










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.47 no.3-4 Córdoba jul./dic. 2012
FICOLOGÍA
Variaciones temporales del fitoplancton de los ríos tributarios y emisario del embalse C. Gelsi (Tucumán, Argentina)
Silvia N. Martinez de Marco1,2 y Beatriz C. Tracanna1,3
1 Instituto de Ficología, Fundación Miguel Lillo, Miguel Lillo 251, C.P. 4000 - San Miguel de Tucumán, Argentina. Email: silmdemar@gmail.com;
2 Instituto de Limnología del NOA, Facultad de Ciencias Naturales e IML. UNT;
3 CONICET.
Resumen: En este trabajo se estudiaron las variaciones temporales del fitoplancton de los tributarios (ríos Salí, Tapia y Vipos) y del emisario (río Salí) del embalse C. Gelsi. La abundancia algal mínima se obtuvo en verano con una biomasa de 0,19-0,58 µg Cl-a L-1 y la máxima en invierno o primavera con una biomasa de 5,43-6,79 µg Cl-a L-1. En los tributarios se destacaron las diatomeas pennadas sobresaliendo Diatoma vulgare y Cymbella affinis y en el río Salí (emisario) dominaron las céntricas especialmente Aulacoseira granulata y Cyclotella meneghiniana. Las Chlorophyta se ubicaron en segundo término en el río Salí, las Cyanobacteria codominaron en los ríos Vipos y Tapia en marzo de 1998 y las Dinophyta sólo fueron observadas en el Salí (emisario). El índice de diversidad específica osciló entre 0,2 y 4,2 y el sapróbico caracterizó las aguas como "no poluídas" o "ligeramente poluídas" en invierno y como "moderadamente poluídas" en verano. El NMDS ordenó temporalmente las muestras según la abundancia de las especies algales consideradas. Los dos primeros ejes del RDA separaron los sitios según gradientes de temperatura, pH, conductividad, DBO5 y nitrato, además de la abundancia de Aulacoseira granulata, Cyclotella meneghiniana, Ulnaria ulna, Cymbella affinis y Leptolyngbya foveolarum.
Palabras clave: Fitoplancton; Densidad algal; Biomasa; Índices de diversidad y sapróbico; Tributarios y emisario; Embalse C. Gelsi.
Summary: Temporal variations of the phytoplankton of the tributaries and effluent from the C. Gelsi reservoir (Tucumán, Argentina). In this paper we analyzed the phytoplankton variation in the tributaries (Salí, Tapia, and Vipos rivers) and effluent (Salí River) from the C. Gelsi reservoir according to a seasonal gradient. The minimum algal abundance took place in summer with a biomass of 0.19-0.58 µg Cl-a L-1, while the maximum occurred in winter or in spring, with a biomass of 5.43-6.79 µg Cl-a L-1. In the tributaries pennate diatoms prevailed, Diatoma vulgare and Cymbella affinis were most abundant, while in the Salí river (Gelsi effluent) centric diatoms were dominant, particularly Aulacoseira granulata and Cyclotella meneghiniana. Chlorophyta were ranked second in the Salí River, Cyanobacteria codominated in the Vipos and Tapia rivers in March 1998, and Dinophyta could only be seen in the Salí River (Gelsi effluent). The specific diversity index ranged from 0.2 to 4.2 and the saprobic index characterized waters as "non-polluted" or "slightly polluted" in winter, and as "moderately polluted" in summer. The NMDS arranged temporarily the samples according to the abundance of the algal species considered. The RDA showed that the two first axes separated the samples according to gradients of temperature, pH, conductivity, BOD5 and nitrate, and also abundance of Aulacoseira granulata, Cyclotella meneghiniana, Ulnaria ulna, Cymbella affinis and Leptolyngbya foveolarum.
Key words: Phytoplankton; Algal density; Biomass; Diversity index and saprobic index; Tributaries and effuent; C. Gelsi reservoir.
INTRODUCCIÓN
La provincia de Tucumán posee una densa red hidrográfca, que abarca la mayor parte de su territorio, y está representada principalmente por la cuenca del río Salí-Dulce (Fernández et al., 2007). Todo el caudal que drena de los faldeos orientales del sistema del Aconquija y de las Cumbres Calchaquíes es captado por el río Salí que es el colector más importante de la cuenca y eje del desarrollo urbano e industrial de la provincia (García et al., 2007). Esta extensa red hídrica constituye el soporte de toda la actividad socioeconómica de Tucumán. Sus aguas se utilizan para consumo, riego, uso industrial y además constituyen el medio de evacuación de muchos de los efluentes que generan estas actividades (Fernández et al., 2007).
Los ambientes lóticos analizados forman parte de la cuenca superior del río Salí que corresponde al sector que se extiende desde sus nacientes hasta su paso por el Cajón del Cadillal. El área de estudio se encuentra ubicada en el Valle de Tapia-Trancas, que es una depresión de origen tectónico, en el centro-norte de la provincia de Tucumán, limitada al oeste por las Cumbres Calchaquíes y al este por las Sierras del noreste tucumano (Páez, 2007). Esta subregión agroecológica tiene limitaciones climáticas y edáficas, con déficit hídrico permanente y suelos de textura liviana con escasa retención de agua. El régimen de precipitaciones con lluvias estivales e inviernos secos condiciona la producción, haciéndose imprescindible el riego como factor determinante de ésta. La principal actividad económica es la ganadería orientada a la producción lechera, el cultivo de forrajes y la producción de hortalizas y granos. En la considerable superficie territorial que ocupa la cuenca de Tapia, la mayor parte de la población, el 71% habita el espacio rural y se distribuye en forma dispersa por toda la geografía departamental tranqueña (Garrido, 2005).
El estudio del embalse C. Gelsi, el más grande e importante de Tucumán, que ya fuera abordado en otras contribuciones, fue creado en 1966 para la atenuación de crecidas, provisión de agua potable para la capital de Tucumán, además de riego, generación de energía eléctrica, recreación y turismo (e.j. Tracanna et al., 1991, 2006; Villagra et al., 1993; Martínez De Marco et al., 2007, 2011). En este trabajo se aborda, por primera vez, el estudio del fitoplancton considerando densidad, diversidad específica, biomasa e índice sapróbico de los afuentes del embalse (ríos Salí (tributario), Tapia, Vipos) y también de su emisario, el río Salí, a lo largo de un gradiente temporal. Asimismo, se analizó la relación entre las variables físicas, químicas y bióticas mediante la aplicación de métodos multivariados de ordenación (NMDS y RDA). Los resultados representan un aporte al conocimiento de la ecología de estos ambientes lóticos que tienen relación directa con la provisión de agua para potabilizar.
