










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
On-line version ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.44 no.3-4 Córdoba Aug./Dec. 2009
ANATOMÍA Y MORFOLOGÍA
Patrones de venación floral en tres especies de Paullinieae (Sapindaceae)
Stella M. Solís1 y María S. Ferrucci2
1 Facultad de Ciencias Exactas, Naturales y Agrimensura, Universidad Nacional del Nordeste, Instituto de Botánica del Nordeste (UNNE-CONICET), C.C. 209, 3400 - Corrientes, Argentina. Autor para correspondencia, E-mail: estelissolis@yahoo.com
2 Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste, Instituto de Botánica del Nordeste (UNNE- CONICET), C.C. 209, 3400-Corrientes, Argentina. E-mail: msferrucci@yahoo.com.ar
Resumen: Se analizaron los patrones de venación floral en Cardiospermum grandiflorum, Urvillea chacoensis y Thinouia mucronata (Sapindaceae-Paullinieae). Estas especies son monoicas y la inflorescencia es un tirso que posee flores estaminadas y bisexuadas funcionalmente pistiladas. Cardiospermum grandiflorum y U. chacoensis pertenecen a la subtribu Paulliniinae, caracterizada por las flores oblicuamente zigomorfas; mientras T. mucronata pertenece a Thinouiinae, subtribu monotípica con flores actinomorfas. En las especies estudiadas el patrón de venación es similar en ambos tipos de flores, a partir de una sifonostela ectofloica central irradian trazas vasculares que inervan los verticilos periánticos, presentando el fenómeno de progresiva fusión y adnación. El tejido vascular que inerva los nectarios está representado por cordones de floema. En relación a la vascularización floral de estas especies, las diferencias en la venación se observan en las trazas de los complejos sépalo-petalar que irradian de la sifonostela ectofloica central; mientras el patrón de venación de los verticilos fértiles es común a todas ellas. La información obtenida apoya la posición basal de Thinouia en la tribu Paullinieae.
Palabras clave: Sapindaceae; Paullinieae; Vascularización floral; Cardiospermum grandiflorum; Urvillea chacoensis; Thinouia mucronata.
Summary: Floral venation patterns in three species of Sapindaceae-Paullinieae. The floral venation patterns of Cardiospermum grandiflorum, Urvillea chacoensis and Thinouia mucronata, are surveyed. These species are monoecious and the inflorescence is a thyrse carrying staminate and bisexual flowers funcionally pistillate. Cardiospermum grandiflorum and U. chacoensis belong to the subtribe Paulliniinae which is characterized by obliquely zygomorphic flowers, whereas T. mucronata belongs to the monotypic Thinouiinae which presents actinomorphic flowers. Vascular pattern is similar in both types of flowers of the studied species; from one ectofloic siphonostele depart vascular traces that supply the perianth whorls displaying phenomena of progressive fusion and adnation. Vascular tissue supplying the nectaries is characterized by strands of phloem. Regarding the floral vasculature in these species, differences in the vasculature lay primarily in the number of the sepal-petal complexes traces departing from the central ectophloic siphonostele; whereas the venation pattern of the fertile verticils is common to them. The data obtained support the basal position of Thinouia in the tribe Paullinieae.
Key words: Sapindaceae; Paullinieae; Flower venation patterns; Cardiospermum grandiflorum; Urvillea chacoensis; Thinouia mucronata.
INTRODUCCIÓN
La familia Sapindaceae consta de 14 tribus, (Radlkofer, 1931-1934), de las cuales sólo Paullinieae posee especies con hábito trepador, las restantes son arbóreas o arbustivas. Esta tribu presenta dos subtribus Paulliniinae y Thinouiinae; las especies son monoicas con flores estaminadas y bisexuadas funcionalmente pistiladas.
Cardiospermum L. y Urvillea Kunth, pertenecen a Paulliniinae, subtribu que se caracteriza por presentar flores oblicuamente zigomorfas, con corola tetrámera, los pétalos con la escama en forma de capucha, generalmente insertos en una pequeña depresión de las glándulas nectaríferas. Los frutos pueden ser esquizocarpos o cápsulas (Radlkofer,1931-1934; Ferrucci, 2000a).
La subtribu Thinouiinae cuenta con un único género, Thinouia Triana & Planch., de posición basal en la tribu (Muller & Leenhouts, 1976; Ferrucci & Anzótegui, 1993; Ferrucci, 2000b). El carácter más significativo para diferenciarla de Paulliniinae es la simetría floral, Thinouia posee flores actinomorfas, con cáliz pentámero, gamosépalo, con los sépalos soldados en 1/3 basal, de apertura precoz y la corola es pentámera, dialipétala; otros caracteres que distinguen este género son el tipo de polen y la morfología del fruto.
