










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versão On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.45 no.3-4 Córdoba jul./dez. 2010
ARTÍCULO INVITADO
Arquitectura de los árboles
Francis Hallé1
1 Université de Montpellier, Francia.
Email: francis.halle@wanadoo.fr
Conferencia presentada en las XXXII Jornadas Argentinas de Botánica, Huerta Grande, Córdoba, el 6 de Octubre 2009.
Resumen. La forma de la copa nunca es aleatoria; cada árbol tiene su programa específico de desarrollo, controlado por genes, desde la germinación. La forma del árbol adulto puede ser modificada por factores ecológicos, pero siempre persisten las reglas de desarrollo; analizar estas reglas es el objetivo de la arquitectura. Llamaré «modelo arquitectural» al programa genético de crecimiento y desarrollo que aparece en un árbol joven. De los 22 modelos actualmente reconocidos, presentaré los cinco más comunes. Cuando el modelo tiene una expresión única, permaneciendo durante toda la vida, el árbol es «unitario»; estos árboles unitarios son arcaicos. El concepto de «reiteración» es clave para entender lo que es un árbol: este mecanismo transforma el árbol en una colonia; los «árboles reiterados» (AR) crecen los unos encima de los otros, tal como parásitos. La reiteración da a luz un árbol entero, con tronco, ramas y raíces. Un AR todavía joven crece verticalmente; luego, con la plasticidad de la madera y la palanca, se inclina y se vuelve horizontal, lo que es eficaz para capturar la luz. Fue importante el descubrimiento, dentro de una misma copa, de variaciones genéticas de un AR al otro: en varias especies, el árbol es una colonia de genomas.
Palabras clave: Arquitectura de árboles; Modelos arquitecturales; Reiteración.
Summary. Tree architecture. Trees never have a random crown form; each tree has its specific developmental program, controlled by genes, since its germination. The form of the adult tree may be modified by ecological factors but developmental rules are ever lasting; analyzing these rules is the objective of architecture. The genetic program of growth and development of a young tree will be referred to as «architectural model». The five most common out of the 22 architectural models so far recognized will be presented. Whenever a model has a unique expression that persists during the whole life of a tree, the tree is called «unitary»; these unitary trees are ancient. The concept of «reiteration» is key to understand what a tree is: this mechanism turns a tree into a colony; «reiterated trees» (RT) grow on top of each other the way parasites do. Reiteration means the birth of an entire tree, with trunk, branches and roots. A young RT grows vertically; then, with the help of wood plasticity and a lever, it inclines and turns horizontal, thus becoming more efficient in light capturing. It has been a relevant discovery to find, within a single tree crown, genetic variations from one RT to another: in several species, a tree is a colony of genomes.
Key words: Tree architecture; Architectural models; Reiteration.
El estudio de la arquitectura de los árboles se inició en Francia y en Holanda y desde hace 40 años se desarolló en Europa, Estados Unidos, y ahora en China. Desde hace 10 años, aparece una convergencia con la filogenía molecular. La arquitectura y la filogenía moderna dicen lo mismo cuando se trata de dividir una familia de árboles como las Euforbiáceas, o de unir varias familias en una, como Malváceas, Sterculiáceas y Bombacáceas.
En Argentina, se da a conocer a través de un trabajo de Tourn et al. (1999) y actualmente hay un centro de investigación sobre arquitectura vegetal en Bariloche con investigadores formados en Francia como Javier Grossfeld y Javier Puntieri. Pero, la mayoría de los botánicos no utilizan la arquitectura, aunque cada planta tiene su propio programa de desarollo.
Por cierto, la forma tridimensional de un árbol puede ser compleja, pero nunca es aleatoria. Cada árbol tiene su programa específico de crecimiento, controlado por los genes. La forma final puede ser modificada por factores ecológicos pero siempre se quedan las reglas genéticas del desarollo, y existen claves que permiten analizarlas. Indicaré seguidamente tres claves. Enfrente de un árbol, qué se debe mirar?
