










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.47 no.3-4 Córdoba jul./dic. 2012
BIOLOGÍA REPRODUCTIVA
Caracterización y variación espacio-temporal del néctar en anarthrophyllum desideratum (Fabaceae): Influencia del clima y los polinizadores
Valeria Paiaro1,3, Gabriel E. Oliva2, Andrea A. Cocucci1 y Alicia N. Sérsic1
1 Laboratorio de Ecología Evolutiva - Biología Floral. Instituto Multidisciplinario de Biología Vegetal (IMBIV), CONICET-Universidad Nacional de Córdoba. CC 495, 5000 Córdoba.
2 INTA - Estación Experimental Santa Cruz. Universidad Nacional de la Patagonia Austral. Unidad Académica Río Gallegos. 9400 Río Gallegos.
3 E-mail: valepaiaro@yahoo.com
Resumen: En este trabajo se caracterizó el volumen, la concentración y la composición de azúcares del néctar floral en Anarthrophyllum desideratum (DC) Benth., un arbusto endémico de la estepa Patagónica polinizado por aves Passeriformes, y se evaluó su variación espacio-temporal. Se analizaron las diferencias entre áreas ecológicas, poblaciones, plantas y años, y las relaciones entre rasgos, la temperatura, precipitación y tasa de visitas de las poblaciones. El néctar mostró un volumen promedio de 8,1 µl, una concentración de 28% y estuvo dominado por hexosas, como en otras especies ornitófilas. Los atributos del néctar estuvieron correlacionados a nivel poblacional, y las diferencias entre poblaciones explicaron la mayor parte de su variabilidad espacial. El volumen y la concentración variaron entre años, mientras que la composición de azúcares se mantuvo constante. La proporción de glucosa disminuyó con el aumento de la temperatura, pero el resto de los caracteres no se asoció con las variaciones climáticas de la región. El volumen de néctar disminuyó a medida que aumentó la tasa de visitas de las poblaciones. Los resultados sugieren la existencia de una importante variación espacio-temporal del néctar en A. desideratum que no respondería a gradientes ambientales simples sino a un escenario complejo con múltiples factores.
Palabras clave: Anarthrophyllum desideratum; Área ecológica; Composición de azúcares; Concentración; Passeriformes; Población; Precipitación; Temperatura; Tasa de visitas; Volumen de néctar.
Summary: Characterization and spatio-temporal variation of nectar in Anarthrophyllum desideratum (Fabaceae): Influence of climate and pollinators. In this study we characterized the volume, concentration and sugar composition of floral nectar in Anarthrophyllum desideratum (DC) Benth., an endemic shrub from the Patagonian steppe pollinated by Passerine birds, and we evaluated it spatio-temporal variation. We analyzed the differences among ecological areas, populations, plants and years, and the relationships between traits, the temperature, rainfall and visitation rate of populations. Nectar showed an average volume of 8.1 µl, a concentration of 28% and was dominated by hexoses, as in other ornithophilous species. The nectar attributes were correlated at population level, and the differences among populations accounted for most of its spatial variability. Volume and concentration varied among years, whereas sugar composition remained constant. The proportion of glucose decreased with increasing temperature, but the rest of the characters did not associate with climatic variations of the region. Nectar volume decreased with increasing the visitation rate of populations. The results suggest the existence of an important spatio-temporal variation of nectar in A. desideratum that would not respond to simple environmental gradients but to a complex scenario with multiple factors.
Key words: Anarthrophyllum desideratum; Concentration; Ecological area; Nectar volume; Passeriformes; Population; Sugar composition; Precipitation; Temperature; Visitation rate.
INTRODUCCIÓN
En las plantas que se reproducen sexualmente, el néctar es la principal recompensa floral (Simpson & Neff, 1983), siendo de vital importancia en la atracción de los polinizadores. Entre los distintos constituyentes químicos del néctar (azúcares, aminoácidos, lípidos, fenoles y antioxidantes), los azúcares dominan el total de solutos y representan la principal fuente de energía para los visitantes (Baker & Baker, 1983a). Los néctares de la mayoría de las especies contienen tres azúcares principales, sacarosa (disacárido), glucosa y fructosa (hexosas), en una proporción característica (Percival, 1961; Baker & Baker, 1983a, b). Otras propiedades importantes del néctar que contribuyen al éxito de la polinización son la concentración de azúcares (Baker, 1975; Pyke & Waser, 1981) y la tasa de producción (Cruden et al., 1983; Pleasants, 1983) que se refeja en el volumen.