La vascularización floral ha constituido siempre un tema relevante de investigación en taxonomía y filogenia. La venación floral no sólo ha sido utilizada para resolver problemas filogenéticos sino también para establecer homologías de los órganos florales (Gustafson, 1995). Endress et al. (2000) sostienen que los modelos de vascularización no gobiernan la morfología, pero dependen principalmente del modo de desarrollo y de la función de los órganos.
Los estudios de vascularización floral en especies de Sapindaceae son muy escasos. Nair & Joseph (1960) describen la anatomía vascular de las flores estaminadas y pistiladas de Cardiospermum halicacabum L. Recientemente, Weckerle & Rutishauser (2005) analizan la estructura del gineceo en antesis, del fruto y de la semilla en especies de Paullinia (5 spp.), en Cardiospermum halicacabum L., en Serjania altissima (Poepp.) Radlk. y en Urvillea ulmacea Kunth.
El objetivo de este trabajo es analizar la vascularización floral de Cardiospermum grandiflorum, Urvillea chacoensis y Thinouia mucronata, estudio que contribuirá a la interpretación de los caracteres florales en la tribu y aportará información para consideraciones filogenéticas en la familia.
MATERIALES Y MÉTODOS
Se utilizaron botones florales preantesis de flores estaminadas y flores pistiladas de las tres especies, los mismos fueron conservados en formol acetoalcohólico (FAA). El material se deshidrató en una serie de deshidratantes y de alcohol butíricoterciario (González & Cristóbal, 1997) y se incluyó en parafina (Johansen, 1940). Los cortes seriados transversales se realizaron con micrótomo rotativo en espesores de 10-15µm, los mismos se tiñeron con safranina-azul de Astra (Luque et al., 1996) y fueron montados en bálsamo de Canadá. Para el estudio de la vascularización se realizaron cortes transversales sucesivos desde el pedicelo hasta el estigma.
Las observaciones, fotografías y dibujos se efectuaron con un microscopio estereoscópico Leica MZ6 y un microscopio óptico Leica DM LB2 ambos equipos con cámara clara y cámara digital.
Los ejemplares testigo se encuentran depositados en el herbario del Instituto de Botánica del Nordeste (CTES).
C. grandiflorum Sw. ARGENTINA. Prov. Corrientes: Dpto. Berón de Astrada, 10-I-2007, Ferrucci et al. 2711; Dpto. Capital, 25-VIII-1978, Ferrucci 32; Dpto. San Cosme, 30-I-2006, Ferrucci 2217; Dpto. San Cosme, 06-II-2008, Ferrucci et al. 2829. Prov. Formosa: Dpto. Formosa, 1-II-2007, Ferrucci et al. 2745. BOLIVIA. Dpto. Santa Cruz, 4IV-2006, Ferrucci et al. 2524.
U. chacoensis Hunz. ARGENTINA. Prov. Salta: Dpto. San Martín, 8-IV-2004, Meza Torres et al. 214. BOLIVIA. Dpto. Santa Cruz, 19-VII-2003, Ferrucci et al. 1785; Dpto. Santa Cruz, 29-III-2006, Ferrucci et al. 2258; Dpto. Santa Cruz, 12-IV-2006, Ferrucci et al. 2680; Dpto. Santa Cruz, 14-IV-2006, Ferrucci et al. 2701.
T. mucronata Radlk. ARGENTINA. Prov. Misiones: Dpto. Eldorado, 4-II-2008, Keller 5018; Dpto. Guaraní, 6-I-2007, Ferrucci et al. 2726; Dpto. SanIgnacio, 12-II-2008, Ferrucci et al. 2832.
RESULTADOS
Cardiospermum grandiflorum Sw. (Fig. 1 A, B)
Morfología floral: Presenta flores reunidas en tirsos racemiformes, blanquecinas, de (5,5) 6-9,25 (14) mm long. El cáliz es tetrámero con 2 sépalos externos breves, suborbiculares y 2 internos obovados, petaloideos. La corola dialipétala, posee 4 pétalos, 2 posteriores con la escama basal simétrica, el apéndice deflexo de la escama es comparativamente largo y alcanza la base del pétalo y 2 anteriores con la escama asimétrica. Dos nectarios corniculiformes se ubican en la base del androginóforo, son blanquecinos, de 1,25-3,5 mm dealtura, protegidos por el apéndice deflexo de la escama de los pétalos posteriores. El androginóforo es pubérulo, en la parte superior se insertan 8 (9) estambres conniventes en la base, de diferenteslongitudes, 3 altos, 2 laterales medianos y 3 bajos. En las flores estaminadas, el gineceo está reducido aun breve pistilodio. En las flores pistiladas sediferencian 8 (9) estambres de menor longitud conanteras indehiscentes, el gineceo es excéntrico, 3carpelar, un óvulo campilótropo por carpelo, conovario trígono-obovoide o elipsoide, velloso y estilofiliforme más largo que las ramas del estigma.