MATERIAL Y MÉTODO
Los muestreos se efectuaron mensualmente por el término de 13 meses desde marzo de 1998 a marzo de 1999. Los sitios de muestreo seleccionados (Fig. 1) fueron: 1.- río Salí (tributario): en la estación de aforo San Vicente (26º 28' LS; 65º 12' LO; altitud: 654 m snm); 2.- río Vipos: en intersección con la ruta nacional Nº 9, km 1336, (26º 28' LS; 65º 18' LO; altitud: 741 m snm); 3.- río Tapia: en intersección con la ruta provincial Nº 348 (26º 36' LS; 65º 14' LO, altitud: 635 m snm); 4.- río Salí (emisario): aproximadamente a 200 metros aguas abajo de la Usina Hidroeléctrica (26º 37' LS; 65º 11' LO; altitud: 557 m snm). Los datos de ubicación de cada sitio se obtuvieron mediante Posicionador Satelital Global (GPS 12 XL Garmin).
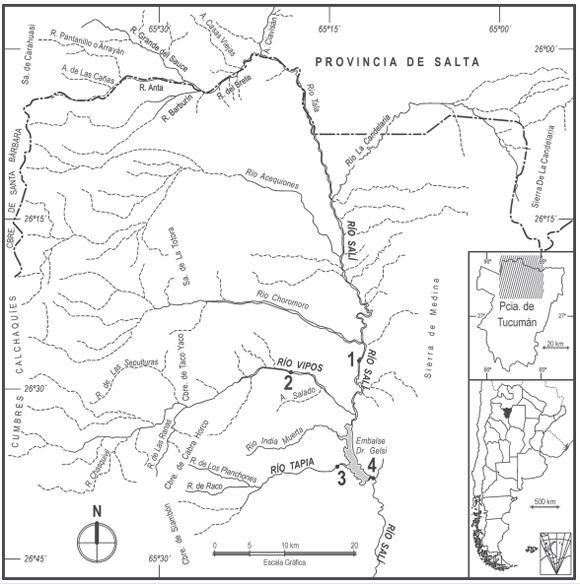
Fig. 1. Mapa de la red hidrográfica de la cuenca superior del río Salí (Cuenca Tapia-Trancas) y ubicación de los sitios de muestreo dentro del área de estudio: 1.- río Salí (tributario), 2.- río Vipos, 3.- río Tapia, 4.- río Salí (emisario).
Las muestras para el estudio cuantitativo del fitoplancton, se obtuvieron directamente con botellas de 500 cm3 y fueron fijadas "in situ". Los recuentos se realizaron bajo microscopio invertido Zeiss IDO2 a un aumento de 400 X. Se utilizaron columnas de diferentes volúmenes (50, 10 y 2 ml) de acuerdo al aumento de la turbidez de las muestras. En algunos casos se hicieron diluciones debido a la excesiva cantidad de sedimentos por las crecidas de los ríos a causa de las lluvias estivales. Se contaron 300 campos elegidos al azar, aceptándose un error máximo del 20 %. Para el cálculo del error se tuvo en cuenta la cantidad de campos contados por cámara en relación al total de campos (N = 3.448), con una confianza del 95 % (α = 0,05). Se expresó la abundancia algal de las especies de cada sistema lótico de acuerdo a los siguientes intervalos o rangos (ind mL-1): (*): 1-2; (+): 2-4; (□): 4-8; (x): 8-16; (◊): 16-32; (■): 32-64; (●): 64-128; (‡): 128-256; (▲): 256-512; (O): >512 (Patrick, 1963).
Para cuantificar biomasa algal (en términos de μg de clorofila a L-1 ) se aplicó el método espectrofotométrico monocromático (Loez, 1995). Se filtraron 5 litros de agua con filtros de fibra de vidrio Whatman GF/C de 1 µm de poro y se usó metanol como solvente. Para el cálculo de la cantidad de clorofila a (expresada en µg L-1) se aplicó la fórmula de Talling & Driver (Ros, 1979).
En el cálculo de la biodiversidad específica del fitoplancton se empleó la fórmula de Shannon & Weaver (1963).
Para la evaluación de la calidad del agua en relación con el fitoplancton se empleó el índice sapróbico (IS) de Pantle & Buck (1955) a través de la fórmula: IS = ∑(s . h) / ∑ h, donde h corresponde a la densidad algal y s al grado sapróbico estimado de acuerdo con la DBO5 y los niveles de saprobicidad dados por Hamm (1969).
Además, se realizó un análisis de correlación de Pearson utilizando el paquete estadístico SPSS, para estudiar las relaciones significativas (p< 0,05 o 0,01) entre las variables biológicas y ambientales.
Para los análisis estadísticos se incluyeron sólo las especies que tuvieron abundancia ≥ al 5 % en relación a la abundancia total de las especies estudiadas, ya que las poco frecuentes, de ocurrencia aleatoria pueden producir efectos espurios (Jongman et al., 1995). Las especies consideradas fueron: Aulacoseira granulata (Ehrenberg) Simonsen (Aulg), Cyclotella meneghiniana Kützing (Cyclm), Cymbella affinis Kützing (Cymba), Diatoma vulgare Bory (Diatv), Leptolyngbya foveolarum (Rabenhorst ex Gomont) Anagnostidis & Komárek (Leptf), Navicula sp. (Navisp) y Ulnaria ulna (Kützing) Compère (Ulnau). Asimismo se utilizaron las siguientes variables físicas y químicas: temperatura, pH, conductividad, oxígeno disuelto (OD), demanda bioquímica de oxígeno (DBO5), nitrato y ortofosfato (Tabla 1).
Tabla 1: Valores mínimos (mín.), máximos (máx.) de variables físicas y químicas de los ríos estudiados (marzo/1998 - marzo/1999)*.

