










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. v.44 n.1-2 Córdoba ene./jul. 2009
Anatomía Vascular de Plántulas de Cicer Arietinum. Genotipo M x WR-315-14 (Fabaceae)
N. S. Ateca1, A. Pascualidades1 y J. Carreras2
1 Cátedra de Botánica Morfológica. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Ciudad Universitaria. 5000 Córdoba. Argentina. E-mail: norateca@agro.unc.edu.ar
2 Cátedra de Mejoramiento Vegetal. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Ciudad Universitaria. 5000 Córdoba. Argentina
Resumen: Se estudiaron los caracteres anatómicos de la vascularización de plántulas de Cicer arietinum L. (Fabaceae, Faboideae, Cicereae) pertenecientes al genotipo M x WR-315-14. La anatomía vascular de la raíz, hipocótilo, nudo cotiledonar, epicótilo, segundo y tercer entrenudo de plántulas normales con el protofilo desplegado, fueron analizados para determinar la localización y detalles sobre las variaciones en el desarrollo de los elementos conductores. Estos datos serán de interés para la identificación en estados juveniles de genotipos de la especie.
Palabras clave: Anatomía; Vascularización; Plántulas; Genotipo; Cicer arietinum.
Summary: Vascular anatomy of seedlings of Cicer aretinum L. M x WR-315-14 genotype (Fabaceae). We studied the vascular anatomy in seedlings of Cicer aretinum L. M x WR-315-14 genotype (Fabaceae, Faboideae, Cicereae), with the aim of determining the localization and development of conducting elements. We analyzed the vascular anatomy of roots, hypocotyls, cotyledonal nudes, epicotyls, second and third internodes of standard seedlings with unfolded protophylls. These data will contribute to the identification of different genotypes of the species in juvenile stages.
Key words: Anatomy; Vascular; Seedlings; Genotype; Cicer arietinum.
INTRODUCCIÓN
La sobrevivencia de una plántula depende del establecimiento de un sistema de transporte continuo dentro del eje de la planta, cualquier interferencia en la diferenciación del sistema, tanto externa como interna, produce una alteración en la conexión vascular que se refleja en el desarrollo de la futura planta.
En plántulas de dicotiledóneas con germinación hipogea la región de transición presenta una longitud máxima, ya que comprende uno o más nudos sobre los cotiledones (Fahn, 1974). La extensión de la región de transición en uno o más entrenudos por encima de los cotiledones, puede explicarse por la influencia del ápice radicular en los entrenudos basales del tallo, como resultado del crecimiento retardado del hipocótilo en tales semillas (Esau, 1976). Por el contrario, Muller (1937) menciona que en Cicer arietinum L., especie de germinación hipogea, la estructura de la porción central del eje de los primeros cuatro internodios no presenta relación con la región de transición de la raíz al tallo y atribuye la organización característica de los internodios inferiores a la orientación de los haces vasculares que recorren las primeras hojas bracteiformes o, a la manera como ellos se reúnen.
Estudios realizados en Pisum sativum L., determinan que la región de transición no se completa en el corto hipocótilo, sino que abarca los primeros tres internodios del tallo, siendo una dictiostela en el cuarto internodio (Hayward, 1953).
En plántulas normales de Phaseolus vulgaris L., Harris & Sinnot (1921) determinaron que la raíz es tetrarca, presenta en la base del hipocótilo cuatro haces vasculares dobles que se dividen en ocho haces en la porción apical del mismo y de los cuales dos parten hacia cada cotiledón. El tejido vascular remanente se reorganiza en seis cordones que continúan dividiéndose en dos, presentándose en el epicótilo doce haces vasculares. Los autores establecen dos tipos diferentes de haces vasculares, primarios dobles e intercalares o adicionales.
Por otra parte, en Arachis hipogaea L., el cilindro vascular de la raíz primaria también es tetrarca y presenta médula. La transición de xilema exarco a endarco se produce en la zona basal del hipocótilo donde se presentan ocho haces vasculares sobre el cual se produce fusión y ramificación con un patrón irregular. Dos grandes haces vasculares del hipocótilo se emiten hacia los cotiledones que pueden estar acompañados de un estrato de protoxilema. La raíz y el hipocótilo desarrollan capas de cámbium vascular. La zona de transición se encuentra rodeada de anillos de células con taninos (Yarbrough, 1949).