Vascularización floral: Dos haces colaterales fundamentales se fusionan a la altura del receptáculo, constituyendo una sifonostela central ectofloica (Fig. 2 A). A partir de la misma se desprenden 6 trazas, 3 van a inervar al sépalo externo derecho y 3 al sépalo externo izquierdo (Fig.2 B).

Fig. 1. Cardiospermum grandiflorum. A: Flor estaminada. B: Flor bisexuada funcionalmente pistilada. Urvillea chacoensis. C: Flor estaminada. D: Flor bisexuada funcionalmente pistilada. Thinouia mucronata. E: Flor estaminada. F: Flor bisexuada funcionalmente pistilada. Escala = 2 mm. A: Ferrucci et al. 2745 (CTES); B: Ferrucci 2217 (CTES); C: Ferrucci et al. 2680 (CTES); D: Meza Torres et al. 214 (CTES); F-G: Keller 5018 (CTES); G, material conservado en FAA).
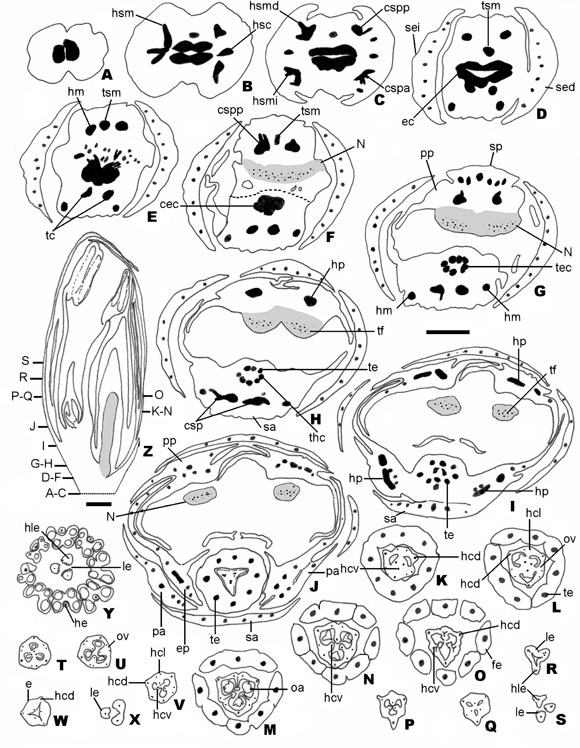
Fig. 2. Vascularización floral de C. grandiflorum. A-S: Cortes transversales sucesivos de una flor pistilada. T-Y: Cortes transversales sucesivos de una flor estaminada. Z: Corte longitudinal de botón floral de flor estaminada en preantesismostrando la posición de los transcortes. Escalas: A-Z = 500 µm. Abreviaturas: sei: sépalo externo izquierdo; sed: sépaloexterno derecho; hsm: haz sepalar marginal; hsc: haz sepalar central; hsmi: haz sepalar marginal izquierdo; hsmd: hazsepalar marginal derecho; csp: complejo sepalar-petalar; N: nectario; ec: estela central; cspp: complejo sepalar-petalarposterior; sp: sépalo posterior; cspa: complejo sepalar-petalar anterior; tsm: traza sepalar media; hm: haz marginal; pp:pétalo posterior; ep: escama petalar; n: tejido nectarífero; tc: traza central; sa: sépalo anterior; hp: haz petalar; tf: trazasfloemáticas; he: haz estaminal; hcl: haz carpelar lateral; hcd: haz carpelar dorsal; hcv: haz sepalar ventral; ov: ovario; o:óvulo; oa: óvulo atrofiado; le: lóbulo estigmático; e: estilo; fe: filamento estaminal; hle: haz lóbulo estigmático. A-S:Ferrucci et al. 2745 (CTES); T-Z: Ferrucci 2217 (CTES).
Cada sépalo posee 2 haces sepalares marginales y un haz central, inmediatamente los 2 sepalares marginales que llamaremos derecho e izquierdo, se bifurcan, el haz sepalar marginal derecho va aconstituir el complejo sépalo-petalar del cual una ramificación inervará el sépalo externo, el sépalo posterior y el pétalo posterior derecho. Lo mismo ocurre con el haz sepalar marginal izquierdo, formando el complejo sépalo-petalar anterior (Fig. 2C-E).
De la porción posterior de la sifonostela central se desprende una traza, denominada traza sepalar media, la misma junto con los haces marginales provenientes de los complejos sépalo-petalares posteriores del sépalo derecho y del sépalo izquierdo respectivamente, vascularizan al sépalo posterior y a los dos pétalos posteriores; los haces petalares se ramifican para inervar la escama (Fig. 2 F-G).
