










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.45 no.3-4 Córdoba jul./dic. 2010
ANATOMÍA Y MORFOLOGÍA
Indumento y modelos epidérmicos en especies argentinas de Elaphoglossum (Dryopteridaceae - Pteridophyta)
María del Carmen Lavalle1 y Mónica Rodríguez1
1 Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata.
Resumen: Este es un estudio comparativo sobre el indumento y los modelos epidérmicos de las siete especies argentinas del género Elaphoglossum: E. crassipes (Hieron.) Diels, E. gayanum (Fée) T. Moore, E. lindbergii (Mett. ex Kuhn) Rosenst., E. lorentzii (Hieron.) H. Christ, E. pachydermum (Fée) T. Moore, E. piloselloides (C. Presl) T. Moore y E. yungense de la Sota. Estos caracteres resultaron de valor diagnóstico a nivel específico. Los resultados obtenidos se aplican en una clave para la determinación de las especies.
Palabras clave: Elaphoglossum; Argentina; Indumento; Epidermis.
Summary: Indument and epidermal pattern in argentine species of the genus Elaphoglossum (Dryopteridaceae-Pteridophyta). This is a comparative study about indument and epidermical patterns of laminae carried out in seven argentine species of the genus Elaphoglossum: E. crassipes (Hieron.) Diels, E. gayanum (Fée) T. Moore, E. lindbergii (Mett. ex Kuhn) Rosenst., E. lorentzii (Hieron.) H. Christ, E. pachydermum (Fée) T. Moore, E. piloselloides (C. Presl) T. Moore y E. yungense de la Sota. These characters are diagnostic at the specific level. A key to identify the species was obtained from the results.
Key words: Elaphoglossum; Argentina; Indument; Epidermis.
INTRODUCCIÓN
Elaphoglossum es un género pantropical que está ubicado en la familia Dryopteridaceae Herter sobre la base de estudios morfológicos (Tryon & Tryon, 1982; Mickel, 1991, 1999) y filogenéticos (Smith et al., 2006; Moran et al., 2007). Las Dryopteridaceae están ampliamente distribuidas; su número de géneros difiere entre los autores, especialmente con respecto a los representantes del este de Asia, donde se ubica el centro de diversidad de la familia. Tryon & Tryon (1982) incluyen más de 50 géneros, 30 de ellos americanos; mientras que Li & Lu (2006) consideran cerca de 30-35 géneros.
A Elaphoglossum se le adjudicaron en tratados florísticos u obras generales (Sehnem, 1979; Mickel & Beitel, 1988; Proctor, 1989; Mickel & Smith, 2004), entre 400 y 600 especies, la mayoría neotropicales.
Las especies argentinas no han sido estudiadas comparativamente en conjunto. Son tratadas en el contexto de floras pteridológicas regionales, que incluyen una o varias especies, sin que haya una evaluación de los caracteres utilizados (Capurro, 1969; de la Sota, 1977, de la Sota et al., 1998). Aparecen además nombradas en diferentes catálogos (Ponce, 1996; Jørgensen & Leon-Yañez, 1999; Ponce, 2008). Por otra parte existen contribuciones sobre diversos rasgos morfológicos del esporófito (Christ, 1899; Bell, 1951, 1955; Nayar & Kaur, 1965; Nayar, 1966; Lloyd, 1970; Mickel & Atehortúa, 1980; Tryon and Lugardon, 1991; Lavalle & Rodríguez, 2006, 2007, 2009; Moran et al., 2007); así como también análisis filogenéticos moleculares, utilizando secuencias de ADN cloroplastidial (Rohuan et al., 2004; Skog et al., 2004).
Las plantas de Elaphoglossum presentan esporófitos de pequeño a mediano tamaño, frondas generalmente dimórficas, láminas simples, soros acrosticoides y esporas monoletes. Crecen en el NE argentino en la provincia de Misiones, en el NO (Jujuy, Salta, Tucumán), y desde las Sierras Centrales de Córdoba hasta las Sierras Australes de Buenos Aires. Viven en selvas montanas altas, medianas o bajas, perennifolias y en bosques de neblina; sobre rocas húmedas; desde el nivel del mar hasta los 4500 m.
En este trabajo se estudia en forma comparada la morfología y naturaleza del indumento presente en láminas y ejes estériles (tipo de escamas, forma, margen, ápice, color, textura, inserción y persistencia); los modelos epidérmicos de hipofilos y epifilos y los tipos de estomas mostrados en las 7 especies argentinas determinadas previamente (Lavalle & Rodríguez, 2009). Se presenta además una clave para la determinación de las especies, realizada en base a caracteres analizados en este trabajo y sumados a otros de trabajos previos.
Las especies estudiadas son: E. crassipes (Hieron.) Diels, E. gayanum (Fée) T. Moore, E. lindbergii (Mett. ex Kuhn) Rosenst., E. lorentzii (Hieron.) H. Christ, E. pachydermum (Fée) T. Moore, E. piloselloides (C. Presl) T. Moore y E. yungense de la Sota.
MATERIALES Y MÉTODOS
Se utilizó material de los herbarios BA, LP, MO, NY y SI (Holmgren et al., 1990), citados a continuación:
Elaphoglossum crassipes (Hieron.) Diels. ARGENTINA. Prov. Catamarca: Dpto. Andalgalá, 2-V-1915, Jörgensen 1496 (MO, SI). Prov. Jujuy: Dpto. Ledesma, camino Mesada de las Colmenas a Abra de Cañas, 17-III-1966, de la Sota 4414 (LP). Prov. Salta: Dpto. Santa Victoria, Camino Los Toldos a Lipeo, Vallecito, Quebrada Honda, 13-XI-1997, Cassau et al. 278 (LP); Los Toldos, Quebrada del Astillero, 17-III-86, Palací 496 (LP). Prov. Tucumán: Dpto. Chicligasta, Cuesta del Clavillo, IV-1939, Schreiter 96048 (SI).