El volumen, la concentración y la composición de azúcares del néctar son generalmente asociados con el grupo funcional de polinizadores o el síndrome floral de las plantas (Faegri & van der Pijl, 1979; Pyke & Waser, 1981; Baker & Baker, 1983b; Baker et al., 1998). Así, las flores de las especies polinizadas por aves son conocidas por secretar grandes volúmenes de néctar relativamente diluidos, en comparación con aquellas visitadas por insectos (Faegri & van der Pijl, 1979; Proctor et al., 1996; Nicolson, 2002; Fenster et al., 2004). En cuanto a su composición, el néctar de las flores visitadas por colibríes, insectos de probóscides largas y mariposas diurnas y nocturnas suele poseer grandes proporciones de sacarosa, mientras que proporciones abundantes de hexosas son características de los néctares de flores polinizadas por murciélagos, aves Passeriformes, moscas y abejas de probóscides cortas (Baker & Baker, 1983b; Proctor et al., 1996; Galetto & Bernardello, 2003; Dupont et al., 2004; Chalcoff et al., 2006).
El hecho de que los néctares ricos en hexosas de las especies polinizadas por Passeriformes sean además diluidos podría explicarse por una presión selectiva dependiente de la concentración ejercida por las aves, que a altas concentraciones de azúcar prefieren la sacarosa por sobre las hexosas, mientras que a bajas concentraciones prefieren glucosa y fructosa (Schondube & Martínez del Rio, 2003). Alternativamente, la asociación entre la composición de azúcares y la concentración del néctar en dichas especies podría ser el resultado de procesos osmóticos (Nicolson, 2002). En el nectario, la savia del floema rica en sacarosa es hidrolizada en glucosa y fructosa por la enzima invertasa (Pate et al., 1985), incrementando la osmolaridad del néctar, y el flujo de agua resultante llevaría a una disminución de la concentración y un incremento de volumen. Estas hipótesis no han sido evaluadas a nivel intra-específico.
Si bien la mayoría de las variaciones en las características del néctar han sido atribuidas a las diferencias en el síndrome de polinización, una variación sustancial del néctar también existe entre especies dentro de un síndrome floral, entre especies visitadas por el mismo polinizador y dentro de una especie con un mismo tipo de visitante floral (Brown & Kodric-Brown, 1979; Brink & DeWet, 1980; Pleasants, 1981). Dentro de una especie, los atributos del néctar pueden variar espacialmente entre regiones geográficas, entre poblaciones, entre plantas dentro de una población y entre flores dentro de una planta (por ej. Bertsch, 1983; Pleasants & Chaplin, 1983; Herrera, 1995; Lanza et al., 1995; Devoto et al., 2006), y temporalmente entre años, meses, días y a lo largo del día (por ej., Freeman & Head, 1990 y referencias allí citadas). Dichas diferencias pueden responder a las variaciones espaciales y temporales de factores abióticos tales como la temperatura y la humedad, los cuales pueden afectar la producción de néctar y provocar cambios post-secreción (Corbet et al., 1979; Bertsch, 1983; Freeman & Head, 1990; Villarreal & Freeman, 1990). El efecto de las condiciones climáticas sobre el néctar puede ser directo a través de la dilución por precipitaciones o de la evaporación por temperatura, sequía y viento, o indirecto por evapotranspiración y déficit hídrico de las plantas (Corbet et al., 1979). El clima también puede afectar al néctar de manera indirecta, a través de su efecto sobre la actividad de los visitantes florales (Heinrich & Raven, 1972; Calder, 1979; Herrera, 1995).
Mientras parece claro que existen correlaciones entre las características del néctar y el tipo de polinizador, actualmente no se sabe con precisión cuánto y cómo varían estos rasgos dentro de un grupo de polinizadores. Además, los datos disponibles acerca de los factores ambientales que pueden alterar el volumen, la concentración y la composición del néctar son contradictorios (Villarreal & Freeman, 1990 y referencias allí citadas). Es entonces importante estimar la contribución relativa de los diferentes niveles de organización biológica a la variabilidad total de los caracteres del néctar dentro de las especies así como evaluar su respuesta a un ambiente dinámico. Las plantas que se distribuyen geográficamente en un ambiente heterogéneo y a la vez poseen un grupo funcional de polinizadores homogéneo en su rango geográfico y estable a lo largo de diferentes temporadas ofrecen la oportunidad de explorar la influencia del ambiente abiótico en un contexto biótico relativamente constante.