De la porción anterior de la sifonostela central emergen 2 trazas. Estas trazas juntas con los haces marginales provenientes de los complejos sépalo-petalares de los sépalos derecho e izquierdo inervanal sépalo anterior y a los 2 pétalos anteriores (Fig. 2F-H). Las trazas petalares de cada pétalo se ramifican para inervar las escamas respectivamente (Fig. 2 I-J).
El tejido vascular central se ramifica hacia laparte posterior con numerosas y reducidas trazas que van a irrigar el tejido nectarífero (Fig. 2 E-H). Se reconocen entre 6-12 trazas floemáticas que llegan hasta el ápice del nectario. Las trazas están representadas por miembros de tubos cribosos, angostos y con el citoplasma teñido, células acompañantes pequeñas y células parenquimáticasde contorno poligonal de paredes rectas y núcleo conspicuo.
Posteriormente el complejo estamino-carpelar se fragmenta en 8 (9) trazas de estambres y 3 trazas dorsales carpelares (Fig. 2 I). Cada una de las trazas estaminales termina ciega en el tejido conectivo; los hacecillos varían en la misma flor y al mismo nivel de colaterales a concéntricos anficribales (Fig. 2 JM). Las 3 trazas dorsales carpelares orientadas hacia el centro formando una estela triangular; cada uno de éstos se divide en dos haces de menor tamaño denominados haces ventrales o marginales. Los lóbulos del ovario aparecen a este nivel (Fig. 2 K, T).
En el centro del ovario se encuentran tres haces producto de la fusión de los haces ventrales del carpelo. A partir de estas trazas se desprenden los haces placentarios que irrigan los respectivos rudimentos seminales; los haces dorsales del ovario también se ramifican en hacecillos más pequeños denominados haces laterales, que recorren la pared de cada carpelo. Por encima de la región ovular, todos los hacecillos de menor calibre se fusionan a tres haces dorsales cada uno proveniente de uncarpelo. El estilo se encuentra inervado por los 3 haces dorsales y los 3 haces ventrales. En la parte superior del estilo los haces ventrales se fusionan a los tres haces dorsales que terminan inervando acada una de las tres ramas del estigma (Fig. 2 K-Y). El ovario es sinascidiado en aproximadamente la mitad de su longitud, luego simplicado, hemisimplicado próximo a la base de las tres ramas del estigma, en éste es asimplicado.
Urvillea chacoensis Hunz. (Fig. 1 C, D)
Morfología floral: Presenta flores dispuestas en tirsos subracemiformes, blanquecinas, de 4-7 mm long. El cáliz es pentámero, con 2 sépalos externos ovados, breves, y 3 internos obovados, los dos anteriores brevemente soldados en la base. La corola dialipétala posee 4 pétalos, 2 posteriores y 2 anteriores, provistos de una escama que en los pétalos posteriores es simétrica y asimétrica en los pétalos anteriores. El nectario floral, en ambos tipos de flores, se presenta fragmentado en 4 lóbulos nectaríferos, dos posteriores ovoideo-obtusos y dos anteriores conformes, menores. El androginóforo es glabro, en la parte superior se insertan 8 estambres de longitud desigual al igual que en C. grandiflorum. En las flores estaminadas el gineceo está reducido a un breve pistilodio. En las flores pistiladas se diferencian 8 estambres de menor longitud con las anteras indehiscentes, el gineceo es excéntrico con 3 carpelos, de3,5 (-6) mm long., un óvulo campilótropo por carpelo, ovario trígono-obovoideo, velloso, estilo general-mente más corto que los lóbulos del estigma.
Vascularización floral: El patrón de vascularización es similar al descripto para flores de C. grandiflorum (Fig. 3 A-F). Las diferencias encontradas sedan en la vascularización de los sépalos anteriores y en los lóbulos nectaríferos.

Fig. 3. Vascularización floral de U. chacoensis. A-Q: Cortes transversales sucesivos de una flor pistilada. R-V: Cortes transversales sucesivos de una flor estaminada. X: Corte longitudinal de botón floral de flor estaminada en preantesismostrando la posición de los transcortes. Escalas: A-W = 500 µm. Abreviaturas: sp: sépalo posterior; sei: sépalo externoizquierdo; sed: sépalo externo derecho; cspp: complejo sepalar-petalar posterior; pa: pétalo anterior; csp: complejo sepalarpetalar; hsc: haz sepalar central; sa: sépalo anterior; ec: estela central; hp: haz petalar; epp: escama pétalo posterior; pp:pétalo posterior; tc: traza carpelar; o: óvulo; oa: óvulo atrofiado; ov: ovario; l: lóbulo; Na: nectario anterior; Np: nectarioposterior; tc: traza central; tl: traza lateral; fe: filamento estaminal; tf: traza floemática; hcd: haz carpelar dorsal; hcl: hazcarpelar lateral; hcv; haz sepalar ventral; he: haz estaminal; le: lóbulo estigmático; e: estilo; hle: haz lóbulo estigmático. A-Q: Ferrucci et al. 2680 (CTES); R-W: Meza Torres et al. 214 (CTES); W: Ferrucci et al. 2680 (CTES).