Elaphoglossum gayanum (Fée) Moore. ARGENTINA. Prov. Buenos Aires: Pdo. Tandil, Tandil, 25-X-1970, Cabrera & Fabris 20871 (BA). Pdo. Tornquist, entre Santa Victoria y Rodeo Pampa, 13-II-1995, Deginiani et al. 755 (SI). Prov. Catamarca: Dpto. Ambato, Los Mogotes, 7-III- 1960, Hunziker 7215 (LP). Prov. Córdoba: Dpto. Punilla, Capilla del Monte, Cerro Uritorco, II-1933, Nicora 125 (SI). Prov. Jujuy: Dpto. Dr. M. Belgrano, Yala, cerros, 19-II-1940, Burkart & Troncoso 11366 (SI). Prov. La Rioja: Dpto. Gobernador Gordillo, Sierra de los Llanos (frente al Chamical) subiendo a la cima del cerro "El Morado", 16-X-1977, Birrum et al. 1141 (LP). Prov. Salta: Dpto. Guachipas, Pampa Grande, I-1897, Spegazzini s.n. (LP). Prov. San Luis: Dpto. Belgrano, Nogoli, X-1922, J. 131 Gez 2 (SI). Prov. Tucumán: Dpto. Chicligasta, Alpachiri, Estancia Los Pinos, 5-VI-1945, Capurro 43804 (BA); La Lagunita, 14-IV-1912, Venturi 488 (LP); s.loc., Castillón 27 (SI).
Elaphoglossum lindbergii (Mett. ex Kuhn) Rosenst. ARGENTINA. Prov. Jujuy: Dpto.Ledesma, camino a Valle Grande desde Ruta Nac. 34, km 30, 16-XII-1986, Zuloaga et al. 2937 (SI). Prov. Salta: Dpto. Santa Victoria, Los Toldos, Finca "El Nogalar", Terraza alta del Río Huaico, Cassau et al. 250 (LP); Parque Nac. Baritú, Arroyo Baritú, 4-X-1985, Brown 2017 (SI); Quebrada de Caspichangi, 25-I-1903, Lillo 2922 (SI). Prov. Tucumán: Dpto. Chicligasta, Alpachiri, Estancia Los Pinos, 5-VI-1945, Capurro 71 (BA, LP); La Quebrada Famaillá, VIII-1912, Castillón (SI 23988).
Elaphoglossum lorentzii (Hieron.) H. Christ. ARGENTINA. Prov. Córdoba: Dpto. San Alberto, Pampa de Achala, quebradas al W., 22-II-1965, Cabrera et al. 16625 (LP). Prov. Salta: Dpto. Santa Victoria, de Abra de Lizoite a Santa Victoria, 22-III-1982, Kiesling et al. 3074 (SI). Prov. Tucumán: Dpto. Chicligasta, Estancia Las Pavas, Pto El Bayo, 17-III-1924, Venturi 3219 (SI). Dpto. Tafí, Cumbres Calchaquíes: La Queñoa, 11-III-1912, Rodríguez 401 (BA, SI); 16-III-1974, Zardini 273 (LP); puerta del Río Blanco, I-1912, Castillón (SI 55552).
Elaphoglossum pachydermum (Fée) T. Moore. ARGENTINA. Prov. Misiones: Dpto. San Ignacio, Parque Provincial Teyú Cuaré, Peñón del Teyú Cuaré, 28-IX-1998, Biganzoli & Giraldo-Cañas 394 (SI); Teyú-Cuaré, cerca de San Ignacio, II-1910, Wagner s.n. (SI 24018).
Elaphoglossum piloselloides (C. Presl) T. Moore. ARGENTINA. Prov. Catamarca: Dpto.Andalgalá, Esquina Grande, 2-V-1915, Jørgensen 1495 (BA, SI). Prov. Jujuy: Dpto. Dr. M. Belgrano, Quebrada de Jaire, 16-XI-1978, Cabrera et al. 29920 (SI). Dpto. Ledesma, llegando a Abra de Cañas, 9-IV-1971, Legname & Cuezzo 7942 (LP). Prov. Salta: Dpto. Capital, Quebrada San Lorenzo, 25-VIII-1985, Palací 161 (LP). Prov. Tucumán: Dpto. Burroyacu, Cerro del Durazno, 4-III-1914, Monetti 2061 (LP). Dpto. Chicligasta, Alpachiri, Arroyo Las Pavas, 8-VI-1945, Capurro 43802 (BA).
Elaphoglossum yungense de la Sota. ARGENTINA. Prov. Jujuy: Dpto. Capital, El Cucho, Quebrada Aº Tacanas, 10-III-1966, de la Sota 4353. Dpto. Dr. M. Belgrano, El Cucho, Quebrada del Río Reyes, 2-VI-1903, Holmberg (SI 24009). Dpto. Ledesma, camino Mesada de las Colmenas a Abra de Cañas, 17-III-1966, de la Sota 4408 (LP). Prov. Salta: Dpto. Orán, Aguas Blancas, en la ladera de un salto de la Quebrada El Nogalar, 23- VII- 1986, Palací 696 (LP). Dpto. Santa Victoria, Baritú, camino desde el poblado a Zenta, 13-VII-1999, Ganem et al. 136 (LP). Prov. Tucumán: s. loc., 10-III-1907, Hauman-Merck 24012 (SI).
Para analizar el indumento se ablandaron los ejes en una solución de éter monobutílico del etilénglicol diluido 1:1 en agua, durante una semana a temperatura ambiente. El indumento de las láminas se observó en material diafanizado.
Para estudiar los modelos epidérmicos de la lámina y los estomas, se rehidrataron trozos de lámina, con agua y unas gotas de detergente. Se aclararon con hidróxido de sodio 5% acuoso, se blanquearon con hipoclorito de sodio comercial diluido 1:1 en agua y se colorearon con TBO (azul de toluidina O) 1% acuoso.
Las ilustraciones se realizaron con un microscopio Wild M-20 y un microscopio estereoscópico Nikon SM Z 1000, ambos equipados con cámara de dibujo. Las fotografías fueron tomadas con una cámara Nikon Optical Zoom 6,3 Mp. 10 X Zoom con microscopio Olympus BX51 equipado con cámara digital Olympus DP71.
La terminología relacionada con el indumento es la propuesta por Lellinger (2002), la utilizada para describir las células epidérmicas, la de Rolleri y Deferrari (1986) y para los tipos estomáticos adultos, la de Prabhakar (2003).
RESULTADOS
Indumento
En las especies argentinas de Elaphoglossum el indumento está representado por escamas y además a veces, por tricomidos.