Esau (1974), refiere que la característica de la zona de transición es enteramente primaria. Cuando se presenta actividad cambial, los tejidos secundarios se forman en completa continuidad entre el tallo y la raíz. El crecimiento secundario oscurece las diferencias iniciales en la estructura de la raíz, hipocótilo y epicótilo, dejando sólo el xilema en la región de transición relegado en el centro del eje.
Por otra parte, Cubero (1987) cita que C. arietinum no presentaría hipocótilo, la transición vascular entre la raíz y el tallo se concreta en la región basal de los cotiledones y se caracteriza por la ausencia de estructuras secundarias.
El objetivo del presente trabajo es analizar la anatomía vascular de plántulas normales de Cicer arietinum (Fabaceae, Faboideae, Cicereae), pertenecientes al genotipo M x WR-315-14. Los datos obtenidos contribuirán al conocimiento de la especie y a la correcta diferenciación entre genotipos en estados juveniles.
MATERIAL Y MÉTODOS
Las semillas del genotipo M x WR-315-14 de C. arietinum provienen de los ensayos realizados en el Proyecto "Contribución al mejoramiento genético del garbanzo (Cicer arietinum L.). Adaptación de líneas selectas", dirigido por la Ing. Agr. J. Carrera y cosechadas en los años 2005 y 2006, en: ARGENTINA. Prov. Córdoba: Dpto. Cruz del Eje, San Marcos Sierra. Las semillas fueron embebidas durante 24 horas y sembradas en bandejas plásticas con sustrato papel a temperatura ambiente ± 20° C. En el año 2006 se analizaron 20 plántulas en diferentes estados de desarrollo, determinando la estructura de plántula normal como aquella compuesta por el primer protofilo con estípulas, expandido y que presente un equilibrio entre el desarrollo de la región subterránea y aérea. En el año 2007 se estudiaron 30 plántulas normales según los criterios definidos previamente. Se realizaron preparados temporarios de cortes seriados transversales por raíz, hipocótilo, nudo cotiledonar, epicótilo, segundo y tercer entrenudo de plántulas. Se tiñeron con Safranina y Azul Astra (Luque et al., 1996). Las fotomicrografías se obtuvieron con microscopio binocular Nikon Fase contraste 2, Altha Thot2 YS2 Cámara de Video color SONY y Programa Aver TX GO 007 Plus. Slow View. Las fotografías de plántulas se obtuvieron con microscopio estereoscópico y cámara digital de 6 megapixeles y precisión 12 X.
RESULTADOS
Plántula
Las plántulas presentan germinación criptohipógea con catafilos alternos dísticos en el segundo y tercer nudo. El protofilo estipulado, imparipinnaticompuesto emerge del cuarto nudo, presentando menor número de foliolos que los nomofilos. La raíz principal presenta raíces laterales y el hipocótilo glabro se ensancha hacia la unión de los pecíolos cotiledonares (Fig. 1 A).
Región subterránea
Raíz. La epidermis es uniestratificada, la corteza comprende siete a nueve capas de células parenquimáticas grandes y redondeadas con marcados espacios intercelulares. Presenta algunos granos de almidón. La exodermis y endodermis exhiben Bandas de Caspary. El periciclo con células de paredes delgadas, es uniseriado en la cara externa del protofloema, y presenta dos a tres capas de células en la zona de los polos del protoxilema. La diferenciación del xilema primario es centrípeta. La raíz primaria es tetrarca. En ese estado, se observa el cámbium vascular con desarrollo continuo entre los cordones de los tejidos vasculares y periciclo (Fig. 1 B, a y b; Fig. 2 A).
Hipocótilo. Se inicia con la diferenciación de elmentos parenquimáticaticos en la zona central del cilindro (Fig. 1 B c y Fig. 2 B). Los nuevos elementos conductores del xilema se originan por actividad del cámbium vascular, mientras que el tejido floemático está constituido por elementos conductores, fibras remanentes del floema primario y el aporte de elementos de origen cambial. En la región media del hipocótilo, el tejido xilemático se organiza en cuatro cordones (en forma de Y) constituidos principalmente por miembros de vaso de origen cambial y procambial (Fig. 1 B d y Fig. 2 C). En la región apical el hipocótilo adopta un contorno circular por incremento de la zona medular; acompañan este incremento divisiones anticlinales y periclinales del periciclo y endodermis, diferenciándose dos pares de haces vasculares constituidos por xilema y floema de origen cambial y procambial, denominados secundarios, y dos pares compuestos por xilema de origen cambial y floema procambial y cambial, denominados intercalares. En esta región las células corticales internas presentan pigmentos (Fig. 1 B e y Fig. 2 D).