En la parte anterior, de la sifonostela central dela flor, emergen 3 trazas, 1 central y 2 laterales;posteriormente las mismas se bifurcan constituyendo 6 trazas. Éstas junto con los haces marginalesprovenientes de los complejos sépalo-petalares delos sépalos externos derecho e izquierdo, van ainervar a los 2 sépalos anteriores y a los 2 pétalosanteriores respectivamente (Fig. 3 B-E).
Las trazas que vascularizan los nectarios posteriores provienen de la sifonostela ectofloica central, a partir de ésta se desprenden 4 ó 5 trazas floemáticas, que inervan la porción abaxial de cada lóbulo nectarífero, las que se vuelven a ramificar hasta llegar a 3 ó 4 capas subepidérmicas. Las trazas están representadas por miembros de tubos cribosos, células acompañantes y células parenquimáticas. Los nectarios anteriores no están inervados (Fig. 3E-G).
Con respecto a la vascularización de los verticilos fértiles el patrón es similar al descripto para C. grandiflorum, en las flores pistiladas (Fig. 3J-Q) y en las flores estaminadas (Fig. 3 R-V).
Thinouia mucronata Radlk. (Fig. 1 E, F)
Morfología floral: Flores reunidas en tirsos umbeliformes, inflorescencias parciales, pedunculadas. Las flores son actinomorfas, pequeñas, de 2,5-4mm long., blanquecino-verdosas. El cáliz escupuliforme, pentámero, sépalos soldados en un tercio basal, lóbulos ovados. La corola es pentámera, pétalos espatulados; glandulosos, con pelos pluricelulares lineares dispersos; escama basal soldada a la uña del pétalo, porción libre bipartida, ramas divergentes vellosas. El disco nectarífero es anular, glabro. El androceo con 5-8 estambres exertos en la flor estaminada, 2,25-4 mm long., con filamentos vellosos en la mitad basal, anteras glabras; pistilodio pubérulo. En las flores pistiladas los estambres menores, de1,5-1,75 mm long.; el gineceo brevemente estipitado, 3-carpelar, 1 óvulo campilótropo por carpelo inserto en la base del lóculo, ovario trígono-ovoideo, velloso, estilo filiforme, pubérulo, más breve o más largo que los lóbulos del estigma, éste trífido.
Vascularización floral: El tejido vascular del pedicelo constituye una sifonostela ectofloica circular, que en el receptáculo, adquiere forma estrellada, de la que se desprenden 5 trazas que previa inervación de los sépalos se bifurcan formando el complejo sépalo-petalar (Fig. 4 A-B). Un haz de cada uno de estos complejos sépalo petalar va a inervar cada uno de los sépalos, éstos se dividen dando lugar a un haz dorsal y dos marginales, todos son colaterales (Fig. 4 C-F). El otro haz del complejo denominado haz petalar se bifurca e ingresa al pétalo hasta finalizar en tres haces colaterales, uno dorsal y dos marginales, posteriormente continúa dividiéndose en varios haces vasculares pequeños que inervan la escama petalar (Fig. 4 G-H).
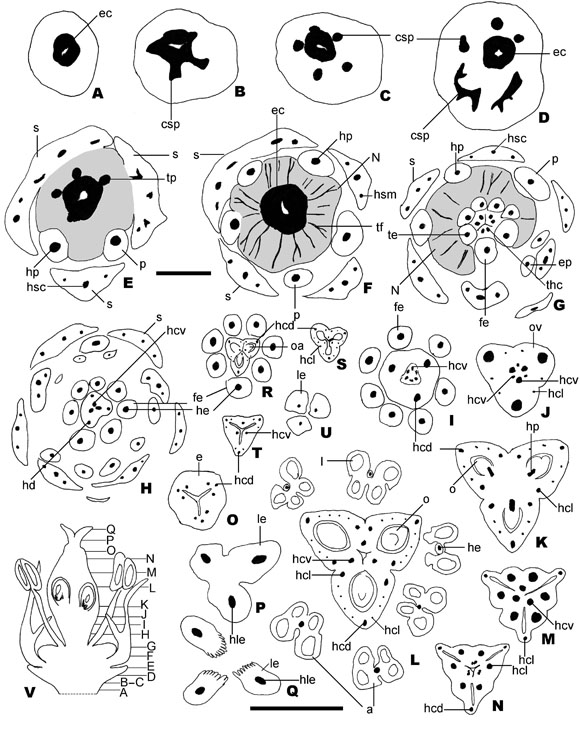
Fig. 4. Vascularización floral de Thinouia mucronata. A-Q: Cortes transversales sucesivos de una flor pistilada. R-U: Cortes transversales sucesivos de una flor estaminada. V: Corte longitudinal de un botón floral de flor pistilada mostrandola posición de los transcortes. Escalas: A-H = 500 µm; R-U = 200 µm I-Q, V = 500 µm. Abreviaturas: s: sépalo; csp: complejo sépalo-petalar; ec: estela central; p: pétalo; tp: traza petalar; hp: haz petalar; hsm: haz sepalar marginal; hsc: hazsepalar central; N: nectario; ep: escama petalar; hp: haz petalar; tf: trazas floemáticas; he: haz estaminal; hcl: haz carpelarlateral; hcd: haz carpelar dorsal; hcv: haz carpelar ventral; ov: ovario; o: óvulo; oa: óvulo atrofiado; e: estilo; le: lóbuloestigmático; hle: haz lóbulo estigmático; fe: filamento estaminal. A-V: Keller 5018 (CTES).
