










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.46 no.3-4 Córdoba jul./dic. 2011
ANATOMÍA Y MORFOLOGÍA VEGETAL
Alteraciones morfo-histológicas en hojas de Solanum chenopodioides (Solanaceae), producidas por ácaros y dípteros minadores
Silvana D. Del V. Figueroa, Nilda Dottori y María Teresa Cosa1
1 Laboratorio de Morfología Vegetal, Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de Córdoba, e Instituto Multidisciplinario de Biología Vegetal (I.M.B.I.V.). C.C. 495. 5000 Córdoba. Argentina. silfigueroa2003@yahoo.com.ar; dottori@imbiv.unc.edu.ar; cosa@imbiv.unc.edu.ar
Resumen: Solanum chenopodioides Lam., conocida como "hierba mora", es una hierba o subarbusto ampliamente distribuido en Argentina y otras zonas de Sudamérica y es una maleza sumamente tóxica para el ganado. Sus hojas tienen propiedades medicinales y son utilizadas como sedante y en oftalmología. Es frecuente el ataque de sus hojas por ácaros fitófagos y por insectos minadores. El objetivo del presente trabajo fue analizar la morfología y la anatomía de las hojas en plantas sanas e infestadas por ácaros (Tetranychidae, Tetranychus sp.) y dípteros minadores de la hoja (Agromyzidae). Los resultados muestran que el ataque de ambos agentes reduce significativamente el volumen del tejido fotosintético por excelencia. Así, se compromete el crecimiento, y por lo tanto la supervivencia de la especie. Aún más, en caso de ser utilizada como especie medicinal se afecta la calidad del principio activo.
Palabras clave: Solanum chenopodioides; Hojas; Acaros fitófagos; Dípteros minadores de hoja.
Summary: Morpho-histological alterations in leaves of Solanum chenopodioides (Solanaceae) produced by mites and leaf miner diptera. Solanum chenopodioides Lam., known as "nightshade", is a herb or subshrub widely distributed in Argentina and another zones of South America, and is a weed highly toxic to livestock. Its leaves have medicinal properties and are used as a sedative and in ophthalmology. The attack of its leaves by phytophagous mites and leafminer insects is frequent. The aim of this study was to analyze the morphology and the anatomy of the leaves of both healthy and infested plants by Tetranychidae, Tetranychus sp. mites and Agromyzidae leafminer diptera. The results show that the attack of both agents significantly reduces the amount of photosynthetic tissue par excellence. Thus, growth is compromised, and therefore the survival of the species. Moreover, when used as medicinal species, the quality the active principle is affected.
Key words: Solanum chenopodioides; Leaves; Phytophagous mites; Leafminer diptera.
Introducción
Solanum chenopodioides Lam. es conocida con el nombre vulgar de "hierba mora", pertenece al subgénero Solanum sect. Solanum Nee, (1999). La especie en estudio integra una familia de reconocida importancia económica, comprende especies de valor alimenticio, otras que contienen compuestos químicos de interés en medicina y farmacología muchos de ellos tóxicos (Hawkes, 1999; Hunziker, 2001) y otras son ornamentales (Sérsic et al., 2006).
De amplia dispersión en Sudamérica crece en Brasil, Chile, Bolivia, Paraguay, Uruguay y Argentina (Marzocca, 1997). En nuestro país, habita desde Jujuy hasta Río Negro; en Córdoba es frecuente en los departamentos Capital, Colón, Juárez Celman, Punilla, Río Cuarto, Río Segundo, San Alberto, San Justo, Santa María y Totoral (Barboza et al., 2006).
Es una maleza de los cultivos de algodón, trigo, café, leguminosas de grano, etc., crece a orillas de cercos, rara vez en alfalfares, huertas, montes frutales; también puede ser hallada en campos abandonados; prefiere terrenos húmedos y sombríos. Su presencia en las pasturas es inconveniente, pues se trata de una especie sumamente tóxica para el ganado (bovinos, ovinos, caprinos y porcinos), debido a la presencia de los alcaloides solanidina y solasodina (Marzocca, 1997).
La parte aérea tiene propiedades medicinales utilizándose como sedante, hepática, narcótica, anticefálica, en oftalmología, etc. Sus hojas cocidas son comestibles y también son utilizadas como cataplasmas (Marzocca, 1997; Barboza et al., 2006).
