [ Links ]
37. PAUCHARD, A. & P. ALABACK. 2004. Influence of elevation, land use and landscape context on patterns of alien plant invasions along roadsides in protected areas of south-central Chile. Conservation Biol. 18: 238-248. [ Links ]
38. PYSEK, P. & A. PYSEK, 1991. Succession in urban habitats: an analysis of phytosociologycal data. Urban ecology: 105 -112. Edited by H. Sukopp et al. [ Links ]
39. RAUNKIAER, C. 1934. Life forms and terrestrial plant geography. Clarendon Press. Oxford. [ Links ]
40. RIVAS - MARTÍNEZ, S. 1978. Sobre la vegetación nitrófila del Chenopodion muralis. Acta Botanica Malacitana 4:71-78. [ Links ]
41. SERVICIO METEOROLÓGICO NACIONAL. 1992. Estadísticas climatológicas 1980-1990. Serie B- N° 37: 709. Primera edición. Bs. As. [ Links ]
42. SHALTOUT, K.H. & R.A. EL FAHAR. 1991. Diversity and phenology of weed communities in the Nile Delta region. J Veg Sci 2: 385-390. [ Links ]
43. SUAREZ, S., E. DE LA FUENTE, C. GHERSA & R. LEÓN.1997. Comunidades de malezas en cultivos estivales: relaciones con el ambiente productivo (Pampa ondulada-Argentina). Actas XIII Congreso. Latinoamericano Malezas. Tomo I: 10-16. [ Links ]
44. THOMAS, A.G. & M. R. T. DALE. 1991. Weed community structure in spring - seeded crops in Manitoba. Can. J. Plant. Sci. 71: 1069 - 1080. [ Links ]
45. THOMAS, A.G., B. FRICK, D.A. DERKSEN, S.A. BRANDT & R.P. ZENTNER. 1996. Crop rotations and weed community dynamics on the Canadian prairies. p. 227- 232. In H. Brown et al. (ed.) Proc. Int. Weed Control Congr. 2nd, Copenhagen, Denmark [ Links ]
46. THORNTHWAITE, C. W. 1948. An approach toward a rational classification of climate. Geograp rev. 55 - 94. [ Links ]
47. WARD, J. H. 1963. Hierarchical grouping to optimize an objective function. Amer. Statist. Ass. J. 58: 236-244. [ Links ]
48. WEBER, H.E., J. MORAVEC & J.P. THEURILLAT. 2000. International Code of Phytosociological Nomenclature. Third edition. J Veg. Sci. 11: 739-768. [ Links ]
Recibido el 29 de Agosto de 2007, a
Aceptado el 28 de Marzo de 2008.