Nudo cotiledonar. Se observan en corte transversal, seis hacecillos colaterales enfrentados, cuatro de tipo secundario, que alternan con dos de tipo intercalar; además hay dos hacecillos opuestos de tipo intercalar más pequeños. Estos últimos, se fragmentan constituyendo cada uno dos trazas. Una de ellas ingresa al pecíolo de los cotiledones, ramificándose en dos en la región media del mismo y posteriormente en cuatro en la región de las aurículas. La segunda traza se dirige a la yema ubicada en la axila del pecíolo cotiledonar (Fig. 1 B f).
b- Región aérea
Epicótilo (Primer entrenudo). En la zona media se observan seis hacecillos vasculares colaterales con cámbium fascicular, cuatro del tipo secundario enfrentados y dos de naturaleza intercalar opuestos, todos relacionados por cámbium interfascicular (Fig. 1 B g y Fig. 3 A). Próximo al segundo nudo del cual nace el catafilo basal, los hacecillos secundarios relacionados a él se dividen anticlinalmente por actividad de células iniciales radiales del cámbium, resultando dos hacecillos de diferente tamaño que determinan la presencia de ocho hacecillos vasculares en la región apical del primer entrenudo (Fig. 1 B h; Fig. 3 B). La epidermis del epicótilo es uniestratificada y presenta abundante pelos pluricelulares glandulares. La corteza externa presenta una capa de células de colénquima laminar y una capa interna, la endodermis.
Segundo entrenudo. Los hacecillos ubicados hacia el nacimiento del segundo catafilo en el tercer nudo, experimentan divisiones radiales, al igual que el primer entrenudo, quedando conformados diez hacecillos vasculares conectados por cámbium interfascicular (Fig. 1, B i).
Tercer entrenudo. En el entrenudo se observan diez hacecicllos vasculares y actividad de cámbium fascicular e interfascicular (Fig. 1 B j; Fig. 3 C y D). El tercer entrenudo está recorrido por una costilla, compuesta por un haz fibroso y colénquima angular, que finaliza en la inserción de las estípulas del protofito imparipinnado.

Fig. 1. Plántula de C. arietinum cv M x WR-315-14. A: Fotografía de la plántula y posición de los cortes transversales representados gráficamente. B: Esquemas con símbolos de los cortes transversales de raíz (a y b), hipocótilo (c, d y e), nudo cotiledonar (f), región media del epicótilo (g), región apical del epicótilo (h), región apical del segundo entrenudo (i) y región media del tercer entrenudo (j). Abreviaturas: ca: cámbium; xs: xilema secundario; fs: floema secundario; hs: haces secundarios; hi: haces intercalares; c: catáfilo.

Fig. 2. Anatomía de raíz e hipocótilo de C. arietinum cv M x WR-315-14, A-D: Fotomicrografías de cortes transversales. A: Desarrollo centrípeto de la raíz y disposición continua del cámbium. B: Región basal de hipocótilo con pocas células parenquimáticas. C: Región media del hipocótilo con región medular y desarrollo de elementos conductores secundarios. D: Región apical del hipocótilo y comienzo de organización de hacecillos secundarios e intercalares Abreviaturas: ca: cámbium; p: parénquima; xs: xilema secundario; fs: floema secundario; hs: haces secundarios; hi: haces intercalares. Barras: A= 300 µm; B, C y en D=500 µm.
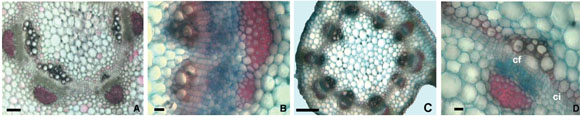
Fig. 3. Anatomía del primer entrenudo (epicótilo) y tercer entrenudo C. arietinum cv M x WR-315-14. A-D: Fotomicrografías de cortes transversales. A: Hacecillos secundarios (cuatro) y hacecillos intercalares (dos más pequeños) en la región media del epicótilo. B: Hacecillo secundario del epicótilo fragmentándose por actividad de células cambiales radiales. C: Tercer entrenudo con diez hacecillos. D: Cámbium fascicular e interfascicular del tercer entrenudo. Abreviaturas: cf: cámbium fascicular; ci: cámbium interfascicular. Barras: A y C= 200 µm, B y D= 500 µm.