Se han realizado en esta especie estudios cariológicos (Moscone, 1992; Acosta et al., 2005), de anatomía y desarrollo del fruto y semilla, de germinación y éxito reproductivo (Cabrera et al., 2008; 2010) y de anatomía de órganos vegetativos (Figueroa et al., 2008).
En relación a los caracteres exomorfológicos, S. chenopodioides es una hierba o subarbusto de 50 a 80 cm de altura, anual o bienal. Posee hojas pecioladas, pilosas, ovado-lanceoladas, con la base cuneada o redondeada, miden de 4,6-6,4 cm de longitud. por 2,1-2,6 cm. Las inflorescencias son umbeliformes con flores blancas dispuestas en cimas y el fruto es una baya globosa negro-azulada (Marzocca, 1997; Barboza et al., 2006.)
Es importante señalar que los autores han detectado que las hojas de esta especie son atacadas con frecuencia por ácaros fitófagos (Tetranychus sp.), conocidos vulgarmente como "arañas rojas y pardas" y que además la especie es hospedante de dípteros "minadores de hoja" (Agromyzidae). Aún más, hay antecedentes en relación a la infestación de plantas hortícolas de Solanáceas como Solanum melongena L. (berenjena), S. lycopersicum L. (tomate), entre otras especies, con Tetranychus urticae Koch (Badii et al., 2010) también en Phaseolus vulgaris L. (Aponte & Aponte, 1990), y en Prunus persica (Doreste, 1978). Además se conoce la presencia de minadores de hojas en especies de Solanaceae como S. melongena (berenjena ), S. tuberosum L. (papa), S. lycopersicum (tomate), Capsicum sp. (Barraco Vega, 2003) y en Commelina diffusa Burm. f. y Floscopa glabata (Kunth) Hassk de la familia Commelinaceae, (Elb et al., 2010).
El principal daño producido por los Agromícidos es debido a las larvas, también las hembras pueden producir daños importantes debido a sus hábitos alimenticios y de oviposición, ellas practican pequeñas perforaciones en las hojas y pétalos de flores, de modo que si la densidad de hembras es muy elevada pueden llegar a producir la destrucción de dichas estructuras (Barraco Vega, 2003). Los daños producidos por los ácaros son más importantes en las plantas jóvenes, provocando un retraso en su crecimiento y disminución de la producción de frutos y calidad de los mismos. En casos extremos de grandes poblaciones de "arañas rojas", pueden llegar a secar la planta (Aponte & Aponte, 1990).
Teniendo en cuenta lo anteriormente expuesto y la importancia de la especie, el objetivo del presente trabajo fue analizar la morfología y anatomía de las hojas en plantas sanas y atacadas por ácaros y por dípteros minadores a fin de conocer las alteraciones morfo-histológicas que provocan.
Materiales y Métodos
Los materiales estudiados de S. chenopodioides se encuentran depositados en el herbario del Museo Botánico de Córdoba, Argentina (CORD).
Prov. Córdoba: Dpto. Capital: Isla de los patos-costanera, Figueroa 1, 07-I-2006, 11-I-2006 y 18- III-2006; Barrio Cerro de las Rosas, Rafael Núñez, Figueroa 2, 29-IV-2007; Barrio Alta Córdoba, Dottori 214, 21-XII-2004. Dottori 220, 29-III-2007. Dottori 222, 27-IV-2010.
Los preparados permanentes por láminas de hojas sanas2, atacadas por ácaros3 y dípteros minadores4 se realizaron con materiales previamente fijados en FAA.
Para esto el material fue sometido a una bomba de vacío a fin de extraer el aire, deshidratado en una serie de alcohol etílico y xilol, e incluido en Histoplast (Conn et al., 1960). Luego se realizaron cortes transversales con micrótomo de 10 a 20 mm de espesor, que fueron coloreados con Azul Astral-Fucsina Básica (Kraus et al., 1998). Las observaciones se realizaron con microscopio óptico y las fotomicrografías se tomaron con cámara digital.
Resultados
Exomorfología:
Las hojas atacadas exhiben externamente los daños producidos por los estiletes de ácaros fitófagos y la reabsorción del contenido celular para su alimentación. El síntoma más característico es la aparición de puntos o manchas amarillentas en la cara adaxial, debido a la ausencia de clorofila en el mesofilo. El envés de las hojas, pueden presentar "arañas rojas" en todos sus estadios y su telaraña. (Fig. 1 A, B y C).