Anarthrophyllum desideratum (DC) Benth. (Fabaceae) es un arbusto endémico de la estepa Patagónica que se distribuye geográficamente en un área extensa que presenta considerable heterogeneidad ambiental, incluyendo diversasáreas ecológicas, identificadas como unidades ambientales dentro de la región (Oliva et al., 2001). Es el primer caso documentado de polinización por aves Passeriformes en la estepa Patagónica, dentro de su rango de distribución presenta ensambles de visitantes florales variables pero funcionalmente equivalentes (Paiaro, 2011). Si bien existe gran cantidad de información acerca de las plantas nectaríferas de la Patagonia y su relación con los síndromes de polinización (Forcone et al., 1997; Bernardello et al., 1999; Galetto & Bernardello, 2003; Chalcoff et al., 2006; Devoto et al., 2006), la mayoría de los trabajos han explorado los constituyentes del néctar a nivel inter-específico y ninguno de ellos ha incluido a esta especie ornitófila característica de las comunidades esteparias. Por su parte, los únicos estudios de néctar a nivel intra-específico en Patagonia fueron realizados en una especie propia de los Bosques Andino-Patagónicos con un ensamble de visitantes florales geográficamente variable y en un área relativamente pequeña (Chalcoff et al., 2006; Devoto et al., 2006).
En el presente estudio, en primer lugar, se caracteriza el néctar floral de Anarthrophyllum desideratum en función de su volumen, concentración y composición de azúcares, con el objetivo de evaluar si estas características se corresponden con las de otras especies ornitófilas. En segundo lugar, se cuantifica su variación espacial (entre áreas ecológicas, entre poblaciones y entre plantas) y temporal (entre años), se analizan las relaciones entre sus distintos componentes, y entre éstos, los factores climáticos y la tasa de visitas de las poblaciones, para determinar si cambios significativos en el volumen, la concentración y la composición del néctar pueden detectarse dentro de esta especie y atribuirse a factores diferentes al grupo funcional de polinizadores, como el estrés hídrico, la temperatura o la actividad de los visitantes florales.
MATERIALES Y MÉTODOS
Especie en estudio
Anarthrophyllum desideratum (DC) Benth. (Fabaceae, Papilionoideae-Genisteae) es un arbusto enano espinoso endémico de la Patagonia Austral (Sorarú, 1974). En Argentina, se distribuye a lo largo de 1000 Km desde los 43º10' hasta los 51º60' de latitud Sur, mientras que en Chile está restringido a la porción continental más austral, en el área del Parque Nacional Torres del Paine (51°00' S, 72°50' O). Fitogeográficamente, la especie ocupa principalmente la provincia Patagónica, que incluye las partes áridas y semiáridas de la estepa, y sólo parte de la provincia Subantártica, donde está presente en los pastizales subandinos, pero no en el bosque (Roig, 1998). Esta región presenta una importante heterogeneidad ambiental, con elevaciones que varían desde el nivel del mar hasta los 900 m de altitud, marcados gradientes climáticos, una variedad de ambientes topográfcos y edáficos, y áreas ecológicas muy diversas (Tabla 1) definidas en función del clima, el suelo y la vegetación (Oliva et al., 2001).
Tabla 1. Coordenadas geográficas e información climática de las 25 poblaciones estudiadas de Anarthrophyllum desideratum en la Patagonia Austral. Cód. = código de la población; los datos climáticos corresponden al período de floración de la especie.

* Obtenida a partir de la base de datos de Worldclim Global Climate GIS (disponible on-line en: http://www.worldclim.org).