Primera clave: la dirección de crecimiento de los vástagos, vertical como en un Pino o un Roble, o horizontal como en Albizzia o Annona (Fig. 1). En la mayoría de los árboles, un vástago vertical tiene una filotaxis espiralada, mientras que un eje que crece horizalmente tiene una filotaxis dística (Fig. 2).
En muchas especies, se pueden observar vástagos verticales y horizontales en el mismo árbol. Son ramas y troncos, por ej. En el cafeto o Arbol de Café, Terminalia (manglar) (Fig. 3).
Segunda clave: la disposición de las ramas puede ser continua o rítmica. La ramificación continua, como las ramas en un Arbol de Café, es común en los trópicos. La ramificación rítmica es más común en las regiones templadas, donde se encuentran muchos ejemplos: Cerezo, Laurel, Acebo, Encina, Pino, etc. (Fig. 4).
Tercera y última clave para entender la forma de un árbol: donde están las estructuras reproductivas (flores)? Hay dos posibilidades. La floración puede ser lateral, como en el Almendro o la Higuera, o puede ser terminal como en la Lila o en la Adelfa (Fig. 5). Lo que pasa con la floración terminal es que el vástago no puede crecer más, y tiene que ramificarse. La floración terminal impone una ramificación.
En resumen, ya tenemos tres alternativas:
ejes verticales u horizontales
ramificación continua o rítmica
floración terminal o lateral.
Eso parece muy poco y demasiado simple, pero es un problema de combinación. La combinación de estos rasgos permite definir muchas arquitecturas diferentes o, mejor dicho, muchos modelos de arquitectura. Presentaré cuatro modelos, con ejemplos que se pueden encontrar fácilmente en Argentina.
- El modelo de LEEUWENBERG fué dedicado a un botánico holandés quien utilizó la arquitectura para mejorar la filogenía de las Apocináceas. Todos los vástagos son verticales, y la floración terminal (Fig. 6). Se nota que los ejemplos no son parientes entre ellos; pues un modelo no es un lazo de parentesco, sino que es un nivel de evolución.
- El modelo de RAUH lleva el nombre de un ilustre botánico alemán. Tronco y ramas verticales, ramificación rítmica, y floración lateral. Muchos árboles tienen este programa de desarollo, en el trópicos como en las regiones templadas (Fig. 7). Otra vez, no hay ningún proximidad botánica entre un Pino y un Roble: la arquitectura del árbol aparece como perpendicular a la filogenía.
- El modelo de MASSART, según un botánico de Bélgica quien estudió el magnífico género Araucaria: tronco vertical, ramificación rítmica, y sexualidad lateral. En la Fig. 8 se aprecia un verticilo (o un piso) de ramas, visto desde arriba. Este modelo lo presentan varios árboles, de zonas tropicales y de zonas templadas, perteneciendo tanto a las Gimnospermas como a las Angiospermas.
- El modelo de TROLL ha sido dedicado a un otro ilustre botánico alemán. Cuando todavía joven, cada vástago crece horizontalmente y, después de la caída de las hojas, la base del vástago se vuelve vertical. Por eso, el árbol crece como una sumatoria de dichos vástagos. La floración puede ser terminal como en Annona o lateral como en un Almez (Fig. 9). Este modelo es sumamente común, de lejos, el más común de todos. Puede ser que este modelo sea la cumbre de la evolución arquitectural.
Describimos 22 modelos y presentamos solamente 4. El desafío es encontrar un árbol que no corresponda a ningún de esos 22 (recuerdo que hay 60.000 especies de árboles!) (Fig. 10).
Después del concepto de modelo arquitectural, quería presentar el concepto de reiteración del modelo (o repetición si así lo prefieren). El mecanismo de la reiteración fué descrito por un botánico holandés, Roelof OLDEMAN en 1974. Para entender cómo funciona, lo mejor es mirar a la distancia entre los meristemas que construyen el árbol.