Fig. 1: Hoja en vista superficial atacada por ácaros fitófagos. A: cara adaxial mostrando zonas claras donde el mesofilo ha sido consumido. B y C: detalle donde se observan ácaros fitófagos (a) y telas (te).
Externamente se observan las minas producidas por dípteros, pardas o amarillentas por carecer de cloroplastos (Fig. 2 A, B, C y D). En la especie estudiada se observaron minas lineares y otras donde las lineares presentan variaciones ya que las galerías se unen irregularmente.

Fig. 2: Hojas en vista superficial atacadas por dípteros minadores. A: cara adaxial mostrando pequeñas perforaciones realizadas por la hembra con el ovipositor. B: detalle de A. C y D: galerías (ga) producidas por larvas de los dípteros minadores.
Anatomía de la hoja:
Las hojas sanas, en transcorte presentan la epidermis unistratificada, de cutícula delgada, con células comprimidas; a la altura del hacecillo central éstas son redondeadas y altas (Fig. 3 A y B).
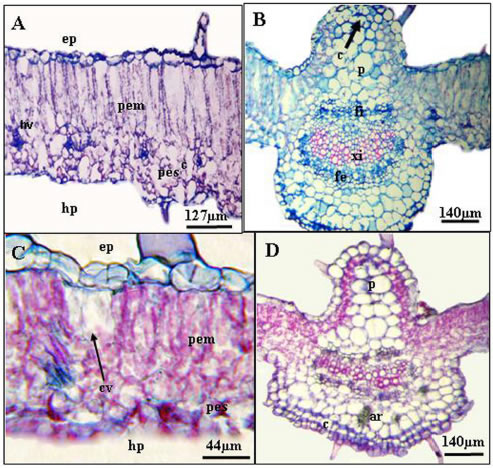
Fig. 3: Anatomía de la hoja en corte transversal. A y B: cortes por la lámina y a nivel de la vena media de hojas sanas, respectivamente. C: corte por la lámina donde se observan células vacías del parénquima en empalizada. D: corte a nivel de la vena media en hojas atacada por ácaros fitófagos, donde se observan variaciones en el número y tamaño de las células parénquimáticas en ambas caras. Abreviaturas. ar: areniscas cristalinas; c: colénquima; cv: células vacías; ep: epifilo; fe: floema externo; fi: floema interno; hp: hipofilo; hv: hacecillo vascular; p: parénquima; pem: parénquima en empalizada; pes: parénquima esponjoso; xi: xilema.
El mesofilo, de estructura dorsiventral, posee 1 ó 2 estratos de clorénquima en empalizada hacia la cara adaxial y 5 ó 6 de clorénquima esponjoso hacia la cara abaxial, con grandes espacios intercelulares (Fig. 3 A).
La zona de la vena media sobresale en el haz y el envés de la lámina y se diferencian 5 capas de colénquima angular subepidérmico hacia la cara adaxial y 1 capa subepidérmica hacia la cara abaxial; además, hay de 5 a 6 estratos de parénquima hacia ambas caras del hacecillo bicolateral (Fig. 3 B).
El haz vascular bicolateral está rodeado por una vaina, no bien definida de células parenquimáticas. El floema externo tiene desarrollo uniforme y el floema interno se dispone en grupos separados por células parenquimáticas (Fig. 3 B).
En las hojas atacadas por ácaros la lámina presenta igual disposición de tejidos que en la hoja sana, sin embargo, se observan en el parénquima en empalizada, células sin su contenido citoplasmático (Fig. 3 C). Además, varía la forma del haz principal debido a cambios en el número y tamaño de las células parenquimáticas hacia ambas caras (Fig. 3 D).
Las alteraciones en la anatomía de las hojas atacadas por dípteros minadores, consisten en la destrucción del parénquima asimilador por el aparato bucal masticador de las larvas. De este modo se forman galerías o túneles lineares. La epidermis queda intacta, protegiendo a las larvas; además, se detecta un tejido de reacción en el parénquima, cuando las hembras perforan la epidermis para depositar sus huevos (Fig. 4 A, B y C).