* Tomado de Martínez De Marco et al. (2007).
Se aplicó el análisis multivariado de ordenación no métrico multidimensional (NMDS) con el fin de ordenar las muestras agrupadas por sitio (ríos Salí (tributario y emisario), Vipos y Tapia), temporada (1998-1999) y estación del año (20 unidades de muestras en total) según las siete especies algales mencionadas con anterioridad. Se utilizó la distancia Bray-Curtis como medida de similitud para la matriz de datos originales. Además, se midió el ajuste de este ordenamiento multivariado mediante el estadístico STRESS. Una limitación en el uso de NMDS es que las interpretaciones podrían ser cualitativas y subjetivas porque los ejes del mapa no son funciones de las variables originales y por lo tanto no pueden formularse hipótesis acerca de relaciones causales (James & McCulloch, 1990). Por tal razón, se realizó un análisis de correlación de Spearman no paramétrico para medir el grado de asociación lineal entre dos variables y así examinar la relación entre las especies en cada una de las muestras y los ejes extraídos del análisis del NMDS.
Para caracterizar las relaciones entre la abundancia de las especies algales y las variables físicas y químicas, junto con la estacionalidad y ubicación espacial de los sitios de muestreo, se usó el análisis de redundancia (RDA) (Legendre & Legendre, 2000). Se seleccionó RDA porque un examen preliminar de los datos mostró que la longitud de los tres primeros gradientes o ejes (2,59; 1,93 y 2,59) fueron cortos con respecto a las variables ambientales (Ter Braak & Šmilauer, 1998). Previo al análisis, se efectuaron transformaciones logarítmicas de las abundancias de las especies (log [abundancia+1]) y para remover la arbitrariedad en las unidades de medición de las variables físicas y químicas, las mismas fueron estandarizadas a media cero y unidad de varianza (Ter Braak, 1986).
La significancia de los ejes de ordenación del RDA fue analizada mediante un test de aleatorización por permutación sobre relaciones proyectadas. También se realizaron test de permutación para evaluar cuales variables explicativas aportaron significativamente al ordenamiento de las especies algales. Se realizaron biplots de especies y variables físicas y químicas del RDA para ilustrar las relaciones de gradientes entre especies algales y variables. Todos los análisis de datos fueron realizados mediante el paquete estadístico R versión 13.1 (R, 2010).
RESULTADOS
Densidad algal
En la Fig. 2 (a-d) se grafican las densidades totales del fitoplancton y de los grupos algales (Bacillariophyceae, Chlorophyta, Cyanobacteria, Dinophyta y Rhodophyta) para cada uno de los ríos estudiados. Los valores de abundancia, considerando diferentes rangos, de las especies detectadas en cada sistema lótico se exponen en la Tabla 2. Al comparar los intervalos de abundancia de todos los ríos se destacaron, en orden decreciente, los cuatro primeros con densidades algales entre 1 y 16 ind mL-1 y los rangos superiores a 128 ind mL-1 estuvieron representados en menor medida.
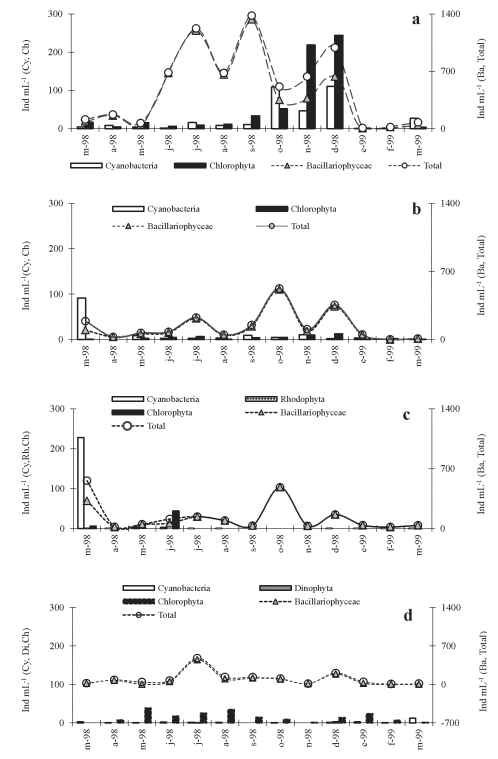
Fig. 2. Variaciones anuales de la densidad del fitoplancton en los ríos estudiados. a) río Salí (tributario), b) río Vipos, c) río Tapia, d) río Salí (emisario). Abreviaturas: Ba: Bacillariophyceae, Cy: Cyanobacteria, Ch: Chlorophyta; Di: Dinophyta; Rh: Rhodophyta.
Tabla 2: Valores de abundancia (por rangos) del fitoplancton de los ríos estudiados




Referencias de intervalos o rangos (ind mL-1 ): (*): 1-2; (+): 2-4; (□): 4-8; (x): 8-16; (◊): 16-32; (■): 32-64; (●): 64-128; (‡): 128-256; (▲): 256-512; (O): >512.
El fitoplancton del río Salí (tributario) tuvo un máximo de 1380 ind mL-1, en septiembre, sobresaliendo Diatoma vulgare Bory con 1179 ind mL-1. El valor mínimo fue obtenido para enero con 8 ind mL-1. Las Bacillariophyceae fueron dominantes seguidas, en general, por las algas verdes que adquirieron importancia numérica en noviembre y diciembre con 219 y 244 ind mL-1,respectivamente. Las Chlorophyta, representadas por Ankistrodesmus fusiformis Corda (187 ind mL-1) y Actinotaenium cucurbitum (Bisset) Teiling (188 ind mL-1), realizaron el mayor aporte a la densidad fitoplanctónica de los meses anteriormente citados. Las cianobacterias superaron a las algas verdes en cinco oportunidades con valores que oscilaron entre 2 y 109 ind mL-1, destacándose Leptolyngbyafoveolarum (Rabenhorst ex Gomont) Anagnostidis et Komárek, Merismopedia convoluta Brébisson y Merismopedia sp.