En las raíces y vástagos jovenes, la distancia entre el meristema terminal y los meristemas laterales, es corta, los meristemas están muy próximos (Fig. 11). En este caso de distancia corta -algunos centimetros no más-, los meristemas se relacionan unos con otros, aparece una jerarquía entre ellos y, por eso, funcionan de una manera coordinada. Es decir, que el crecimiento vertical de AA' impone el crecimiento mas horizontal de BB' y CC', e impone también la inhibición completa de los meristemas laterales de los ejes. Así, se construye una arquitectura coordinada, lo que se llama un modelo (Fig. 12).
En árboles de grupos antiguos, Helechos arbóreos, Cycas, Araucaria, Palmeras, Ceiba o Árbol de la nuez de Moscada (Miristicáceas), la jerarquía entre los meristemas nunca se rompe. Durante toda la vida, estos árboles quedan arquitecturalmente unitarios.
Árboles unitarios son pocos. En el caso general, cuando aumenta la distancia entre los meristemas, se rompe la jerarquía y cada uno de los meristemas se vuelve libre (Fig. 13), si se me permite la expresión: Libertad!. La consecuencia es que cada uno de los meristemas funciona como el meristema de una semilla durante la germinación, construyendo un nuevo árbol. Por el mecanismo de la reiteración, un árbol unitario puede volverse en un árbol colonial. Entonces, en este punto, nos encontramos frente a dos categorías de árboles: unitarios y coloniales.
Como ya se dijo, los árboles unitarios (= sin reiteración) son arcaicos. Muchos árboles fósiles fueron unitarios: Psaronicus, Sigillaria o Lepidodendron. En esas épocas geológicas, la reiteración no estaba inventada todavía. En un árbol unitario, el modelo arquitectural tiene una expresión única, que permanece durante toda la vida (Fig. 14). Cualquiera sea su edad, el árbol unitario conserva la misma silueta. Por eso, los árboles unitarios son hermosos y muy utilizados como ornamentales. Pero tienen una limitación: una vida bastante corta, de 2-3 siglos no más.
La reiteración es un progreso, es una forma más moderna y más eficaz de crecer, que se ha generalizada a la mayoría de nuestros árboles. El árbol colonial modifica su silueta durante su vida. Agudo cuando joven, esférico cuando adulto. En el árbol viejo de muchas especies, la copa se vuelve plana, con pocas capas de hojas. Con el tiempo, este tipo de árbol se vuelve cada vez más eficaz para captar la energía luminosa: reiterar es un progreso porque significa aumentar la fotosíntesis, y aumentar también la longevidad del árbol (Fig. 15). En la Fig. 16, se encuentra una vista más detallada del mecanismo de la reiteración, desde el árbol joven hasta el viejo.
Hablemos ahora sobre las hojas de la reiteración. En árboles como Eucalyptus, Acacia, o el Árbol del Pan (Terap), hay hojas juveniles que generalmente desaperecen del árbol adulto. Este follaje juvenil reaparece en la copa del árbol adulto, en la base de cada reiteración (Fig. 17). Eso significa que la reiteración es un árbol entero y completo.
Es importante entender que una reiteración es un árbol pequeño pero entero y completo, con tronco, ramas, hojas y flores. A partir de una yema, este arbolito crece desde el tronco, o encima de una rama, tal como un parásito de tipo muérdago. En castellano se dice «brote» o «chupón», en inglés se llama «sucker» y en francés decimos «gourmand». Todas estas palabras tienen en cuenta el buen apetito del nuevo árbol que crece encima del árbol inicial.
A veces, la biología de la reiteración puede ser extraña. Daré dos ejemplos que están al límite de nuestro conocimiento.
Mi primer ejemplo es la timidez entre árboles vecinos que pertenecen a la misma especie. Eso ocurre en Pináceas, Fagáceas, Mirtáceas y Dipterocarpáceas-Vochysiáceas. Los vecinos nunca se tocan, dejando entre ellos una fisura de timidez (Fig. 18). En una copa muy grande, aparece la timidez entre las reiteraciones, tal como entre árboles de la misma especie.
Un último punto con respecto al genoma. En varios árboles, Tejo del Canadá o Hymenolobium de la Amazonia, hay una variación genética dentro de la misma copa, de una reiteración a la otra: la secuencia de nucleotidos del ácido nucléico no es la misma entre reiteraciones vecinas. Sucede así que durante su vida, un árbol colonial acumula en sí mismo, dentro de una misma copa, esta variación genética y, por eso, se vuelve una colonia de genomas.