La floración de A. desideratum ocurre desde mediados de Octubre a mediados de Noviembre. Las plantas producen un gran número de flores solitarias, típicamente papilionadas, con colores uniformes que van del rojo al anaranjado (Sorarú, 1974), y abundante cantidad de néctar expuesto en forma de gota entre el estandarte y la quilla (V. Paiaro, obs. pers.). El color, el tamaño y la forma floral varían considerablemente entre las poblaciones naturales de la especie en asociación con las variaciones del ambiente abiótico (Paiaro et al., en prensa). Aunque las plantas son parcialmente auto-compatibles, la reducida autopolinización espontánea y la presencia de protandria parcial (A. Sérsic, obs. pers.) indicarían una dependencia de los polinizadores para la transferencia de polen (Paiaro, 2011). Las flores son polinizadas por distintas especies de aves Passeriformes, principalmente Emberízidos (Paiaro, 2011), las cuales debido a su tamaño y comportamiento pueden considerarse funcionalmente equivalentes. Las semillas poseen un mecanismo balístico de dispersión y son arrojadas a unos pocos metros de la planta madre.
Procedimiento de muestreo y caracteres medidos
En todo el rango geográfico de la especie un total de 25 poblaciones de A. desideratum separadas entre sí por al menos 5 Km y distribuidas en diferentes áreas ecológicas fueron muestreadas durante la época de floración de 2005, 2006 y 2008. (Tabla 1, Fig. 1)
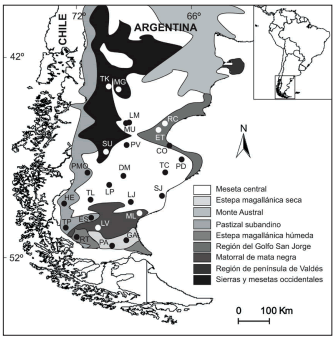
Fig. 1. Ubicación geográfica de las 25 poblaciones estudiadas de Anarthrophyllum desideratum en la Patagonia Austral. En distintos colores se muestran las diferentes áreas ecológicas de la región. Los nombres completos de las poblaciones se proveen en la Tabla 1.
Utilizando tubos capilares de volumen conocido, se extrajo el néctar de tres flores por individuo en 30 individuos por población separados al menos por 5 m a lo largo de una transecta. La extracción del néctar se realizó generalmente en días soleados durante la mañana, en flores nuevas pero no protegidas de las visitas. Esta manera de obtener el néctar, denominada cosecha en pie o"standing crop", proporciona información sobre la cantidad y calidad del néctar disponible en una flor en un momento dado, y depende de la cantidad producida y secretada, menos la cantidad reabsorbida o removida desde el comienzo de la secreción (Corbet, 2003). Esta técnica no afecta la medición de la composición de azúcares. El volumen de néctar de cada flor se obtuvo a partir de la longitud (mm) de la columna de néctar dentro del capilar, la cual fue medida con calibre digital. La concentración del néctar en g/g (g de solutos por 100 g de solución) se midió con un refractómetro de mano (Atago; rango: 0-62%), el cual determina el índice de refracción de una solución en grados BRIX como medida de equivalentes de sacarosa (Dafni, 1992).
La composición de azúcares se determinó mediante cromatografía gaseosa. Se extrajo el néctar de dos a cinco flores por individuo en cinco individuos por población. Las muestras de néctar obtenidas a partir de las diferentes flores de un individuo se agruparon para el análisis cromatográfico. Previamente, el néctar fue liofilizado mediante una bomba de vacío a -80°C para eliminar toda su humedad, luego derivatizado (transformado en derivados volátiles) con hexametildisilasano y trimetilclorosilano en piridina, y finalmente calentado en horno a 75°C durante 15 minutos. Se inyectaron 3μl de cada muestra en un cromatógrafo HP 6890 equipado con un detector de llama ionizada (FID) y una columna Rtx-1. Se utilizó nitrógeno como gas portador y el siguiente programa de temperatura: 145°C por 1 minuto, 8°C/minuto hasta 160°C, 160°C por 5 minutos, 10°C/minuto hasta 225°C, 10°C/minuto hasta 275°C (modificado de Rojas-Escudero et al., 2004). La temperatura del detector fue de 300°C. Los azúcares estándar (glucosa, fructosa y sacarosa) fueron preparados usando el mismo método. En cada muestra, se calculó la proporción de estos tres azúcares como la cantidad (μg/μl) de cada uno de ellos sobre el total de azúcares del néctar.