Con respecto a la densidad fitoplanctónica del río Vipos se encontraron tres valores relativamente altos que correspondieron a julio, octubre y diciembre de 1998 con registros totales de 225, 524 y 355 ind mL-1, respectivamente. El valor mínimo se registró para febrero con 3 ind mL-1. En esos meses la comunidad fitoplanctónica estuvo representada casi exclusivamente por las diatomeas pennadas destacándose Cymbella affinis Kützing, con valores que oscilaron entre 168 y 201 ind mL-1 y en octubre sobresalió Ulnaria ulna (Kützing) Compère con 206 ind mL-1. Se debe resaltar que únicamente en marzo de 1998 las cianobacterias tuvieron una igualdad numérica con las diatomeas pennadas, destacándose L. foveolarum con 91 ind mL-1. En el período estudiado las algas verdes fluctuaron entre 1 a 13 ind mL-1 y la densidad total de este grupo superó sólo en tres oportunidades a las cianobacterias.
En el caso del río Tapia la abundancia del fitoplancton mostró los valores máximos en marzo y octubre de 1998 con registros de 559 y 482 ind mL-1, respectivamente. El valor mínimo se obtuvo para abril y febrero con 20 ind mL-1. En el primer caso la comunidad estuvo casi exclusivamente compuesta por diatomeas pennadas (324 ind mL-1) y por L. foveolarum que llegó a 227 ind mL-1. En octubre, en cambio, sólo se registraron Bacillariophyceae, sobresaliendo C. affinis y D. vulgare. Las algas verdes fueron escasas y sólo se destacó Oocystis solitaria Wittrock en junio, mientras que las Rhodophyta únicamente se contaron en marzo de 1998.
El fitoplancton del sitio de muestreo correspondiente al río Salí (emisario), ubicado aguas abajo del embalse tuvo una densidad máxima de 449 ind mL-1, en julio de 1998. El valor mínimo se registró para febrero con 9 ind mL-1. Estuvo ampliamente dominado por las diatomeas céntricas
Aulacoseira granulata (Ehrenberg) Simonsen con una abundancia de 135 ind mL-1 (julio) y 132 ind mL-1 (diciembre) y Cyclotella meneghiniana Kützing con 293 ind mL-1 (julio). En segundo término se ubicaron las Chlorophyta siendo las más importantes: Closterium acutum var. variabile (Lemmermann) Geitler, O. solitaria y Staurastrum sp. En el mes de marzo de 1999 estos grupos fueron desplazados por las Cyanobacteria que se ubicaron en primer término con un valor total de 12 ind mL-1. Peridinium volzii Lemmermann y Glochidinium penardiformis Boltovskoy (Dinophyta) se registraron únicamente con valores bajos en julio y diciembre de 1998.
Biomasa algal (expresada como μg de clorofila a L-1)
Las concentraciones de biomasa algal (Fig. 3) encontradas en el río Salí (tributario) oscilaron entre 0,19 (marzo de 1998) y 6,59 µg Cl-a L-1 (junio de 1998), mientras que en el río Vipos la biomasa fluctuó ente 0,58 (abril de 1998) y 6,79 µg Cl-a L-1 (noviembre de 1998). La clorofila a del río Tapia fue mínima en febrero de 1999 con 0,52 µg Cl-a L-1 y la máxima se obtuvo en agosto de 1998 con 5,43 µg Cl-a L-1. Para el río Salí (emisario) el mayor valor de clorofila a encontrado fue de 5,62 µg Cl-a L-1 en julio de 1998 y los mínimos se registraron en noviembre de 1998 y febrero de 1999 con 0,58 µg Cl-a L-1.

Fig. 3. Fluctuaciones de la biomasa algal (expresada como clorofila a) durante el periodo estudiado.
Índice de diversidad específica de Shannon & Weaver
La Fig. 4 muestra las variaciones del índice de diversidad específica para todos los ríos. En el río Salí (tributario) fluctuó entre 1,2 (septiembre) y 4,2 (diciembre). La dominancia de D. vulgare con un valor de correlación de - 0,76 (p < 0,01), sería la responsable del valor mínimo, mientras que en el río Vipos, osciló entre 1,0 (julio) y 2,7 (enero y marzo de 1999). En este último río, si bien muchos de los valores encontrados fueron comparativamente bajos (inferiores a 2) no se obtuvo una correlación con las especies más abundantes. En el río Tapia el índice de diversidad osciló entre 0,2 (agosto) y 3,4 (mayo y marzo de 1998). Los valores bajos se debieron a la abundancia de D. vulgare y presentó una correlación negativa de - 0,56 (p < 0,05) con esta especie. El índice, en el río Salí emisario, fluctuó entre 0,7 (septiembre) y 3 (marzo de 1998). La dominancia de A. granulata fue la responsable de los bajos valores del índice con una correlación negativa de - 0,76 (p < 0,01).

Fig. 4. Variaciones del índice de diversidad específica de Shannon-Weaver del fitoplancton de los ríos estudiados.
Las correlaciones realizadas entre este índice y las variables ambientales para cada ambiente lótico sólo fueron significativas con la profundidad (0,70; p < 0,05) en el río Vipos y con la temperatura (0,61; p < 0,05) y la conductividad (- 0,67; p < 0,05) en el río Salí (emisario).
Índice sapróbico
Según los valores del índice sapróbico determinados para el río Salí (tributario) las aguas pudieron caracterizarse como oligosapróbicas a β-mesosapróbicas excepto en el mes de marzo de 1999 que fueron β-α-mesosapróbicas (críticamente poluído). En general, el río se pudo clasificar como no poluído durante los meses fríos y moderadamente poluído el resto del año. Las aguas del río Vipos se caracterizaron como oligosapróbicas a β-mesosapróbicas. De esta manera, el río se pudo clasificar como no poluído durante siete meses, ligeramente poluído para junio, julio de 1998 y enero de 1999 y moderadamente poluído para marzo de 1999. En el río Tapia, las aguas fueron caracterizadas como oligosapróbicas durante gran parte del año y oligo-β-mesosapróbicas en febrero de 1999, es decir, ligeramente poluídas. En abril de 1998 y marzo de 1999 se las pudo clasificar como moderadamente poluídas. La clasificación de las aguas del río Salí (emisario) fue oligosapróbicas de marzo a diciembre de 1998 y oligo-β-mesosapróbicas sólo en septiembre. En los meses cálidos de 1999 resultaron moderadamente poluídas (β-mesosapróbicas).