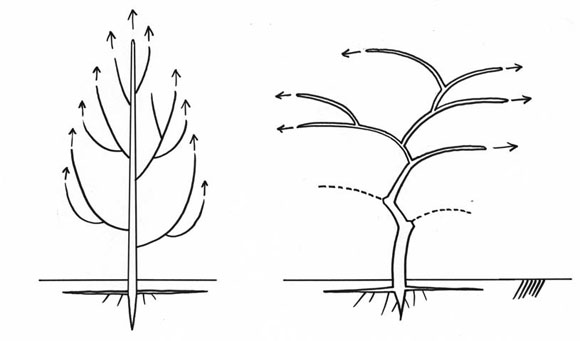
Fig. 1: Dirección de crecimiento de los vástagos. A. vertical; B. horizontal.
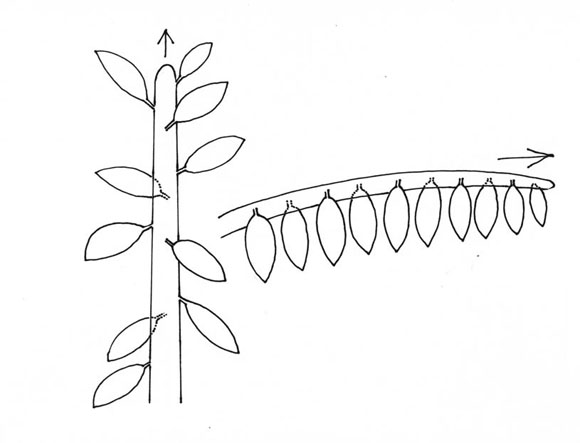
Fig. 2: Filotaxis. A. espiralada en vástagos verticales; B. dística en vástagos horizontales.
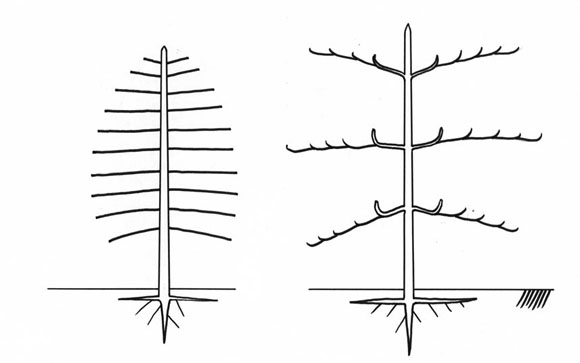
Fig. 3: Combinación de vástagos verticales (tronco) y horizontales (ramas) en el mismo individuo. A. cafeto; B. manglar Terminalia.
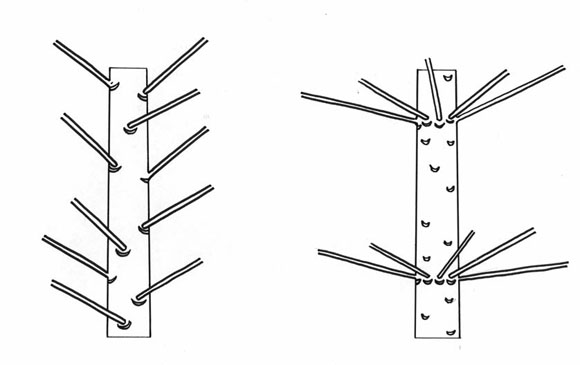
Fig. 4: Tipo de ramificación. A. contínua; B. rítmica.
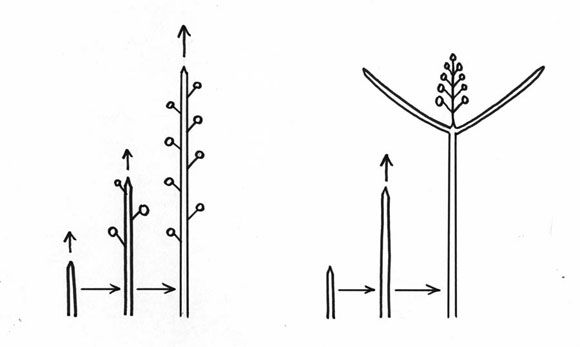
Fig. 5: Posición de las estructuras reproductivas. A. laterales; B. terminales.