Los rizomas son densamente escamosos, llevan escamas basifijas ó peltadas, lanceadas, ovadas ó deltoides, de márgenes subenteros, a veces con unas pocas lacinias, bicolores o concolores, canela, castaño ó atrocastaño y textura papirácea ó cartácea. Las escamas pueden estar casi totalmente esclerosadas o estar constituidas por células engrosadas, esclerificadas y oscuras, localizadas en distintas áreas.
Las escamas del estípite son patentes y concolores en la mayoría de las especies. En algunas especies las escamas basales presentan contorno similar a las del rizoma pero con margen más lacerado, mientras las escamas distales son semejantes a las de la lámina.
Las láminas son subglabras en algunas especies, escamosas en otras y ocasionalmente llevan tricomidios. Las escamas presentan gran diversidad morfológica, desde formas deltoides con margen eroso ó laciniado hasta escamas reducidas (estrelladas, aracniformes). El indumento es superficial y a veces sobresale del margen; puede localizarse en la costa y/o en los márgenes.
Elaphoglossum crassipes. Los rizomas (Fig. 1, A) están tapizados por escamas basifijas, atrocastañas, lustrosas, cartáceas, lanceadas, con bases cordadas, ápices largamente acuminados y márgenes erosos. Los estípites llevan escamas persistentes, fibrilosas y translúcidas con un área color canela; las escamas basales (Fig. 1, G) son patentes y semejantes en forma a las del rizoma pero de márgenes subenteros y ápices color canela; las de la parte media (Fig. 1, E) son ovadas, con bases obtusas, ápices agudos, márgenes erosos y, ápices y parte central color canela; las de la parte distal (Fig. 1, D) son adpresas, oblongas, con bases y ápices obtusos, márgenes lacerados y color canela en la parte central. Las costas (Fig. 2, B) llevan abaxialmente abundantes escamas persistentes, adpresas y bicolores. El indumento de las láminas sobresale del margen (Fig. 2, B) y está formado en ambas superficies por abundantes escamas deltoides translúcidas, constituidas por células de formas y contornos irregulares similares a piezas de rompecabezas (Fig. 2, A). Los epifilos (Fig. 1, C) presentan escamas basifijas con bases cordadas y márgenes erosos o irregulares, distribuidas de manera uniforme; los hipofilos están tapizados por escamas imbricadas, peltadas con márgenes laciniados (Fig. 1, B; Fig. 2, C-E), presentan además escamas estrelladas (Fig. 1, F).
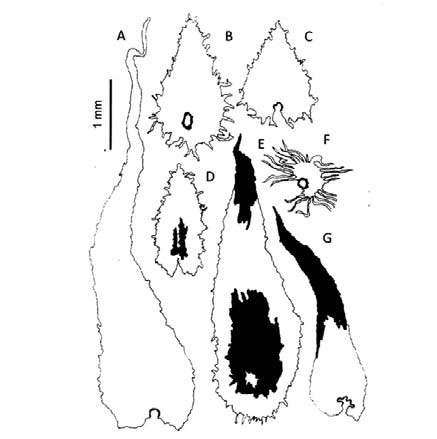
Fig. 1. A-G: Elaphoglossum crassipes. A: escamas del rizoma. B, F: escamas del hipofilo. C: escama del epifilo. D, E, G: escamas del estípite.
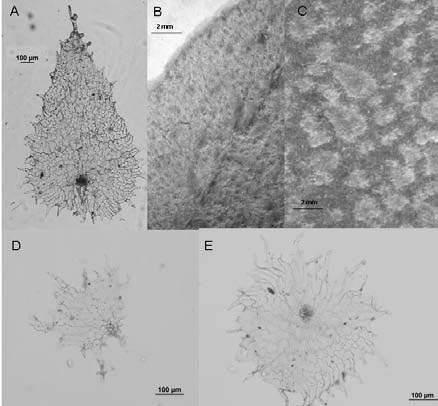
Fig. 2. A-E: Elaphoglossum crassipes. A, D, E: escamas del hipofilo. B: Hipofilo, costa y margen escamosos. C: detalle del hipofilo escamoso. A-B: de la Sota 4414 (LP). C: Cassau et al. 278 (LP). D-E: Palací 496 (LP).
Elaphoglossum gayanum. Presenta rizomas con escamas abundantes y persistentes, estípites con escamas deciduas y dispersas, costas glabras y, láminas casi glabras. Las escamas del rizoma (Fig. 3, A, C; Fig. 4, B) están formadas por células rectangulares largas de paredes delgadas; son patentes, peltadas, bicolores, canelas con la parte distal castaña, con frecuencia lustrosas, papiráceas, lanceadas a deltoides anchas, con ápices agudos y márgenes subenteros. Los estípites llevan escamas peltadas o basifijas constituidas por células poligonales de paredes rectas, patentes, bicolores, canelas con un área distal castaña, ovadas anchas, con márgenes algo laciniados (Fig. 3, B; Fig. 4, A). Los epifilos son glabros y los hipofilos casi glabros, con algunas escamas translúcidas, dispersas, caedizas, aracniformes (Fig. 3, D; Fig. 4, C) y estrelladas (Fig. 3, E).
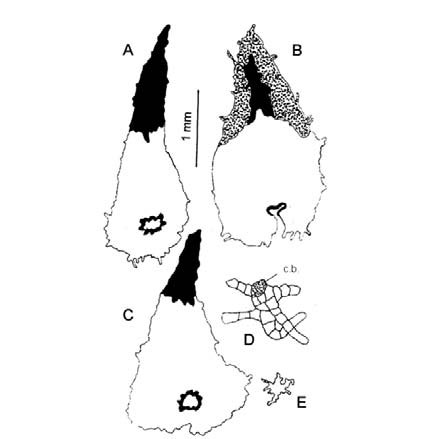
Fig. 3. A-E: Elaphoglossum gayanum. A, C: escamas del rizoma. B: escama del estípite. D-E: escamas del hipofilo.
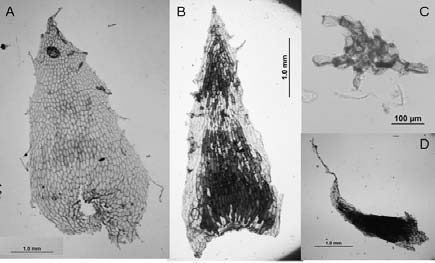
Fig. 4. A-D: Elaphoglossum gayanum. A: escama del estípite. B, D: escamas del rizoma. C: escama del hipofilo. A, C: Castillón 27 (SI). B, D: Venturi 488 (LP).