Análisis de ordenación no métrico multidimensional (NMDS)
La Fig. 5 muestra el ordenamiento de los sitios según la abundancia de las especies algales seleccionadas. La Tabla 3 presenta los valores de correlación no paramétrica entre los dos primeros ejes del NMDS y las abundancias de las siete especies algales. El valor de 0,06 del estadístico STRESS indicaría que hay un buen ajuste entre las variables observadas y el método de ordenamiento NMDS ya que es un valor cercano a cero y significativo según test de Monte Carlo (p < 0,05). El eje 1 (NMDS 1) ubica hacia la izquierda las muestras del río Salí emisario de los veranos 1998 y 1999 que tuvieron las menores abundancias de Cymbella affinis, Diatoma vulgare, Navicula sp. y Ulnaria ulna. De la misma manera, hacia la derecha se encuentran las muestras de otoño, invierno y primavera del río Salí tributario y primavera de los ríos Vipos y Tapia con las mayores abundancias de las especies anteriormente citadas. En el extremo negativo del eje 2 se ubica el río Salí emisario (otoño, invierno y primavera) por presentar las mayores abundancias de Aulacoseira granulata y Cyclotella meneghiniana y menores de Leptolyngbya foveolarum. En el lado positivo de este eje se encuentran los veranos de todos los tributarios con densidades menores de A. granulata yC. meneghiniana y mayores de L. foveolarum.

Fig. 5. Análisis de Ordenación no Métrico Multidimensional (NMDS) de los sitios estudiados según las especies consideradas. Abreviaturas: S: río Salí (tributario), V: río Vipos, T: río Tapia, E: río Salí (emisario); V: verano, O: otoño, I: invierno y P: primavera.
Tabla 3: Valores de correlación de Spearman de los ejes NMDS1 y NMDS2 con las abundancias de las especies algales.

Referencias: ns: no significativa; * significativa p<0,05.
Análisis de Redundancia (RDA)
Por medio de un RDA se relacionó la distribución de las especies seleccionadas y las variables físicas y químicas.
El valor de inercia total del RDA fue de 0,29 y el modelo respondió efectivamente a los datos observados (F: 3,17; p < 0,05; N = 199 permutaciones). Resultaron significativos tres ejes RDA (F: 58,8; 20,8; 8,12 correspondientes a RDA1, RDA2 y RDA3 con p < 0,05 y 199 permutaciones). Los tres primeros ejes acumularon un total de 86 % de la variabilidad en los datos de abundancia de las especies algales (Tabla 4).
Tabla 4: Eigenvalues y varianzas de los ejes 1, 2 y 3 del RDA

La Tabla 5 indica cuales variables físicas y químicas resultaron significativas en el ordenamiento de las especies algales mediante el RDA. Estas variables fueron temperatura, conductividad eléctrica, pH y NO3. En este análisis no se colocaron como variables a la ubicación de los sitios.
Tabla 5: Coeficientes de correlación lineal intersets entre las variables físicas y químicas y los tres primeros ejes del RDA que describen la comunidad algal durante las temporadas 1998 y 1999.

Referencia: (*) Significancia estadística a p<0,1
A través de la Fig. 6 (a-c, biplots) teniendo en cuenta no sólo las variables significativas sino también aquéllas que presentan valores de correlación mayores a 0,40 (Tabla 5); se interpretaron cada uno de los tres ejes de RDA de las figuras basadas en puntuaciones de las especies y variables ambientales.
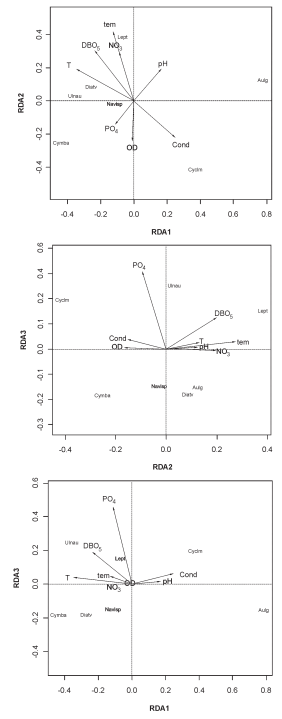
Fig. 6. Análisis de Redundancia (RDA): Gráficos entre variables físicas y químicas y especies algales, a: de los ejes RDA 1 y RDA 2; b: de los ejes RDA 1 y RDA 3 y c: de los ejes RDA 2 y RDA 3. Las variables están representadas por flechas.
En el eje 1 o RDA1 las especies Cyclotella meneghiniana (Cyclm) y Aulacoseira granulata (Aulg) respondieron positivamente con sus abundancias a valores bajos de temperatura, altos de conductividad, presentes en río Salí (emisario) y ausentes en el río Vipos y Salí (tributario). En condiciones opuestas a las mencionadas se encontraron Ulnaria ulna (Ulnau) y Cymbella affinis (Cymba).
En el eje 2 o RDA 2, Leptolyngbya foveolarum (Lept) respondió positivamente con sus abundancias a valores altos de pH y nitrato, con densidades mayores en 1999, presente en otoño, con registros altos de DBO5, ausente en primavera en el río Tapia. En extremo contrario se ubicó Cyclm.
En el eje 3 o RDA 3, se observaron dos grupos separados de especies algales. En el extremo positivo del eje se encontraron Ulnau, Lept y Cyclm y en el extremo negativo el resto. Este eje se correlacionó significativamente con el ortofosfato.
DISCUSIÓN Y CONCLUSIONES
Los resultados obtenidos mostraron que la abundancia algal fue máxima en el período invernal o primaveral, excepto en el río Tapia donde ocurrió en el mes de marzo de 1998. Los valores mínimos coincidieron con los meses cálidos. Esta disminución de la densidad del fitoplancton podría atribuirse a una dilución provocada por el notable aumento del caudal de los ríos estudiados, que se caracterizan por las marcadas diferencias de éstos a lo largo del año. Durante el verano los ríos aumentan considerablemente y las crecientes se producen principalmente entre diciembre y marzo debido a que más del 90 % de las precipitaciones se concentran durante estos meses (Sesma et al., 1998; Georgieff, 2007). Por carecer de valores de caudales para el período estudiado, en la Tabla 6 se presentan los datos promedios históricos de los sistemas fluviales analizados, excepto para el Tapia (Anónimo, 2004). Estos registros de 18 a 48 años de mediciones, según los ríos, concordaron con nuestras observaciones para el período 1998-1999. Una situación similar se registró en ríos de Córdoba donde las fluctuaciones estacionales de la densidad del fitoplancton mostraron un valor máximo al comienzo del invierno y un mínimo en el período de aguas altas (e.j. Luque & Martínez de Fabricius, 2003, 2010, entre otros). Este patrón también se podría homologar a lo observado en muchos ríos tropicales y subtropicales donde las comunidades fitoplanctónicas no alcanzan una densidad alta debido a los niveles elevados de turbidez causados por los sólidos suspendidos aportados por procesos erosivos de los suelos (Meybeck et al., 1992).