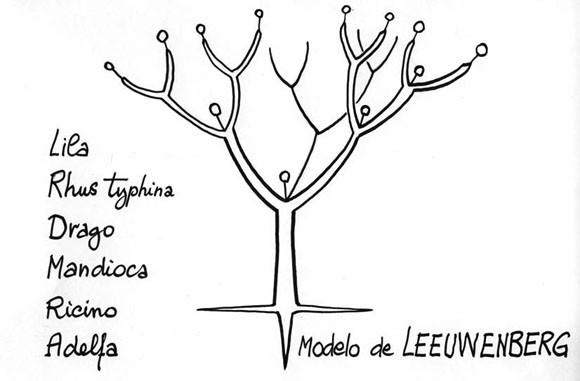
Fig. 6: Modelo de LEEUWENBERG.

Fig. 7: Modelo de RAUH.
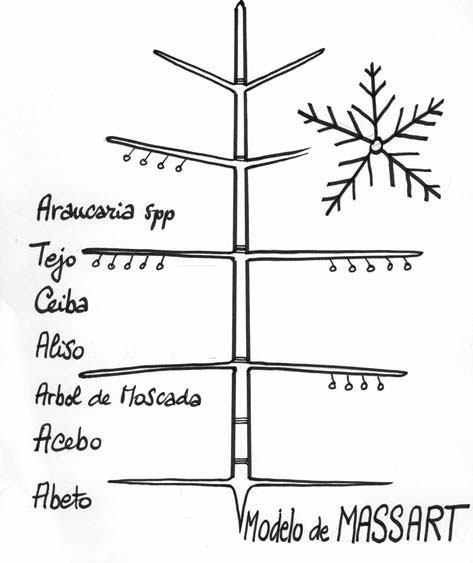
Fig. 8: Modelo de MASSART. Detalle de un verticilo de ramas visto desde arriba.
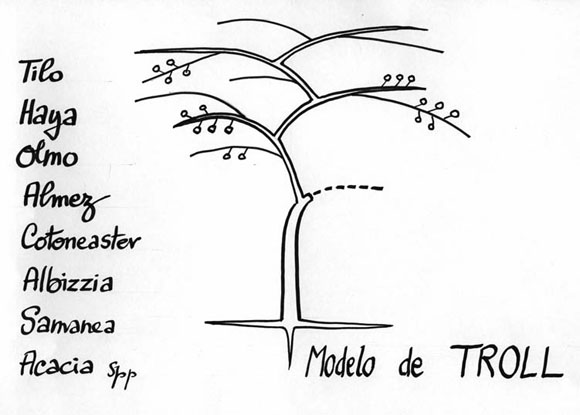
Fig. 9: Modelo de TROLL.
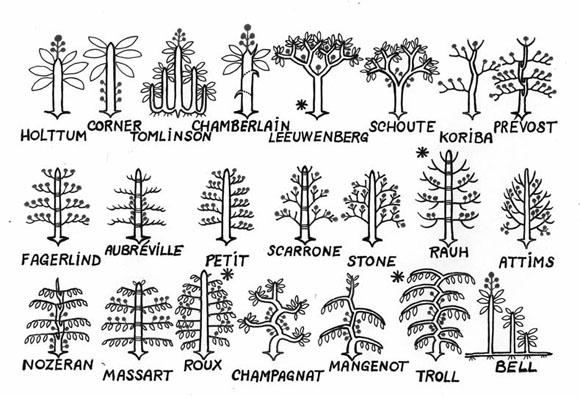
Fig. 10: Los 22 modelos descriptos.