Elaphoglossum lindbergii. Los rizomas están totalmente tapizados por escamas lustrosas de textura papirácea, constituidas por células cortas en la base y largas en el resto de la superficie (Fig. 6, D). Los estípites (Fig.6, A) presentan escamas patentes, persistentes, esparcidas en toda su longitud pero con mayor densidad en la parte basal, las de la parte distal del estípite son similares a las de la costa. Las escamas de los rizomas (Fig. 5, A-B; Fig.6, D) así como las de los estípites (Fig. 5, C-D) son basifijas, concolores, castañas, aciculares con bases auriculadas ó deltoides angostas con inserción basal y, siempre con ápices largamente acuminados de márgenes irregulares. Las costas (Fig. 5, E, F, J; Fig. 6, C) y los márgenes (Fig. 5, G-I; Fig. 6, B) presentan escamas patentes, basifijas, deltoides, con ápices largamente acuminados y márgenes subenteros a enteros; las de la costa son deciduas y de mayor tamaño que la de los márgenes. Los epifilos y los hipofilos llevan escasos tricomas translúcidos; sobre los epifilos se encuentran escamas peltadas, orbiculares con márgenes irregulares (Fig. 5, K) y; sobre los hipofilos tricomidios, pelos pequeños estrellados, ramificados ó dendroides (Fig. 5, L); en ambas caras hay pelos bifurcados en forma de "Y" (Fig. 5, M).

Fig. 5. A-M: Elaphoglossum lindbergii. A, B: escamas del rizoma. C, D: escamas del estípite. F, J: escamas de la costa. G-I: escamas del margen. K: escamas del epifilo. L: tricomidios del hipofilo. M: pelos del epifilo e hipofilo.
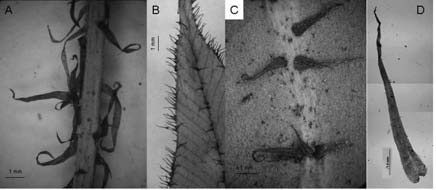
Fig. 6. A-D: Elaphoglossum lindbergii. A: parte del estípite con escamas. B: costa y margen de la lámina con escamas. C: costa con escamas. D: escama del rizoma. B, D: Zuloaga et al. 2937 (SI). A, C: Capurro 71.
Elaphoglossum lorentzii. Presenta rizomas, estípites y epifilos escamosos. Los rizomas (Fig. 7, AD; Fig.8, D) llevan abundantes escamas fibrilosas de paredes gruesas, algunas basifijas con base cordada y otras peltadas con base obtusa, concolores, castañas y de textura cartácea, ovadas a deltoides, con ápices agudos y márgenes irregulares. Las escamas de los estípites, patentes, persistentes y muy próximas entre sí (Fig.8, A) son peltadas, concolores, canelas, ovadas, lanceadas, con márgenes dentados a laciniados (Fig. 7, E- F). Los epifilos presentan distribuidas en toda la superficie (Fig. 8, C) escamas peltadas de células alargadas, translúcidas, deltoides con márgenes irregularmente dentados con algunas lacinias que sobresalen del margen (Fig. 7, G-H; Fig. 8, F); además se han encontrado tricomidios (Fig. 8, G). Los hipofilos (Fig. 7, I-J) son casi glabros o glabrescentes, con escasas escamas similares a las del epifilo; los márgenes presentan escamas clocleariformes (Fig. 8, B, E).
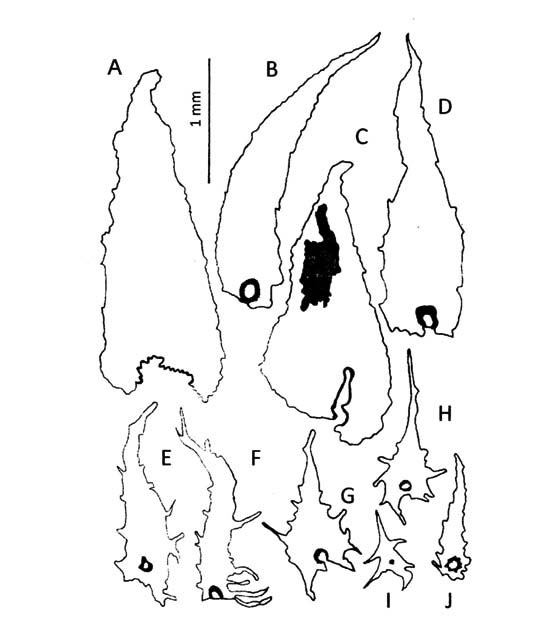
Fig. 7. A-J: Elaphoglossum lorentzii. A-D: escamas del rizoma. E-F: escamas del estípite. G-H: escamas del epifilo. IJ: escamas del hipofilo.
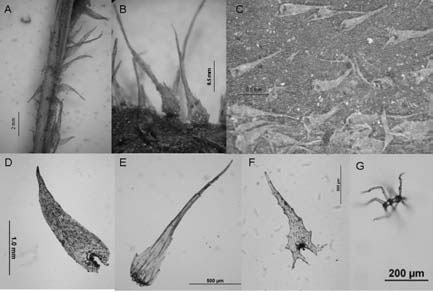
Fig. 8. A-G: Elaphoglossum lorentzii. A: parte del estípite con escamas. B: margen con escamas. C: epifilo escamoso. D: escama del rizoma. E: escama del margen. F: escama del epifilo. G: tricomidio del epifilo. A, C, D: Zardini 273 (LP). B, E-G: Castillón (SI 55552).