Tabla 6: Caudales medios mensuales de los ríos (m3 seg-1)

Tomado de: Anónimo (2004)
El potamoplancton estuvo conformado casi íntegramente por las diatomeas en los meses de julio y agosto, destacándose las pennadas en los tributarios del embalse, cuyos representantes más numerosos fueron Diatoma vulgare y Cymbella affinis. Asimismo en el río Salí, aguas abajo del embalse C. Gelsi, se notó la dominancia de las diatomeas céntricas tales como Aulacoseira granulata y Cyclotella meneghiniana, que sobresalieron en este ambiente léntico artificial (Tracanna et al., 2006). El mayor número de diatomeas puede atribuirse a su gran eficiencia fotosintética, contenido alto de clorofila y un bajo umbral de saturación de luz, que le confiere ventajas adaptativas con respecto a otros componentes del plancton en ambientes turbios (Reynolds, 1988). Las especies de Aulacoseira son características de sistemas turbulentos y sometidos a constantes cambios en la calidad de luz debido a sus movimientos a través del gradiente lumínico y generalmente son citadas para ambientes leníticos (Gómez, 1991; Tracanna & Seeligmann, 1992; Tracanna et al. 1999; Salusso, 2005; Seeligmann & Tracanna, 2009) y grandes ríos como el Paraná y Uruguay (García de Emiliani, 1981; O'Farrell et al., 1996; Zalocar de Domitrovic & Maidana, 1997, entre otros). Las algas verdes que se ubicaron en segundo lugar en el río Salí, tanto tributario como emisario, fueron escasas en Vipos y en Tapia. Las cianobacterias codominaron en los ríos Vipos y Tapia en el mes de marzo de 1998, alcanzando el 35 % del total del fitoplancton. Esto podría atribuirse a condiciones favorables tales como incrementos de la temperatura, una alta irradiación, entre otros factores (Reynolds, 1972; Zalocar de Domitrovic et al., 1998; Paerl & Huisman, 2009). Los dinoflagelados, Peridinium volzii y Glochidinium penardiforme, sólo fueron observados en el río Salí (emisario) y Chantransia sp. fue registrada únicamente para Tapia.
Con respecto a la clorofila a en los meses cálidos se midieron los menores registros conjuntamente con las bajas densidades de fitoplancton. Esto estaría vinculado al aumento del caudal de los ríos y a la elevada turbidez de las aguas. En diciembre se verificó una gran uniformidad en los valores de biomasa algal de los cuatro ríos. Los bajos promedios encontrados se asemejaron a los mencionados por Mirande (2001) para el río Gastona perteneciente a la cuenca Salí-Dulce, aunque este último posee una importante contaminación debida principalmente a la actividad azucarera y a otros ríos no poluídos del noroeste argentino (Salusso, 2005).
Existen distintos criterios sobre la efectividad de la aplicación de los índices de saprobicidad y diversidad específica para evaluar la calidad del agua (Lobo et al., 1996; Gómez, 1998, 1999). El índice de Shannon & Weaver se ha utilizado en limnología para estimar, entre otras cosas, la posible contaminación. De acuerdo a este índice, valores entre 1 y 3 indicarían una contaminación moderada y los mayores a 3 buena calidad de agua (Branco, 1984). Este índice demostró variaciones de buena a moderada calidad del agua dependiendo de cada río. Numerosos autores consideran que este coeficiente no siempre es una herramienta eficiente en la determinación de la contaminación (e.j. Archibald, 1972; del Giorgio et al., 1991). Por tal motivo se aplicó el índice de saprobicidad de Pantle & Buck, que definió a las aguas de los ríos como "no poluídas" o "ligeramente poluídas" en los meses fríos y como "moderadamente poluídas" en los cálidos, ampliándose en el caso del río Salí (tributario) a otras épocas del período estudiado.
Los índices de diversidad y saprobiedad coincidieron en la caracterización de la calidad del agua para los ríos Salí (tributario) y Tapia. En el caso de los ríos Vipos y Salí (emisario) fue el índice de saprobiedad el que reflejó de manera más eficiente la diferenciación de los meses donde hubo una mayor contaminación orgánica.
El NMDS permitió ordenar espacial y temporalmente a las muestras de los diferentes ríos según las abundancias de las siete especies algales seleccionadas. Los dos ejes separaron notoriamente las muestras de todas las estaciones del río Salí, emisario. El NMDS 1 segregó hacia la parte positiva las muestras correspondientes al período de estiaje de los tributarios, mientras que el eje 2 lo hizo con las del período estival de estos últimos.
El RDA que incluyó variables físicas, químicas y biológicas permitió ordenar espacial y temporalmente a los ríos estudiados (Tabla 5). Se detectaron dos patrones de variación: el primero fue espacial asociado al embalse C. Gelsi que evidenció su efecto moderador y su influencia sobre la taxocenosis del fitoplancton del río Salí emisario en especial por la abundancia de Cyclotella meneghiniana y Aulacoseira granulata (Tracanna et al., 2006). El segundo patrón fue temporal y se relacionó con gradientes térmicos, de conductividad eléctrica (mineralización), de pH, de polución orgánica, de concentración de nitrato y de ortofosfato y de abundancia de Cymbella affinis, Ulnaria ulna y Leptolyngbya foveolarum.
Debido a la importancia que tiene la cuenca de Tapia-Trancas para el abastecimiento del embalse C. Gelsi y por ende la provisión de agua potable a la ciudad capital, se debería proceder a la sistematización de la cuenca alta, a la conservación de los suelos junto con una explotación racional de sus recursos y relevamientos permanentes de las comunidades biológicas y variables físicas y químicas del agua.