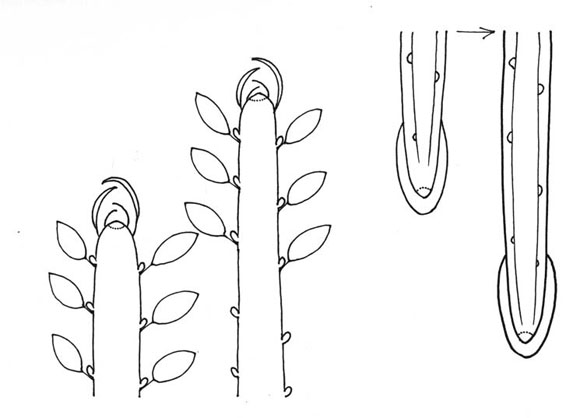
Fig. 11: Distancia entre los meristemas apical y axilares. A. en vástagos; B. en raíces.

Fig. 12: Crecimiento coordinado de los meristemas a distancias cortas. Árboles unitarios.
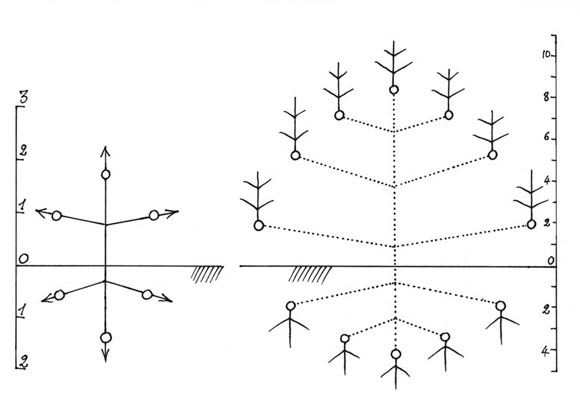
Fig. 13: Diferentes jerarquías en los meristemas, aparecen las reiteraciones. Árboles coloniales.
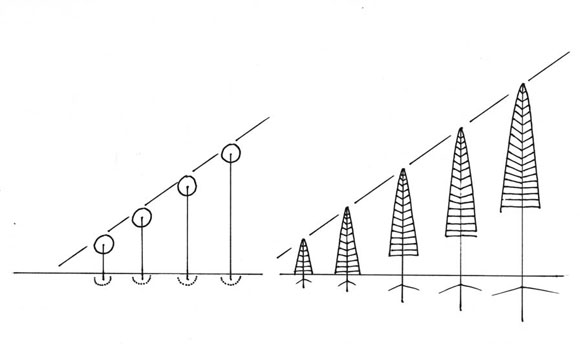
Fig. 14: Modelo arquitectural de árboles unitarios conservan siempre la misma silueta.

Fig. 15: Desarrollo de la copa del árbol por el proceso de reiteración.
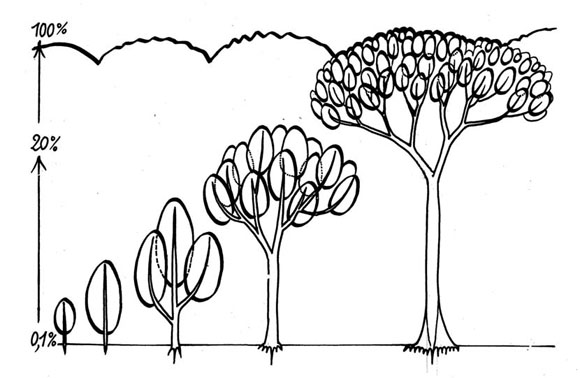
Fig. 16: Vista más detallada del mecanismo de la reiteración, desde el árbol joven hasta el viejo.