Fig. 4: Anatomía de hojas en corte transversal atacadas por minadores. A: lámina mostrando galerías. B: detalle de una amplia galería donde el mesofilo ha sido totalmente consumido por la larva, quedando sólo ambas epidermis. C: detalle del tejido de reacción producido por el ovipositor. Abreviaturas. ep: epifilo; gal: galería; hp: hipofilo; pem: parénquima en empalizada; pes: parénquima esponjoso; tejr: tejido de reacción.
Discusión y Conclusiones
La estructura dorsiventral de la hoja en S. chenopodioides, característica de especies mesofíticas, concuerda con la descripción realizada para la misma especie por Figueroa et al., (2008) y permite explicar su preferencia por terrenos húmedos tal como lo indica para esta especie Marzocca (1997).
Los daños producidos por los ácaros son más importantes en las plantas jóvenes, provocando un retraso en su crecimiento. En casos extremos de grandes poblaciones de "arañas rojas", pueden llegar a secar la planta esto también fue observado por Aponte & Aponte, (1990) en P. vulgaris.
Tetranychus urticae ataca cultivos de plantas hortícolas de Solanáceas como "berenjena", "tomate" (Badii et al., 2010), Phaseolus vulgaris L. "poroto" (Aponte & Aponte, 1990), los daños son similares al causado por la mayoría de los ácaros; se observan células vacías ya que es removido el contenido celular del tejido afectado. Aparecen manchas blanquecinas que aumentan de tamaño y zonas cloróticas; además, sedas e hilos entre las hojas esto coincide con lo observado en S. chenopodiodes mientras que en Prunus persica L. "duraznero" se observaron daños semejantes en el envés de la hoja cuyos bordes se doblan longitudinalmente hacia arriba y luego cuando la invasión es grande se extienden a toda la hoja (Doreste, 1978). En cuanto a los daños provocados por dípteros minadores de hoja Agromyzidae, estudios realizados han permitido conocer su presencia en especies de Solanaceae como S. melongena, S. tuberosum L., S. lycopersicum, Capsicum sp. (Barraco Vega, 2003) y en C. diffusa y Floscopa glabata, (Elb et al., 2010) y detectar la presencia de minas como ocurre en S. chenopodiodes. Las minas pueden clasificarse según su forma en dos tipos principales: linear o formando amplias cámaras (Hering, 1951). Los estudios de Elb et al., (2010) en Commelinaceae han permitido reconocer minas de forma linear y de serpentina, mientras que en S. chenopodioides son en su mayoría lineares y otras presentan variaciones donde las lineares se unen irregularmente.
Según Salvo & Valladares (2007), el daño que producen los insectos minadores en plantas de importancia hortícola u ornamental puede ser controlado por parasitoides; insectos cuyas larvas se alimentan de otros insectos como son los minadores. Este mecanismo de control biológico, no fue advertido en esta especie, que interesa por sus propiedades medicinales, si bien es posible que ocurra.
Los resultados muestran que el ataque de ambos agentes patógenos reduce significativamente el volumen del tejido fotosintético por excelencia. Así se ve comprometido el crecimiento, y por lo tanto la supervivencia de la especie. Aún más, en caso de ser utilizada como medicinal se afecta la calidad del principio activo.
Agradecimientos
Agradecemos al Doctor Miguel Delfino por confirmar la identificación de los ácaros fitófagos, a las Doctoras Graciela Valladares y Adriana Salvo por su ayuda en la identificación de los dípteros minadores y aporte de bibliografía. A la Secretaría de Ciencia y Tecnología, de la Universidad Nacional de Córdoba (SECyT), por el apoyo económico brindado.