AGRADECIMIENTOS
Este trabajo fue financiado por el Consejo de Investigaciones de la Universidad Nacional de Tucumán y por la Fundación Miguel Lillo. Asimismo, deseamos expresar nuestro más sincero agradecimiento a la Dra. Mariela Alderete por su valiosa colaboración y asesoramiento en el análisis estadístico de los resultados y su posterior interpretación, como así también, al Geol. Alberto Gutiérrez por la realización de las figuras de esta publicación.
BIBLIOGRAFÍA
1. ANÓNIMO. 2004. Estadística hidrológica de la República Argentina. Ministerio de Planificación Federal, Inversión Pública y Servicios, Secretaria de Obras públicas, Subsecretaría de Recursos Hídricos, Evarsa, Buenos Aires. [ Links ]
2. ARCHIBALD, R. M. 1972. Diversity in some South African diatom associations and its relation to water quality. Water Res. 6: 1229-1238. [ Links ]
3. BRANCO, S. M. 1984. Limnología Sanitaria, estudio de la polución de aguas continentales. Monografía de la Secretaría General de la Organización de los Estados Americanos, Serie de Biología 28, Washington D.C. [ Links ]
4. DEL GIORGIO, P., A. L. VINOCUR, R. J. LOMBARDO & H. G. TELL. 1991. Progressive changes in the structure and dynamics of the phytoplankton community along a pollution gradient in a lowland river - a multivariate approach. Hydrobiologia 224: 129-154. [ Links ]
5. FERNÁNDEZ, D., M. DEL V. HIDALGO & D. CICERONE. 2007. Humedales de la provincia de Tucumán. En: CICERONE, D. S. & M. HIDALGO (eds.), Los humedales de la cuenca del río Salí, Argentina, pp. 201-210. Jorge Baudino Ediciones, Bs. As. [ Links ]
6. GARCÍA, M. G., M. DEL V. HIDALGO & M. A. BLESA. 2007. Impacto del hombre sobre la calidad del agua en los humedales de la cuenca del río Salí, Provincia de Tucumán, Argentina. En: CICERONE, D. S. & M. HIDALGO (eds.), Los humedales de la cuenca del río Salí, Argentina, pp. 127-144. Jorge Baudino Ediciones, Bs. As. [ Links ]
7. GARCÍA DE EMILIANI, M. O. 1981. Fitoplancton de los principales cauces y tributarios del valle aluvial del río Paraná: tramo Goya-Diamante. Rev. Asoc. Cienc. Nat. del Litoral 12: 112-125. [ Links ]
8. GARRIDO, H. B. 2005. Población y tierra en la cuenca de Trancas, provincia de Tucumán (República Argentina). Cuad. Desarrollo Rural 54: 33-60. [ Links ]
9. GEORGIEFF, S. M. 2007. Aspectos hidrológicos de la cuenca del río Salí. En: CICERONE, D. S. & M. DEL V. HIDALGO (eds.), Los Humedales de la Cuenca del Río Salí. Argentina, pp. 29-48. Jorge Baudino Ediciones, Bs. As. [ Links ]
10. GÓMEZ, N. 1991. Flora diatomológica del Embalse Río III (Prov. Córdoba, Argentina) I. Centrales. Gayana Bot. 48: 3-9. [ Links ]
11. GÓMEZ, N. 1998. Use of epipelic diatoms for evaluation of water quality in the Matanza-Riachuelo (Argentina), a pampean plain river. Wat. Research 32: 2029-2034. [ Links ]
12. GÓMEZ, N. 1999. Epipelic diatoms from the Matanza-Riachuelo River (Argentina), a highly polluted basin from the pampean plain: biotic indices and multivariate analysis. Aquat. Ecosyst. Health Managem. 2: 301-309. [ Links ]
13. HAMM, A. 1969. Die Ermittlung der Gewässergüteklassen bei Fließgewässern nach dem Gewässergütesystem und Gewässergütenomogramm. Münchner Beitr. Abwass. Fischerei und Flussbiologie 15: 46-48. [ Links ]
14. JAMES, F. & C. MCCULLOCH. 1990. Multivariate analysis in ecology and systematics: panacea or Pandora's Box?. Annu. Rev. Ecol. Syst. 21: 129-66. [ Links ]
15. JONGMANN, R. H. G., C. J. F. TER BRAAK & O. F. R. VAN TONGEREN. 1995. Data analysis in community and landscape ecology. Cambridge University Press, Cambridge, U.K. [ Links ]
16. LEGENDRE, P. & L. LEGENDRE. 2000. Numerical Ecology. Second English Edition, Elsevier Science B.V., Amsterdam. [ Links ]
17. LOBO, E. A., V. L. M. CALLEGARO, M. A. OLIVEIRA, S. E. SALOMONI, S. SCHULER & K. ASAI. 1996. Pollution tolerant diatoms from lotic systems in the Jacuí Basin, Rio Grande do Sul, Brazil. Iheringia Ser. Bot. 47: 45-72. [ Links ]
18. LOEZ, C. R. 1995. Determinación de clorofila a. En: TELL, G. & E. C. LOPRETTO (eds.), Ecosistemas de aguas continentales. Metodología para su estudio. Tomo 1, pp. 263-269. Ediciones Sur, Bs. As. [ Links ]
19. LUQUE, M. E. & A. L. MARTÍNEZ DE FABRICIUS. 2003. Distribución temporal del fitoplancton y epiliton en el río Piedra Blanca (Córdoba, Argentina). Limnetica 22: 19-34. [ Links ]
20. LUQUE, M. E. & A. L. MARTÍNEZ DE FABRICIUS. 2010. Estudio del componente algal en la cuenca baja del río Cuarto (Córdoba, Argentina). Lilloa 47: 95-106. [ Links ]
21. MARTÍNEZ DE MARCO, S. N., B. C. TRACANNA, M. A. BARRIONUEVO, M. G. NAVARRO, G. S. MEONI & A. P. CHAILE. 2007. Fluctuaciones de las variables físicas y químicas de los ríos Salí, Vipos y Tapia (Tucumán, Argentina). Lilloa 44: 19-38. [ Links ]