Para evaluar la variación temporal del néctar, los distintos caracteres fueron medidos durante dos años consecutivos (2005 y 2006) en tres poblaciones: Escarchados (ES), Monte León (ML) y Río Chico (RC) - volumen y concentración -; y ML, Perito Moreno (PM) y San Julián (SJ) - composición de azúcares -. Las plantas muestreadas en los diferentes años no fueron las mismas.
A partir de la base de datos WorldClim (http://www.worldclim.org/; Hijmans et al., 2005) se obtuvieron las temperaturas máxima y mínima y las precipitaciones de los meses de floración de A. desideratum (Octubre y Noviembre) para cada población, y se estimaron los valores promedio para el período de floración (Tabla 1). Estas variables corresponden a los valores medios obtenidos a partir de datos observados entre 1950 y 2000. Además, a partir de la información proporcionada por estaciones meteorológicas del Instituto Nacional de Tecnología Agropecuaria (INTA) se obtuvo la temperatura y precipitación del período de floración de los años de muestreo en sitios cercanos a las localidades estudiadas. Si bien las temperaturas del 2005 y 2006 fueron similares, la época de floración del 2006 fue más seca que la del 2005.
En 16 poblaciones que cubrieron el rango completo de distribución de A. desideratum, se realizaron observaciones directas de los visitantes florales. Cada población fue muestreada intensivamente durante un día, desde las 9.00 a las 19.00 hs. A lo largo de dicho período, se observaron grupos de 8 a 10 plantas durante intervalos regulares de 20 minutos, y se registró el número de plantas visitadas. En total, se realizaron 370 períodos de observación, distribuidos en 123 horas, a lo largo de los tres años de muestreo. Cinco poblaciones fueron muestreadas en dos años consecutivos del estudio, 2005 y 2006. La frecuencia de visitas en cada población se calculó como el número de plantas visitadas dividido por el número de plantas observadas dividido por el tiempo de observación de cada período.
Análisis de datos
Para evaluar la contribución relativa de cada nivel de organización (región ecológica, población, individuo) a la variabilidad espacial total de los distintos caracteres del néctar, se utilizaron modelos lineales mixtos univariados para datos anidados y la significancia estadística de cada nivel se estimó con máxima verosilimiltud restringida (REML) usando el paquete nlme del programa R 2.12.2 (Pinheiro et al., 2009). El volumen y la concentración del néctar fueron particionados de acuerdo a sus diferencias entre áreas ecológicas, entre poblaciones dentro de las áreas ecológicas, entre individuos dentro de las poblaciones y entre flores dentro de los individuos. La proporción de los distintos azúcares se particionó sólo en los tres primeros niveles, dado que el individuo fue la mínima unidad muestreada en este caso. El último nivel de variación en ambos análisis (flor e individuo, respectivamente) fue utilizado como término del error (Sokal & Rohlf, 1981).
Mediante correlaciones de Spearman se evaluó la asociación entre las medias poblacionales de los distintos caracteres del néctar (volumen, concentración y proporción de los diferentes azúcares), y entre éstas, las variables climáticas de la época de floración (temperatura máxima, temperatura mínima, precipitación) y la tasa de visitas de las distintas poblaciones.
Para evaluar la existencia de variación temporal en los caracteres del néctar, y así explorar el potencial efecto de la variación ambiental interanual, se realizaron análisis de Kruskal-Wallis utilizando como factor el año de muestreo (2005 y 2006) en cada una de las tres poblaciones estudiadas para cada rasgo. Mediante este mismo análisis se evaluó si existían diferencias entre años en la tasa de visitas en cada una de las cinco poblaciones muestreadas durante el 2005 y el 2006.
Se utilizaron análisis no paramétricos para evaluar las correlaciones y la variación temporal debido a que los datos no cumplían con los supuestos de normalidad y homogeneidad de la varianza.
RESULTADOS
Las flores de A. desideratum mostraron, en promedio (± error estándar), un volumen de néctar de 8,11 ± 0,13 µl, y una concentración de 27,98 ± 0,39 %. En todas las muestras se identificaron los tres principales azúcares (glucosa, fructosa y sacarosa), y el néctar estuvo dominado por hexosas (glucosa y fructosa), con similares proporciones de glucosa (51,85 ± 0,32 %) y fructosa (46,86 ± 0,38 %), y un porcentaje muy bajo de sacarosa (1,29 ± 0,14 %).