Elaphoglossum pachydermum. Tiene rizomas con abundantes escamas, estípites con escaso indumento, epifilos glabros e hipofilos con escamas no visibles a simple vista . Las escamas del rizoma (Fig. 9, A, F) son peltadas, persistentes, concolores, atrocastañas, lustrosas, papiráceas, clatradas, ovadas, con ápices acuminados y márgenes subenteros con 2-3 prolongaciones aisladas largas. Los estípites llevan en forma aislada a lo largo de toda su longitud, escamas basifijas, patentes, deciduas, concolores, castañas, deltoides ó irregulares, con bases auriculadas y márgenes laciniados en todo su contorno (Fig. 9, B, D; Fig. 10, B); además se han hallado tricomidios, diminutos pelos simples (Fig. 9, C). Las costas y los hipofilos presentan el mismo tipo de indumento (Fig. 10, A), escamas patentes, irregulares, laciniadas con cuerpos prolongados en brazos cortos uniseriados (Fig. 9, G, H; Fig. 10, D, E); y escamas aracniformes, concolores, castañas (Fig. 9, E; Fig. 10, C).
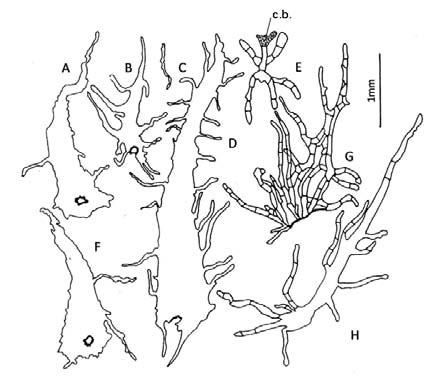
Fig. 9. A-H: Elaphoglossum pachydermum. A, F: escamas del rizoma. B, D: escamas del estípite. C: tricomidio del estípite. E, H: escamas de la costa y del hipofilo. G: protoescama de la costa y del hipofilo.

Fig. 10. A-E: Elaphoglossum pachydermum. A: costa e hipofilo con escamas no visibles a simple vista. B: escama del estípite. C-E: escamas de la costa. A, E: Biganzoli & Giraldo-Cañas 394 (SI). B, C-D: Wagner s.n. (SI 24018).
Elaphoglossum piloselloides. Presenta los rizomas, los ejes y las láminas densamente escamosos. Los rizomas (Fig. 11, A, B, H) llevan escamas basifijas, bicolores, castañas con la parte basal canela, papiráceas, lanceadas, ovadas con base cordada, ápices acuminados y márgenes subenteros. Los estípites presentan escamas patentes y persistentes, muy próximas entre si a lo largo de toda su longitud (Fig. 12, A); las escamas de las costas también son patentes. Un carácter distintivo de esta especie es la presencia de escamas similares en estípites (Fig. 11, CD; Fig. 12, B), costas (Fig. 11, I; Fig. 12, D), hipofilos (Fig. 11, J-K) y epifilos (Fig. 11, E; Fig. 12, E): escamas concolores, canelas, patentes, clatradas, cocleariformes con ápices largamente acuminados y márgenes regularmente dentados y reflexos en la parte basal de la escama, siendo irregularmente dentados en los estípites. En los epifilos (Fig. 12, C) las escamas sobresalen del margen mientras en los hipofilos están distribuidas espaciadamente; los hipofilos tienen además tricomidios (Fig. 11, F-G).
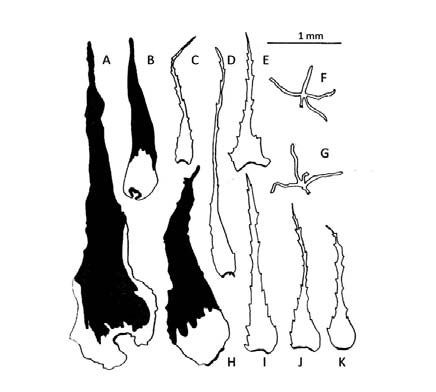
Fig. 11. A- K: Elaphoglossum piloselloides. A, B, H: escamas del rizoma. C, D: escamas del estípite. E: escama del epifilo. F, G: tricomidios del hipofilo. I: escama de la costa. J, K: escamas del hipofilo.
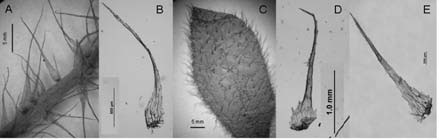
Fig. 12. A- E: Elaphoglossum piloselloides. A: parte del estípite con escamas. B: escama del estípite. C: epifilo con escamas en la superficie y en el margen. D: escama de la costa. E: escama del epifilo. A-C: Legname & Cuezzo 7942 (LP). D: Monetti 2061 (LP). E: Palací 161 (LP).
Elaphoglossum yungense. Se caracteriza por presentar rizomas y ejes escamosos, epifilos e hipofilos con diminutas escamas no visibles a simple vista. Los rizomas están totalmente tapizados en forma imbricada por escamas basifijas, atrocastañas, lustrosas, de textura cartácea, formadas por células rectangulares largas de paredes engrosadas (Fig. 14, A, D). Predominan las formas deltoides angostas, con ápices acuminados y márgenes subenteros con unas pocas lacinias localizadas generalmente en la parte basal (Fig. 13, A-B); en menor cantidad se encontraron escamas deltoides anchas, con ápices obtusos, bases con algunos pliegues y márgenes irregulares (Fig. 13, C, D, K; Fig. 14, D). Las escamas de los estípites están formadas por células rectangulares, de paredes engrosadas las de la parte basal y de paredes delgadas el resto de la superficie (Fig. 14, E); son deciduas, peltadas, patentes, translúcidas ó bicolores, canelas con castaño y forman una cobertura densa en la parte basal. Algunas escamas son semejantes en forma a las del rizoma aunque de menor tamaño (Fig. 13, M); mientras otras tienen formas irregulares con largas lacinias o prolongaciones en casi todo su contorno (Fig. 13, F, L); además se encontraron escamas estrelladas (Fig. 13, E, N). Las costas (Fig. 13, G, J, O; Fig. 14, B, F) llevan escamas patentes, concolores, canelas, ovadas, deltoides, con ápices acuminados que culminan en una cabezuela glandular y márgenes subenteros; se encontraron también escamas estrelladas (Fig. 14, H) y escamas deltoides con márgenes laciniados (Fig. 14, F). Los epifilos (Fig. 13, H-I) presentan escasas escamas pequeñas difusamente distribuidas, adpresas, translúcidas con el centro algo oscuro, estrelladas, con cuerpo paucicelular prolongado en brazos de distinta longitud. Los hipofilos están laxamente cubiertos por escamas estrelladas; además presentan tricomidios deciduos, translúcidos, dendriformes, que tienen una base globosa glandular resinosa visible como un punto rojo con márgenes subenteros; se encontraron también escamas estrelladas (Fig. 14, H) y escamas deltoides con márgenes laciniados (Fig. 14, F). Los epifilos (Fig. 13, H-I) presentan escasas escamas pequeñas difusamente distribuidas, adpresas, translúcidas con el centro algo oscuro, estrelladas, con cuerpo paucicelular prolongado en brazos de distinta longitud. Los hipofilos están laxamente cubiertos por escamas estrelladas; además presentan tricomidios deciduos, translúcidos, dendriformes, que tienen una base globosa glandular resinosa visible como un punto rojo con lupa de mano, rasgo que caracteriza a esta especie (Fig. 13, P-Q; Fig. 14, B, G, I). Los márgenes llevan escamas estrelladas (Fig- 14, C).