22. MARTÍNEZ DE MARCO, S. N., B. C. TRACANNA & N. MAIDANA. 2011. Fitoplancton de ambientes lóticos de la cuenca superior del río Salí, Tucumán, Argentina. Lilloa 48: 100-123. [ Links ]
23. MEYBECK, M., G. FRIEDRICH, R. THOMAS & D. CHAPMAN. 1992. Rivers. In: CHAPMAN, D. (ed.), Water Quality Assessments: a guide to use of biota, sediments and water in environmental monitoring, 2nd ed., pp. 239-316. Chapman & Hall, London. [ Links ]
24. MIRANDE, V. 2001. Dinámica del fitoplancton del río Gastona (Tucumán, Argentina) en relación a la calidad de sus aguas. Tesis Doctoral, FCN e IML, UNT. [ Links ]
25. O'FARRELL, I., I. IZAGUIRRE & A. VINOCUR. 1996. Phytoplankton ecology of the Lower Paraná River (Argentina). Arch. Hydrobiol. Suppl. 115/Large Rivers, 11: 75-89. [ Links ]
26. PAERL, H. W. & J. HUISMAN. 2009. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 1: 27-37. [ Links ]
27. PÁEZ, S. V. 2007. Análisis de los ambientes geomorfológicos de la cuenca del Río Acequiones (Trancas, Tucumán) utilizando fotointerpretación y factores climáticos. Acta Geol. Lilloana 20: 19-28. [ Links ]
28. PANTLE, R. & H. BUCK. 1955. Die biologisch Überwachung der Gewässer und die Darstellung der Egerbnisse. Gas- und Wasserfach 96: 604-607. [ Links ]
29. PATRICK, R. 1963. The structure of diatom communities under varying ecological conditions. Ann. New York Acad. Sci. 108: 359-365. [ Links ]
30. REYNOLDS, C. S. 1972. Growth, gas vacuolation and buoyancy in a natural population of planktonic blue-green algae. Freshwater Biol. 2: 87-106. [ Links ]
31. REYNOLDS, C. S. 1988. Functional morphology and adaptative strategies of freshwater phytoplankton. In: SANDGREN, C. (ed.), Growth and reproductive strategies of freshwater phytoplankton, pp. 388-433. Cambridge University Press, London. [ Links ]
32. R DEVELOPMENT CORE TEAM (2010). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.or. [ Links ]
33. ROS, J. 1979. Prácticas de ecología. Eds. Omega, Barcelona. [ Links ]
34. SALUSSO, M. M. 2005. Evaluación de la calidad de los recursos hídricos superficiales en la alta cuenca del Juramento (Salta). Tesis Doctoral, Universidad de Buenos Aires. [ Links ]
35. SHANNON, C. E. & W. WEAVER. 1963. The mathematical theory of communication. Univ. Illinois Press, Urbana. [ Links ]
36. SEELIGMANN, C. & B. C. TRACANNA. 2009. Dinámica del fitoplancton en un embalse de alta cota del Noroeste Argentino (Tucumán). Limnetica 28: 105-124. [ Links ]
37. SESMA, P., E. GUIDO & M. PUCHULU. 1998. Clima de la provincia de Tucumán: En: GIANFRANCISCO, M., M. PUCHULU, J. DURANGO DE CABRERA & G. ACEÑOLAZA (Eds.), Geología de Tucumán, pp. 41-46. Publicación Esp. del Colegio de Graduados en Ciencias Geológicas de Tucumán, Tucumán. [ Links ]
38. TER BRAAK, C. J. F. 1986. Canonical correspondence analysis: a new eigenvector method for multivariate direct gradient analysis. Ecology 67: 1167-1179. [ Links ]
39. TER BRAAK, C. J. F. & P. MILAUER. 1998. CANOCO reference manual and user's guide to CANOCO for Windows: software for canonical community ordination (Version 4). Ithaca, New York: Microcomputer Power. [ Links ]
40. TRACANNA, B. C. & C. SEELIGMANN. 1992. Limnología del Embalse El Cadillal - Tucumán, Argentina - III: Estudio cuantitativo del fitoplancton. Lilloa 38: 65-75. [ Links ]
41. TRACANNA, B. C., A. VILLAGRA DE GAMUNDI, C. SEELIGMANN, C. LOCASCIO DE MITROVICH & E. DRAGO. 1991. Limnología del embalse El Cadillal (Tucumán - Argentina) I: Variables morfométricas y fisicoquímicas. Rev. Hydrobiol. Trop. 24: 269-286. [ Links ]
42. TRACANNA, B. C., C. T. SEELIGMANN, V. MIRANDE, L. B. DE PARRA, M. T. DE PLAZA & F. M. MOLINARI. 1999. Cambios espaciales y temporales del fitoplancton en el embalse Río Hondo (Argentina). Bol. Soc. Argent. Bot. 34: 101-105. [ Links ]
43. TRACANNA, B. C., S. N. MARTÍNEZ DE MARCO, M. J. AMOROSO, N. ROMERO, P. CHAILE & A. MANGEAUD. 2006. Physical, chemical and biological variability in the Dr. C. Gelsi reservoir (NW Argentine): A temporal and spatial approach. Limnetica 25: 787-808. [ Links ]
44. VILLAGRA DE GAMUNDI, A., C. SEELIGMANN DE SOSA GÓMEZ, B. C. TRACANNA & C. LOCASCIO DE MITROVICH. 1993. Sobre la limnología fisicoquímica del río Salí y sus tributarios en áreas próximas al embalse El Cadillal (Tucumán, Argentina). Miscelánea 91, Fundación Miguel Lillo, Tucumán. [ Links ]
45. ZALOCAR DE DOMITROVIC, Y. & N. I. MAIDANA. 1997. Taxonomic and ecological studies of the Paraná River diatom flora (Argentina). Bibl. Diatomol. 14. J. Cramer, Berlin. [ Links ]
46. ZALOCAR DE DOMITROVIC, Y., V. M. ASSELBORN & S. L. CASCO. 1998. Variaciones espaciales y temporales de un lago subtropical de Argentina. Revista Bras. Biol. 58: 359-382. [ Links ]
Recibido el 16 de septiembre de 2011
Aceptado el 24 de marzo de 2012.