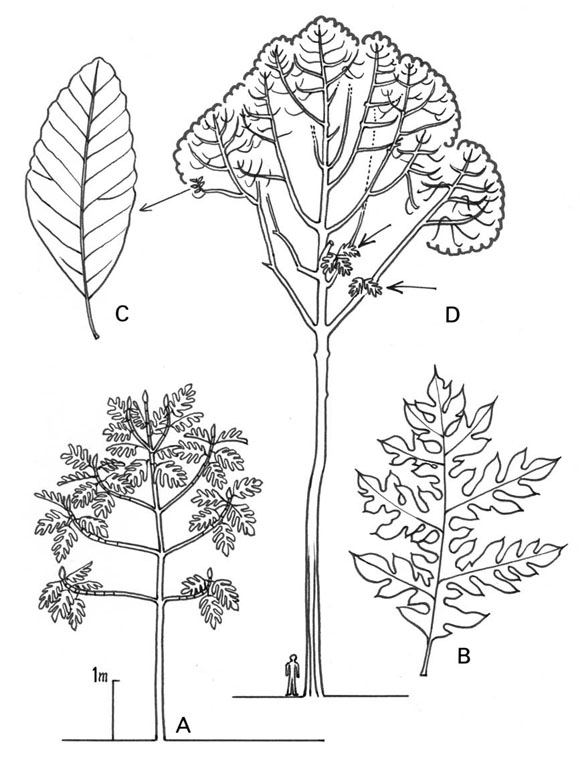
Fig. 17: Reiteraciones en especies con diferentes tipos de hojas: juveniles y adultas. A. árbol joven; B. hojas juveniles; C. hojas adultas en el árbol reiterado; D. hojas juveniles en las reiteraciones.
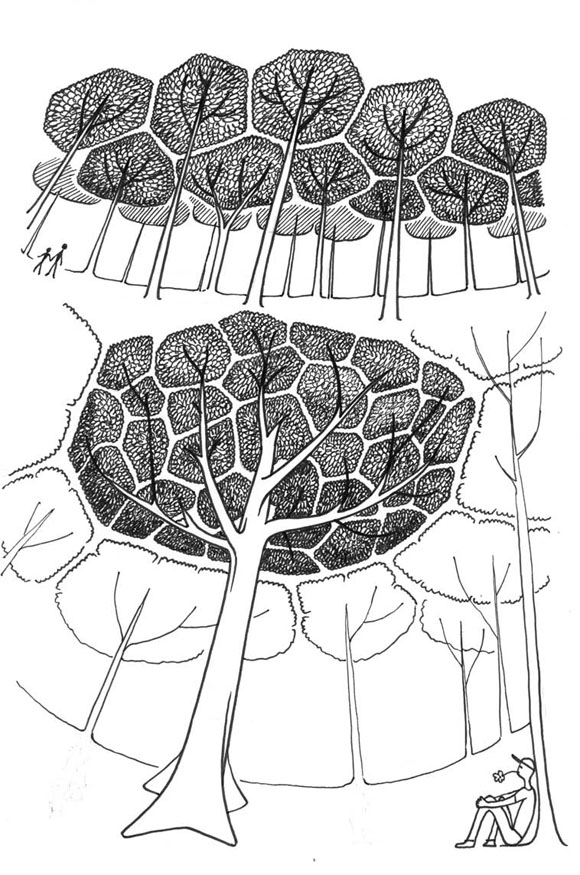
Fig. 18: Fenómeno de timidez entre árboles de la misma especie.
BIBLIOGRAFÍA
1. BARTHÉLÉMY, D., C. EDELIN & F. HALLÉ. 1989. Architectural concepts for tropical trees. In: Holm-Nielsen, L. B. & H. Balslev (eds.), Tropical forests: botanical dynamics, speciation and diversity, pp. 89-100. Academic Press, London. [ Links ]
2. BELL, A. D. 1991. Plant Form: An Illustrated Guide to Flowering Plant Morphology. Oxford University Press, Oxford. [ Links ]
3. HALLÉ, F. & R. A. A. OLDEMAN. 1970. Essai sur l'architecture et la dynamique de croissance des arbres tropicaux. Masson Ed., Paris. [ Links ]
4. HALLÉ, F., R. A. A. OLDEMAN & P. B. TOMLINSON. 1978. Tropical trees and forests. Springer, Berlin. [ Links ]
5. TOURN, G. M., G. BARTHÉLÉMY & J. GROSSFELD. 1999. Una aproximación a la arquitectura vegetal. Bol. Soc. Argent. Bot. 34: 85-99. [ Links ]
Recibido el 5 de abril de 2010
Aceptado el 4 de junio de 2010.