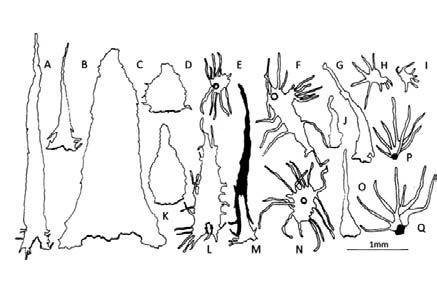
Fig. 13. A- Q: Elaphoglossum yungense. A-D, K: escamas del rizoma. E-F, L-N: escamas del estípite. G, J, O: escamas de la costa. H, I: escamas del epifilo. P, Q: tricomidios del hipofilo.
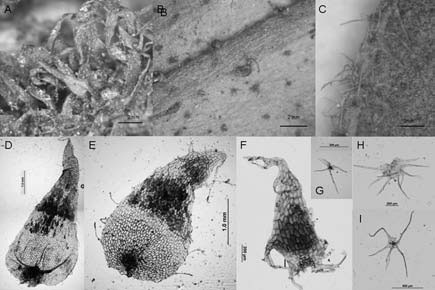
Fig. 14. A-I: Elaphoglossum yungense. A: rizoma con escamas. B: costa con escamas eglandulares e hipofilo con escamas glandulares. C: margen de la lámina con escamas. D: escama del rizoma. E: escama del estípite. F, H, I: escamas de la costa. G: tricomidio en hipofilo. A, E, H: de la Sota 4408 (LP). G, B: Palací 696 (LP). D, F: de la Sota 4353 (LP). I, C: Ganem et al. 136 (LP).
Modelos epidérmicos
Las epidermis de las láminas presentan modelos poligonales típicos y modelos sinuosos con ondulaciones que varían en amplitud, profundidad y frecuencia. También cambia, en las distintas especies, el tamaño de las células epidérmicas y se presentan células isodiamétricas ó más largas que anchas. La relación largo/ancho de las células se expresa mediante el radio celular.
Elaphoglossum crassipes. Epifilos con células subpoligonales de contornos casi rectos; hipofilos sinuosos con ondas curvas de escasa amplitud y poca profundidad; ambos con un radio celular 1,5:1 (Fig. 15, A-B).
Elaphoglossum gayanum. Epifilos con células poligonales de paredes rectas, con un radio celular 2-3:1; hipofilos sinuosos frecuentes, con ondas curvas de escasa amplitud y poca profundidad (Fig. 15, C-D).
Elaphoglossum lindbergii. Epifilos e hipofilos sinuosos; epifilos con ondas curvas uniformes poco profundas y amplias, células con un radio celular 1-3:1; hipofilos con células irregulares tipo puzzle con ondas curvas profundas (Fig. 15, E-F).
Elaphoglossum lorentzii. Epifilos subsinuosos de células isodiamétricas con radio celular 1:1 y células alargadas con radio celular 1,5-2:1, con ondas curvas de amplitud variable pero poco profundas; hipofilos sinuosos con radio celular 2-3:1, con ondas curvas profundas y amplias (Fig. 15, G-H).
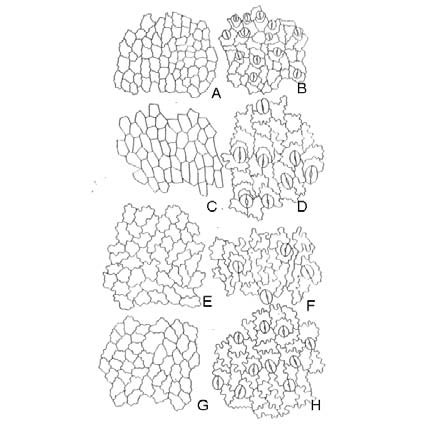
Fig. 15. Modelos epidérmicos y estomas en especies de Elaphoglossum. A-B: E. crassipes. C-D: E. gayanum. E-F: E. lindbergii. G-H: E. lorentzii. A, C, E, G: superficie adaxial. B, D, F, H: superficie abaxial.
Elaphoglossum pachydermum. Epifilos e hipofilos muy similares entre sí, sinuosos, con ondas curvas uniformes, de escasa amplitud y poca profundidad, distantes en el epifilo y frecuentes en el hipofilo, con células alargadas con radio celular 2:1 y 1-3:1, respectivamente (Fig. 16, A-B).
Elaphoglossum piloselloides. Epifilos e hipofilos sinuosos, con ondas curvas y angulosas, amplias y profundas; epifilos con células isodiamétricas o algo más largas que anchas, tipo puzzle, con sinuosidades distantes; hipofilos con radio celular 2-4:1 y con sinuosidades frecuentes y subsinuosidades (Fig. 16, C-D).
Elaphoglossum yungense. Epifilos subsinuosos e hipofilos sinuosos con ondas curvas de poca amplitud y profundidad, distantes en el epifilo y frecuentes en el hipofilo (Fig. 16, E-F).

Fig. 16. Modelos epidérmicos y estomas en especies de Elaphoglossum. A-B: E. pachydermum. C-D: E. piloselloides. E-F: E. yungense. A, C, E: superficie adaxial. B, D, F: superficie abaxial.
Estomas
En vista superficial los estomas tienen contorno circular en E. crassipes y, son elípticos y 1,5 veces más largos que anchos en el resto de las especies. Están rodeados por células vecinas que guardan disposiciones características: dos células orientadas perpendicularmente respecto de las oclusivas, tres células, o bien 4-5 células, rodeando los estomas.