De la estela central a la altura del disco nectarífero, se desprenden entre 15-20 trazas floemáticas que se extienden y vuelven a ramificarse en el tejido nectarífero (Fig. 4 F-G). Estas trazas están constituidas por miembros de los tubos cribosos largos con paredes engrosadas, células acompañantes pequeñas y células parenquimáticasde mayor tamaño.
Por encima del nectario, la sifonostela central se fragmenta en 6 a 8 (en algunos casos 5) trazas concéntricas perifloemáticas, las que van a inervar a los filamentos estaminales, estos haces estaminales se extienden hasta el tejido conectivo (Fig. 4 G-I, L).
En la base del ovario, desde la estela central divergen 3 haces dorsales, 6 haces ventrales y numerosos hacecillos laterales, de menor calibre (Fig. 4 I-J).
Poco por encima, donde se diferencian los lóculos del ovario, se distinguen tres carpelos fusionados, inervados por 3 trazas dorsales y 3 trazas ventrales estas últimas formadas por la fusión de los 6 haces ventrales. Entre las trazas dorsales y las trazas ventrales se presentan numerosas trazas laterales que se extienden a lo largo de cada carpelo. De las trazas ventrales emergen 3 trazas que van a inervar los óvulos de cada carpelo, son los haces placentarios (Fig. 4 K-L y R-S).
Por encima de los lóculos del ovario se visualizan los hacecillos dorsales, 6 hacecillos laterales, resultado de la fusión de los hacecillos laterales menores, y 3 hacecillos ventrales; éstos se continúan en la pared del estilo (Fig. 4 M-O, T). Antes de llegar al estigma los hacecillos de cada carpelo se fusionan en un único haz, los que inervarán cada uno de los tres lóbulos del estigma (Fig.4 P-Q, U). El ovario es sinascidiado en aproximadamente la mitad de su longitud, luego simplicado, hemisimplicado próximo a la base de las tres ramas del estigma, en éste es asimplicado.
DISCUSIÓN Y CONCLUSIONES
Un elevado número de géneros de Sapindaceae presenta flores monosimétricas oblicuamente zigomorfas, carácter que comparte con Hippocastanaceae, familia incluida en Sapindaceae (Gadek et al., 1996), Vochysiaceae y Moringaceae (Ronse Decraene et al., 2000); mientras unos pocos géneros poseen flores actinomorfas con ciclos pentámeros alternos. En Sapindaceae la monosimetría floral está ligada a pérdida de órganos tales como pétalos, estambres y carpelos. El gineceo triangular tiene un fuerte impacto en la reorganización oblicua de la simetría de la flor, pérdida de órganos y desvíode los estambres (Ronse Decraene et al., 2000).
Cardiospermum grandiflorum, U. chacoensis (Paulliniinae) y T. mucronata (Thinouiinae) comparten el suministro vascular inicial, en estas especies ingresa a la flor una sifonostela central ectofloica. Con respecto a la inervación del perianto se reconocen tres patrones, en C. grandiflorum divergen 7 trazas (3 derechas y 3 izquierdas en el nivel inferior, poco más arriba 1 traza posterior y por encima 2 trazas anteriores); en U. chacoensis, 8 trazas (3 derechas y 3 izquierdas en el nivel inferior, poco más arriba 1 traza posterior y por encima 3 trazas anteriores) y en T. mucronata divergen 5 trazas responsables de la inervación del perianto. Con respecto a la vascularización del nectario, Thinouia se destaca por una profusa irrigación, estado del carácter queestaría asociado a la posición basal de este género enla tribu (Solís, inéd.). Por otra parte, las tres especiesanalizadas comparten el patrón de venación de losverticilos fértiles.
En Sapindaceae (incl. Hippocastanaceae) son comunes los pétalos provistos de una escama ventral (Radlkofer, 1895; Leinfellner, 1955, 1958). Las escamas en la superficie ventral de los pétalos son comúnmente interpretadas como parte de los pétalos (Leinfellner, 1954, 1958). En las tres especies aquí analizadas la escama está vascularizada, observación que coincide con lo citado por Radlkofer (1895) y por Leinfellner (1958) para otras especiesde la familia. En las eudicotiledóneas, en Rosídeas, clado eurosídeas II varios grupos presentan elaboraciones ventrales del pétalo (Endress & Matthews,2006a,b).