No se encontraron diferencias significativas entre las áreas ecológicas para ninguno de los rasgos estudiados (P > 0,05), mientras que todos los caracteres difirieron significativamente entre poblaciones dentro de las áreas ecológicas (P < 0,05; Fig. 2). Tanto el volumen como la concentración del néctar, cuya variabilidad fue también analizada a nivel intra-poblacional, mostraron diferencias significativas entre individuos dentro de las poblaciones (P < 0,05; Fig. 2). En cuanto a la contribución relativa de cada nivel a la variabilidad total, las diferencias entre áreas ecológicas explicaron un porcentaje muy bajo o nulo de la variación del néctar (Fig. 2). Las diferencias entre poblaciones contribuyeron con la mayor proporción de la varianza explicada en todos los caracteres analizados, representando entre el 29% y el 52% de la varianza total según el rasgo considerado (Fig. 2). El volumen de néctar de las poblaciones varió entre menos de 1 µl y 16 µl, la concentración, entre 9% y 53%, mientras que la proporción de hexosas, sólo lo hizo entre 95% y 99%. El porcentaje de varianza no explicado (error) fue en general elevado, especialmente para la proporción de los tres principales azúcares, lo que sugiere una importante variación en la composición del néctar entre individuos dentro de las poblaciones.

Fig. 2. Componentes de varianza, expresados como porcentajes de la varianza total, entreáreas ecológicas, entre poblaciones dentro de lasáreas ecológicas, entre individuos dentro de las poblaciones, y entre flores dentro de los individuos. El término del error corresponde a las diferencias entre flores dentro de los individuos para el volumen y la concentración de néctar, y a las diferencias entre individuos dentro de las poblaciones para los porcentajes de glucosa, fructosa y sacarosa. La significancia estadística de cada componente se muestra dentro de las barras. *** P < 0,0001; ** P < 0,001; * P < 0,05; n.s. = no signifcativo.
La composición de azúcares de las diferentes poblaciones no mostró un patrón claro en relación a las áreas ecológicas de la estepa Patagónica (Fig. 3).

Fig. 3. Diagrama ternario que muestra los porcentajes de azúcares (fructosa, glucosa y sacarosa) del néctar de 25 poblaciones de Anarthrophyllum desideratum en la Patagonia Austral. Con distintos símbolos se muestran las diferentes áreas ecológicas que ocupa cada población.
Los atributos del néctar de las distintas poblaciones estuvieron en general significativamente correlacionados (Tabla 2). El volumen y la concentración se correlacionaron de manera positiva aunque marginalmente significativa, y ambos caracteres se asociaron de manera negativa con la proporción de fructosa y de manera positiva (aunque sólo marginalmente en el caso de la concentración) con el porcentaje de glucosa del néctar. La concentración se correlacionó de manera positiva con la proporción de sacarosa y de manera negativa con la proporción de hexosas (glucosa + fructosa), mientras que el volumen no mostró una relación significativa con estas variables. Finalmente, la proporción de fructosa de las poblaciones se correlacionó negativamente con la proporción de los otros dos azúcares (glucosa y sacarosa), los cuales estuvieron positivamente correlacionados entre sí.
Tabla 2. Correlaciones de Spearman entre los caracteres del néctar floral en las poblaciones estudiadas de Anarthrophyllum desideratum.

† P < 0,1; * P < 0,05; ** P < 0,001; *** P < 0,0001.
Las correlaciones con las variables climáticas mostraron que, en general, las características del néctar de las poblaciones no se asociaron de manera significativa con las temperaturas (máxima y mínima) ni con las precipitaciones de la época de floración (P > 0,05). Sólo la proporción de glucosa se correlacionó negativamente y de manera marginalmente significativa con la temperatura mínima de dicho período (r = -0,37; P = 0,07). Por su parte, el volumen del néctar fue el único rasgo que se correlacionó, aunque de manera marginalmente significativa, con la tasa de visitas de las diferentes poblaciones, mostrando una disminución a medida que aumentó la frecuencia de visitas (r = -0,49; P = 0,06).