En las especies estudiadas los estomas son diacíticos, anomocíticos y, más raramente, anisocíticos (Prabhakar, 2003).


DISCUSIÓN Y CONCLUSIONES
El indumento es utilizado en las floras regionales, como un rasgo sistemático en la determinación de las especies. Bell (1955) reconoce que la forma de las escamas es un carácter constante para las especies de Elaphoglossum, justificando así el gran valor diagnóstico que es asignado en la taxonomía del género. Este autor muestra la variación presente en las escamas de la lámina y los distintos grados de complejidad de las mismas dentro del género. Posteriormente Mickel & Atehortúa (1980) también consideran que la taxonomía del género se basa principalmente en las escamas del rizoma y de la lámina y enumeran la diversidad morfológica de las escamas con algunos ejemplos a nivel de sección o subsección.
• La presencia o ausencia, la densidad, la ubicación, la morfología y la naturaleza del indumento son rasgos taxonómicos de variación específica.
• Todas las especies argentinas tienen los rizomas tapizados por escamas. Así, E. gayanum y E. pachydermum, ubicadas por Lavalle & Rodríguez (2009) en la subsección Pachyglossa presentan escamas persistentes y abundantes; sin embargo, Mickel y Atehortúa (1980) citan para esta subsección rizomas con escamas deciduas que los tornan glabros.
• Las escamas de los estípites son patentes en todas las especies, siendo adpresas en la parte distal de los mismos en E. crassipes. Son deciduas en E. gayanum, E. pachydermum y E. yungense y persistentes en el resto de las especies. Las costas están totalmente tapizadas por escamas en E. crassipes y son glabras en E. gayanum.
• El indumento de las láminas es más denso con respecto al de los estípites y está formado por escamas más disectas en el hipofilo que en epifilo; esto coincide con lo expuesto por Mickel & Smith (2004). Es diferente en epifilo e hipofilo, solamente E. piloselloides presenta escamas similares en ambas superficies; sin embargo Mickel & Smith (2004) encontraron el mismo tipo de escamas en haz y envés, en todas las especies por ellos estudiadas.
• Las autoras coinciden con Mickel & Smith (2004) en que las láminas subglabras llevan escamas reducidas a diminutos tricomidios, si bien Mickel & Atehortúa (1980) hablan de ausencia de pelos multicelulares en la fronda de Elaphoglossum.
• Los epifilos son glabros en E. gayanum y E. pachydermum, siendo escamosos en el resto de las especies. Sin embargo, de la Sota (1977) registra como glabrescente o con pocas escamas hialinas al epifilo de E. lorentzii y, como glabro al epifilo y al hipofilo de E. lindbergii.
• Los hipofilos de E. crassipes presentan escamas formando un tapiz y los de E. piloselloides escamas distribuidas espaciadamente; en el resto de las especies los hipofilos son subglabros o llevan escamas no visibles a simple vista.
• Los hipofilos con escamas laciniadas con pronunciados procesos laterales irregularmente distribuidos, observados en E. pachydermum, coinciden con los presentados por Mickel (1995) para la sección Elaphoglossum, categoría a la que pertenece esta especie (Lavalle & Rodríguez, 2009).
• Las autoras coinciden con Roux (1982) en que los puntos rojizos observados con lupa de mano en los hipofilos de E. yungense representan las células basales de escamas deciduas extremadamente reducidas.
• Elaphoglossum crassipes es fácilmente distinguible por las escamas atrocastañas lustrosas de los rizomas, por el envés de las láminas totalmente tapizado por escamas translúcidas y por las costas con escamas adpresas.
• Elaphoglossum lindbergii se caracteriza por presentar escamas castañas visibles a simple vista en la costa y en el margen, que contrastan con el color verde claro de la superficie de la lámina.
• Elaphoglossum piloselloides presenta escamas superficiales cocleariformes que coinciden morfológicamente con las escamas linear lanceoladas de márgenes enrollados y semejantes a pelos, descritas para la sección Setosa, por Mickel & Atehortúa (1980).
• Los modelos epidérmicos resultan un carácter diagnóstico a nivel específico. Los epifilos presentan modelos poligonales, subpoligonales y sinuosos; mientras que los hipofilos son sinuosos en todas las especies.
BIBLIOGRAFÍA
1. BELL, P. R. 1951. Studies in the genus Elaphoglossum. III. Anatomy of the rhizome and frond. Ann. Bot. (London) II 15: 347-357. [ Links ]
2. BELL, P. R. 1955. Studies in the Genus Elaphoglossum. IV. The Morphological Series in the Genus and their Phylogenetic Interpretation. Ann. Bot., Lond., N. S. 19: 173-199. [ Links ]
3. CAPURRO, R. H. 1969[1968]. División Pteridophyta. In: CABRERA, A. L. (ed.), Flora de la Provincia de BuenosAires. Colecc. Ci. Inst. Nac.Tecnol. Agropecu. 4: 123 -146. [ Links ]
4. CHRIST, H. 1899. Monographie des Genus Elaphoglossum. Denkschr. Schw. Naturf. Ges. 36: 1-159. [ Links ]
5. DE LA SOTA, E. 1973. Sinopsis de las Pteridofitas del Noroeste de Argentina, II. Darwiniana 18: 240-248. [ Links ]