Aunque los nectarios florales, en las especies aquí analizadas, varían en la morfología desde un par de lóbulos nectaríferos corniculiformes posteriores en C. grandiflorum, 4 lóbulos, 2 posteriores y 2 anteriores, en U. chacoensis y un disco en T. mucronata, en todos los casos son extraestaminales y están inervados por trazas de floema que se desprenden de la estela central. En especies de Paullinieae, el néctar es secretado a través de nectarostomas (Solís, inéd.). La presencia del disco nectarífero extraestaminal es un carácter que aparece regularmente en Sapindales.
Las especies analizadas comparten el patrón de venación de los verticilos fértiles. Con respecto alandroceo, un carácter notable en especies de Paulliniinae es la diferencia en longitud que presentan los estambres, 3 altos anteriores, 2 medianos laterales y 3 cortos posteriores. Los hacecillos que inervan los estambres en C. grandiflorum y U. chacoensis varían de colaterales a perifloemáticos, mientras que en T. mucronata son siempre perifloemáticos. En la familia, los géneros con 8 estambres han perdido 2 en un proceso de reducción. La interpretación más plausible para Sapindaceae sería que han derivado de un androceo diplostémono ancestral, en la actualidad se observa este carácter en los géneros paleotropicales Blighia K. D. Koenig, Diploglottis Hook. f., Eriocoelum Hook. f., Laccodiscus Radlk. y Lecanodiscus Planch. ex Benth. (Radlkofer, 1895).
En las especies aquí estudiadas, el patrón de desarrollo del gineceo, con el ovario sinascidiado en aproximadamente la mitad de la longitud, luego simplicado, hemisimplicado próximo a los lóbulos del estigma y asimplicado en éstos últimos coincidecon lo citado por Weckerle & Rutishauser (2005) para especies de Paullinia, Serjania, Cardiospermum y Urvillea. Los lóculos se extienden en el estilo como 3 hendiduras angostas opuestas a los hacecillos dorsales, carácter que concuerda con lo observado en Koelreuteria paniculata Laxm. (Ronse Decraene et al., 2000).
El patrón de vascularización observado en C. grandiflorum concuerda en líneas generales con lo descripto por Nair & Joseph (1960) quienes analizaron la vascularización floral de C. halicacabum. Sin embargo, una de las diferencias observadas en esta última es la presencia de 4 hacecillos en el receptáculo del cual divergen 4 trazas. La vascularización de los verticilos fértiles es similar en ambas especies.
Con respecto a la valorización de los caracteres asociados con la vascularización, en algunos trabajos sobre morfología floral se estima que estos caracteres son evolutivamente más conservadores que otros referidos a la morfología externa (Puri,1951; Eyde, 1971); mientras que Carlquist (1969) y Schmid (1972) discuten el principio de que el sistema vascular sea casi siempre más conservador que los órganos que inerva. Sin embargo, Carlquist (1969) sugiere que las similitudes y diferencias en la vascularización floral deberían conservarse como un carácter de valor taxonómico.
Los resultados obtenidos en este trabajo apoyan la hipótesis sobre la tendencia evolutiva postulada para Paullinieae por Ferrucci & Anzótegui (1993). Thinouia con una vascularización floral más sencilla, respecto a Cardiospermum y Urvillea, que asociada a otros caracteres como flores actinomorfas, con cáliz y corola pentámeros, las escamas de los pétalos iguales entre sí, disco nectarífero, polen tricolporado y número cromosómico 2n = 28, sería un género con posición basal en la tribu.
Como corolario se debe enfatizar que los estudios estructurales comparativos significan un complemento importante para valorizar las relaciones filogenéticas en la tribu.
AGRADECIMIENTOS
A la Dra. Ana María González por el asesoramiento en el uso de los programas para digitalizar las imágenes, como también su ayuda en la interpretación de la vascularización floral. Este trabajo es parte de la tesis doctoral que será presentada por S. M. Solís en la Facultad de Ciencias Exactas, Físicasy Naturales de la Universidad Nacional de Córdoba, Argentina. Ha sido subsidiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 6000), por la Agencia Nacional de Promoción Científica, Tecnológica y de Innovación (ANPCyT-UNNE, PICTO 00096) y por la Secretaría General de Ciencia y Técnica de la Universidad Nacional del Nordeste (PI 038/06).