2 Plantas sanas ( Isla de los Patos, Cerro de las Rosas, Alta Córdoba, Dottori 214);
3 Plantas atacadas por ácaros (Alta Córdoba, Dottori 220).
4 Plantas atacadas por dípteros minadores (Alta Córdoba, Dottori 222)
Bibliografía
1. ACOSTA, M. C., G. BERNARDELLO, M. GUERRA & E. A. MOSCONE. 2005. Karyotype analysis in several South American species of Solanum and Lycianthes rantonnei (Solanaceae). Taxon 54: 713- 723. [ Links ]
2. APONTE, O. R. & F. APONTE.1990. Daño de Tetranychus urticae Koch y su influencia en el cultivo de la caraota (Phaseolus vulgaris L.). Bol. Entomol. Venez. N.S. 5: 197-204. [ Links ]
3. BARBOZA, G., J. CANTERO, C. NÚÑEZ & L. ARIZA ESPINAR. 2006. Flora medicinal de la Provincia de Córdoba (Argentina) Pteridófitas y Antófitas silvestres o naturalizadas. Museo Botánico Córdoba. [ Links ]
4. BADII, M. H., J. LANDEROS & E. CERNA. 2010. Regulación poblacional de ácaros plaga de Impacto Agrícola. Daena Int. J. of Good Conscienc. 5: 270- 302. [ Links ]
5. BARRACO VEGA, P. 2003. Dípteros de interés agronómico. Agromícidos plaga de cultivos hortícolas intensivos. Bol. S.E.A. 33: 293-307. [ Links ]
6. CABRERA, V. A.; N. DOTTORI & M. T. COSA. 2008. Anatomía y desarrollo de fruto y semilla en Solanum chenopodiodes Lam. (Solanaceae). Arnaldoa 15: 237-246. [ Links ]
7. CABRERA, V. A.; N. DOTTORI & M. T. COSA. 2010. Germinación, Éxito reproductivo y Fenología de Solanum chenopodiodes (Solanaceae). Bol. Soc. Argent. Bot. 45: 73-80. [ Links ]
8. CONN, H. J., M. A. DARROW & V. M. EMMEL. 1960. Staining procedures. Ed. Williams and Wilkins Co., Baltimore. [ Links ]
9. DORESTE, S. E. 1978. Dos ácaros plagas del duraznero Prunus persica en Venezuela. Agron. Trop. 10: 121-127. [ Links ]
10. ELB, P. M., G. F. MELO DE PINNA & N. LUIZA DE MENESES. 2010. Morphology and anatomy of leaf miners in two species of Commelinaceae (Commelina diffusa Burm. f. and Floscopa glabata (Kunth) Hassk). Acta Bot. Bras. 34: 283-287. [ Links ]
11. FIGUEROA, S. D., N. DOTTORI & M. T. COSA. 2008. Anatomía de órganos vegetativos en Solanum chenopodiodes (Solanaceaea). Arnaldoa 15: 247- 254. [ Links ]
12. HAWKES, J. G. 1999. The economic importance of the family Solanaceae. In: M. NEE, D.E SYMON, R.N. LESTER & J.P. JEPSON (eds.). Solanaceae IV. Advances in biology and utilization. Royal Botanical Gardens, Kew. [ Links ]
13. HERING, E. M. 1951. Biology of the leaf miners. Dr. W. Junk. The Hague. The Netherlands. [ Links ]
14. HUNZIKER, A. T. 2001. Genera Solanacearum. The genera of Solanaceae illustrated, arranged according to a new system. A. R. G. Gantner Verlag, K. G. Ruggell. [ Links ]
15. KRAUS, J. E., H. DE SOUSA, M. E. REZENDE, N. M. CASTRO, C. VECCHI & R. LUQUE. 1998. Astra Blue and Basic Fuchsin double staining of plant materials. Biotech. Histochem. 73: 235-243. [ Links ]
16. MARZOCCA, A. 1997. Vademécum de Malezas Medicinales de la Argentina. I-XIX, P. 385. Orientación Gráfica Editora. Buenos Aires. [ Links ]
17. MOSCONE, E. A. 1992. Estudios de cromosomas meióticos en Solanum de Argentina. Darwiniana 31: 261-297. [ Links ]
18. NEE, M. 1999. Synopsis of Solanum in the New World. In: M. NEE, D. E. SYMON, R. N. LESTER & J.P. JESSOP (eds.). Solanaceae IV: 285-333. Royal Botanic Gardens, Kew. [ Links ]
19. SALVO, A. & G. R. VALLADARES. 2007. Parasitoides de minadores de hojas y manejo de plagas. Cien. Inv. Agr. 34 (3): 167-185. [ Links ]
20. SÉRSIC, A., A. COCUCCI, S. BENÍTEZ-VIEYRA, A. COSACOV, L. DÍAZ, E. GLINOS, N. GROSSO, C. LAZARTE, M. MEDINA, M. MORÉ, M. MOYANO, J. NATTERO, V. PAIARO, C. TRUJILLO & P. WIEMER. 2006. Flores del Centro de Argentina. Una Guía Ilustrada para conocer 141 especies típicas. Academia Nacional de Ciencias de Córdoba, Argentina. 354 pp. [ Links ]
Recibido el 26 de mayo de 2011,
aceptado el 4 de octubre de 2011.