DISCUSIÓN Y CONCLUSIONES
La raíz principal de las plántulas de garbanzo pertenecientes al genotipo M x WR-315-14 es tetrarca y contrariamente a lo sostenido por Cubero (1987) se distingue morfológicamente el hipocótilo. El periciclo enfrentado al protoxilema presenta dos a tres estratos de células. El floema primario presenta cuatro grupos de fibras correspondientes al protofloema y enfrentados al periciclo uniestratificado. Entre las regiones del floema y xilema se diferencian el cámbium vascular que es activo, produciendo elementos conductores secundarios. Este aspecto, no coincide con lo encontrado por Hayward (1953) en Pisum sativum donde el cámbium es inactivo o su actividad está restringida a una pequeña extensión lateral, y lo citado por Cubero (1987) sobre ausencia de elementos conductores de origen secundario. El hipocótilo se caracteriza por la presencia de células parenquimáticas en el lugar ocupado por el metaxilema y elementos conductores secundarios derivados del cámbium vascular.
La zona de transición del genotipo no es de naturaleza exclusivamente primaria como lo manifiesta Esau (1976), ya que se observa rotación de posición exarca a endarca del xilema de origen procambial en los cuatro hacecillos secundarios. En consecuencia, la transición vascular sólo comprendería a esos hacecillos. La presencia de hacecillos secundarios, sumados a los de tipo intercalar con xilema de origen cambial, establecen una conexión entre la región subterránea (raízhipocótilo) con la aérea de acuerdo a lo afirmado por Esau (1976).
Con respecto a la pigmentación de las células corticales observadas por Yarbrough, (1949) y que delimitan para el autor la zona de transición en Arachis hypogaea se correspondería con lo observado para la línea estudiada.
La ordenación vascular de la región aérea se relaciona con la disposición de los catafilos y el protofilo. Los entrenudos exhiben ramificaciones progresivas de hacecillos a partir del epicótilo y originadas por divisiones periclinales de células iniciales radiales del cámbium que cesan en el cuarto entrenudo.
Según las observaciones realizadas, se determina que la región de transición no se extiende hasta los primeros internodios como lo manifiestan Hayward (1953), Fahn (1974) y Esau (1976), y se coincide con lo expresado por Muller (1937) para C. arietinum, quien atribuye la organización de los internodios inferiores a la orientación de los haces vasculares que recorren las primeras hojas catafilares o a la manera como ellos se reúnen. Cabe destacar que el acuerdo expresado con la autora es válido para el análisis del estado de desarrollo de la plántula normal del genotipo definido en el presente estudio.
BIBLIOGRAFÍA
1. CUBERO, J. 1987. Morphology of chickpea. In: SAXENA M.C.& K.B. SINGH (eds), The Chickpea, pp. 35-66. CAB International, Wallingford, UK. [ Links ]
2. ESAU, K. 1976. Anatomía Vegetal. Ed. Omega. Barcelona. [ Links ]
3. FAHN, J. 1974. Anatomía Vegetal. H. Blume Ediciones. Madrid. [ Links ]
4. HARRIS J. & E. SINNOT. 1921. The vascular anatomy of normal and variant seedlings of Phaseolus vulgaris. Proc. Nat. Acad. Sci. U.S.A. 7 (1): 35-41 [ Links ]
5. HAYWARD, H. E. 1953. Estructura de las plantas útiles. Acme. Buenos Aires [ Links ]
6. LUQUE, R., H.C. SOUSA & J.E. KRAUS. 1996. Métodos de coloração de Roeser (1972) modificado e Kropp (1972) visando a substituição do Azul de Astra por Azul de Alcião 86S 86X. Acta Bot. Brasil. 10: 199-212. [ Links ]
7. MULLER, C. 1937. La tige feuillée et le cotylédons des Viciées a germination hypogée. Cellule 46: 195-354. [ Links ]
8. YARBROUGH, J. 1949. Vascular transition of Arachis hypogaea seedling. Amer. J. Bot. 36 (10): 758-772. [ Links ]
Recibido el 27 de octubre de 2008
Aceptado el 12 de mayo de 2009.