BIBLIOGRAFÍA
1. CARLQUIST, S. 1969. Toward acceptable evolutionary interpretations of floral anatomy. Phytomorphology 19: 332-362. [ Links ]
2. ENDRESS, P. K., P. BAAS & M. GREGORY. 2000. Systematic morphology and anatomy: 50 years of progress. Taxon 49: 401-434. [ Links ]
3. ENDRESS, P. K. & M. L. MATTHEWS. 2006a. First steps towards a floral structural characterization of the major rosid subclades. Plant Syst. Evol. 260: 223-251. [ Links ]
3. ENDRESS, P. K. & M. L. MATTHEWS. 2006b. Elaborate petals and staminodes in eudicots: Diversity, function, and evolution. Org. Divers. Evol. 6: 257-293. [ Links ]
4. EYDE, R. H. 1971. Evolutionary morphology: distinguishing ancestral structure from derived structure in flowering plants. Taxon 20: 63-73. [ Links ]
5. FERRUCCI, M. S. 2000a. Revisión de los géneros Cardiospermum y Urvillea para el neotrópico (Sapindaceae). Tesis Doctoral, Universidad Nacional de Córdoba. [ Links ]
6. FERRUCCI, M. S. 2000b. Cytotaxonomy of Sapindaceae with special reference to the tribe Paullinieae. Genet. Mol. Biol. 23: 941-946. [ Links ]
7. FERRUCCI, M. S & L. M. ANZÓTEGUI. 1993. El polen de Paullinieae (Sapindaceae). Bonplandia 6: 211-243. [ Links ]
8. GADEK, P. A., E. S. FERNANDO, C- J. QUINN, S. B. HOOT, T. TERRAZAS, M. C. SHEAHAN & M. W. CHASE. 1996. Sapindales: Molecular delimitation and infraordinal groups. Amer. J. Bot. 83: 802-811. [ Links ]
9. GONZÁLEZ, A. M & C. L. CRISTÓBAL. 1997. Anatomía y ontogenia de semillas de Helicteres Lhotzkyana (Sterculiaceae). Bonplandia 9: 287-294. [ Links ]
10. GUSTAFSON, M. H. G. 1995. Petal venation in the Asterales and related orders. Bot. J. Linn. Soc. 118: 1-18. [ Links ]
11. JOHANSEN, D. A. 1940. Plant microtechnique. McGraw-Hill Book Company, New York. [ Links ]
12. LEINFELLNER, W. 1954. Beiträge zur Kronblattmorphologie. I. Erythroxylon novogranatense. Österr. Bot. Z. 101: 428-434. [ Links ]
13. LEINFELLNER, W. 1955. Beiträge zur Kronblattmorphologie.V. Über den homologen Bau der Kronblattspreite und der Staubblattanthere bei Koelreuteria paniculata. Österr. Bot. Z. 102: 89-98. [ Links ]
14. LEINFELLNER, W. 1958. Über die peltaten Kronblätter der Sapindaceen. Österr. Bot. Z. 105: 443-514. [ Links ]
15. LUQUE, R., H. C. SOUSA & J. E. KRAUS. 1996. Métodos de coloração de Roeser (1972) - modificado - E. Kropp (1972), visando a substituição do azul de astra por azul de alcião 8GS ou 8GX. Acta Bot. Bras. 10: 199-212. [ Links ]
16. MULLER, J. & P. W. LEENHOUTS. 1976. A general survey of pollen types in Sapindaceae in relation to taxonomy. In: FERGUSON I. K. & J. MULLER (eds.) The evolutionary significance of the exine. Linn. Soc. Symp. Ser. 1: 407-445. [ Links ]
17. NAIR, N. C. & J. JOSEPH. 1960. Morphology and embryology of Cardiospermum halicacabum. J. Indian Bot. Soc. 39: 176-194. [ Links ]
18. PURI, V. 1951. The role of floral anatomy in the solution of morphological problems. Bot. Rev. 17: 471-533. [ Links ]
19. RADLKOFER, L. 1895. Sapindaceae. In: ENGLER, A. & K. Prantl (eds.), Nat. Pflanzenfam. 3, pp. 277-366. Engelmann, Leipzig. [ Links ]
20. RADLKOFER, L. 1931-1934. Sapindaceae. In: ENGLER, A. (ed.), Pflanzenr. 98 (IV, 165), pp. 1-1539. Wilhelm Engelmann, Leipzig. [ Links ]
21. RONSE DECRAENE, L. P., E. SMETS & D. CLINCKEMAILLIE. 2000. Floral ontogeny and anatomy in Koelreuteria with special emphasis on monosymmetry and septal cavities. Plant Syst. Evol. 223: 91-107. [ Links ]
22. SCHMID, R. 1972. Floral bundle fusion and vascular conservation. Taxon 21: 429-446. [ Links ]
23. WECKERLE, C. S. & R. RUTISHAUSER. 2005. Gynoecium, fruit and seed structure of Paullinieae (Sapindaceae). Bot. J. Linn. Soc. 147: 159-189. [ Links ]
Recibido el 2 de febrero de 2009
Aceptado el 2 de septiembre de 2009.