6. DE LA SOTA, E. 1977. Elaphoglossum. In: CABRERA, A. L. (ed.), Flora de la Provincia de Jujuy. Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 13: 230-240. [ Links ]
7. DE LA SOTA, E., M. PONCE, M. MORBELLI & L. CASSÁ DE PAZOS. 1998. Pteridophyta. In: CORREA, N (ed.). Flora Patagónica. Colecc. Ci. Inst. Nac. Tecnol. Agropecu. I: 282-369. Buenos Aires. [ Links ]
8. HOLMGREN, P. K., N. H. HOLMGREN & L. BARNETT. 1990. Index Herbariorum, Part I, Herbaria of the World, ed. 8. New York Botanical Garden, Bronx, New York. [ Links ]
9. JØRGENSEN, P. M. & S. LEON-YAÑEZ (eds.). 1999. Catalogue of the Vascular Plants of Ecuador: 128-134. [ Links ]
10. LAVALLE, M.C. & M. RODRÍGUEZ. 2006. Indumento en especies argentinas de Elaphoglossum Schott ex Sm. (Lomariopsidaceae- Pteridophyta). IX Jornadas de Ciencias Naturales del Litoral, Libro de resúmenes: 127. [ Links ]
11. LAVALLE, M.C. & M. RODRÍGUEZ. 2007. Arquitectura foliar y otros caracteres vegetativos en especies argentinas de Elaphoglossum Schott ex Sm. (Lomariopsidaceae - Pteridophyta). Bol. Soc. Argent. Bot. 42, Supl.: 107. [ Links ]
12. LAVALLE, M.C. & M. RODRÍGUEZ. 2009. Taxonomía de las especies argentinas de Elaphoglossum (Dryopteridaceae - Pteridophyta). Darwiniana 47:125-139. [ Links ]
13. LELLINGER, D. B. 2002. A Modern Multilingual Glossary of Taxonomic Pteridology. Pteridologia 2A: 1-364. [ Links ]
14. LI, C.-X. & C. G. LU. 2006. Phylogenetic analysis of Dryopteridaceae based on chloroplast rbcL sequences. ActaPhytotaxonomica Sinica, 44, 503-15. [ Links ]
15. LLOYD, R. M. 1970. Asurvey of some morphological features of the genus Elaphoglossum in Costa Rica. Amer. Fern J. 60: 73-83. [ Links ]
16. MICKEL, J. T. 1991. Pteridophyta of Peru, IV, 17. Dryopteridaceae. Fieldiana, Bot. 27: 98-165. [ Links ]
17. MICKEL, J. T. 1995. Elaphoglossum. In: DAVIDSE, G; M. SOUZA & S. KNAPP (eds.), Flora Mesoamericana 1, pp. 250-283. Universidad Nacional Autónoma de México. México, D. F. [ Links ]
18. MICKEL, J. T. 1999. Elaphoglossum. In: BERRY, P.; B. HOLST & K. YATSKIEVYCH (eds.), Flora of the Venezuelan Guayana. vol. 2. pp. 89-101. Missouri Botanical Garden Press, St. Louis. [ Links ]
19. MICKEL, J. T. & L. ATEHORTÚA.1980. Subdivision of the genus Elaphoglossum. Amer. Fern J. 70: 47-68. [ Links ]
20. MICKEL, J. T. & J. M. BEITEL. 1988. Pteridophyte flora of Oaxaca, Mexico. Mem. New York Bot. Gard. 46: 166-186. [ Links ]
21. MICKEL, J. T. & A. R. SMITH. 2004. The Pteridophytes of Mexico. Mem. New York Bot. Gard. 88: 282-315. [ Links ]
22. MORAN, R. C., J. GARRISON HANKS & G. ROUHAN. 2007. Spore morphology in relation to phylogeny in the fern genus Elaphoglossum (Dryopteridaceae). Int. J. Plant Sci. 168: 905-929. [ Links ]
23. NAYAR, B. K. 1966. Morphology of the fertile leaves of the Lomariopsidaceae, with special reference to the venation. New Phytol. 65: 221-238. [ Links ]
24. NAYAR, B. K. & S. KAUR. 1965. Spore morphology of the Lomariopsidaceae. J. Palynol. 1: 10-26. [ Links ]
25. PONCE, M. M. 1966. Pteridophyta. In: ZULOAGA, F. & O. MORRONE (eds.), Catálogo de las Plantas Vasculares de la República Argentina I. Monogr. Syst. Bot. Missouri Bot. Gard. 60: 1-79. [ Links ]
26. PONCE, M. M. 2008. Pteridophyta. In: ZULOAGA, F., O. MORRONE & M. BELGRANO (eds.), Catálogo de las Plantas Vasculares del Cono Sur (Argentina, sur de Brasil, Chile, Paraguay y Uruguay). Monogr. Syst. Bot. Missouri Bot. Gard. 107, 3 volúmenes.
27. PRABHAKAR, M. 2003. Structure, delimitation, nomenclature and classification of stomata. Acta Bot. Sin. 46: 242-252.
28. PROCTOR, G. R. 1989. Ferns of Puerto Rico and the Virgin Islands. Mem. New York Bot. Gard. 53: 289-301.
29. ROLLERI, C. & A. DEFERRARI. 1986. Modelos epidérmicos y otros caracteres foliares en la sistemática y ecología de especies de Lycopodium L. Sección Crassistachys Herter. Revista Mus. La Plata, n.s., Bot. 14: 65-87.
30. ROUHAN, G., J. DUBUISSON, F. RAKOTONDRAINIBE, T. J. MOTLEY, J. T. MICKEL, J. LABAT & R. C. MORAN. 2004. Molecular phylogeny of the fern genus Elaphoglossum (Elaphoglossaceae) based on chloroplast non-coding DNA sequences: contributions of species from the Indian Ocean area. Molec. Phylogen. Evol. 33: 745-763.
31. ROUX, J. 1982. The ferns genus Elaphoglossum Schott (Filicales) in South Africa. S. African J. Bot. 48: 481-526.
32. SEHNEM SJ., A. 1979. Aspidiáceas. In: REITZ P. R. (ed.), Flora Ilustrada Catarinense. Vol. 1, pp. 1-91. Herbario Barbosa Rodríguez. Itajaí, Santa Catarina, Brasil.
33. SKOG, J. E., J. T. MICKEL, R.C. MORAN, M. VOLOVSEK & E.A. ZIMMER. 2004. Molecular studies of the New World species in the fern genus Elaphoglossum (Dryopteridaceae) based on chloroplast DNA sequences. Int. J. Plant Sci. 165: 1063-1075.
34. SMITH, A. R., K. M. PRYER, E. SCHUETTPELZ, P. KORALL, H. SCHNEIDER & P. G. WOLF. 2006. A classification for extant ferns. Taxon 55:705-731.
35. TRYON, R & A. TRYON. 1982. Ferns and Allied Plants with special reference to tropical America. Springer-Verlag, New York-Berlín.
36. TRYON, A. F. & B. LUGARDON. 1991. Spores of the Pteridophyta. New York: Springer-Verlag.
Recibido el 9 de marzo de 2010
Aceptado el 14 de junio de 2010