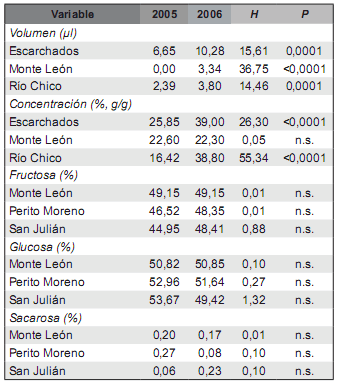
El volumen de néctar difirió significativamente entre los dos años consecutivos de muestreo (2005 y 2006) en las tres poblaciones estudiadas (Escarchados, Monte León y Río Chico), y la concentración, en dos de ellas (Escarchados y Río Chico) (Tabla 3). Tanto el volumen como la concentración del néctar fueron mayores en el 2006 que en el 2005 en dichas poblaciones. Por su parte, la proporción de los distintos azúcares no difirió entre los dos períodos muestreados en ninguna de las tres poblaciones estudiadas (Monte León, Perito Moreno y San Julián) (Tabla 3). La tasa de visitas de las cinco poblaciones muestreadas en el 2005 y el 2006 (incluida Río Chico) no difirió significativamente entre años (P > 0,05).
Tabla 3. Resultados de los análisis no paramétricos de la varianza (Kruskal-Wallis) para los caracteres del néctar de Anarthrophyllum desideratum entre dos años de muestreo consecutivos (2005 y 2006) en tres poblaciones en cada caso. Se reportan las medianas para los diferentes años muestreados.
DISCUSIÓN
Caracterización del néctar de A. desideratum
Los datos presentados en este trabajo constituyen los primeros registros de néctar para Anarthrophyllum desideratum. En promedio, las flores produjeron una copiosa cantidad de néctar (8 µl) de baja concentración (28%). El elevado volumen de néctar de las flores de esta especie coincide con lo hallado en otras especies polinizadas por aves (Faegri & van der Pijl, 1979; Proctor et al., 1996; Fenster et al., 2004), y podría responder a los altos requerimientos energéticos de estos polinizadores de mayor tamaño en comparación con polinizadores más pequeños como insectos. Por su parte, la concentración de A. desideratum fue menor que la observada en la especie co-distribuida del mismo género polinizada por abejas, Anarthrophyllum rigidum (H. & A.) Hieron. (60%, Forcone et al., 1997), y en otras especies entomófilas de la estepa (45% en promedio, Forcone et al., 1997, y 42% en promedio, Bernardello et al., 1999); mientras que fue similar a las registradas en especies polinizadas por aves del bosque templado Andino-Patagónico (29% en promedio, Chalcoff et al., 2006). Sin embargo, la concentración del néctar de A. desideratum fue mayor que la de otras plantas polinizadas por aves Passeriformes a latitudes menores que las Patagónicas (5-15%), en Sudamérica (Bernardello et al., 1994; Rocca & Sazima, 2010; Agostini et al., 2011) y otras partes del mundo (Nicolson, 2002). Esto quizás se deba al mayor requerimiento energético de los polinizadores que viven a bajas temperaturas, como las de la estepa Patagónica, donde la recompensa alimenticia debería brindar un gran aporte energético (Heinrich & Raven, 1972). Diferentes hipótesis han sido postuladas para explicar la presencia de néctares diluidos en las flores ornitófilas: 1) su extracción por parte de las aves es más eficiente (Baker & Baker, 1975); 2) evita a las abejas, las cuales constituyen polinizadores menos efectivos que prefieren néctares muy concentrados (Bolten & Feinsinger, 1978); 3) además de energía, provee suficiente cantidad de agua a las aves (Baker, 1975; Calder, 1979); y 4) fomenta las visitas de las aves, ya que el consumo de energía a partir de unas pocas visitas no es suficiente (por ej., Martínez del Rio et al., 2001). Sin embargo, algunos trabajos indican que las aves no eligen exclusivamente néctares diluidos, sino el máximo volumen de néctar disponible independientemente de la concentración, siendo el criterio energético el determinante más importante en la elección de flores por parte de las aves (Bolten & Feinsinger, 1978; Tamm & Gass, 1986; Mitchell & Paton, 1990). Este criterio podría ser clave en las aves que visitan las flores de A. desideratum, las cuales no son nectarívoras, sino principalmente granívoras o insectívoras y ocasionalmente frugívoras (Montaldo N. H., com. pers.), y parecen comportarse como oportunistas aprovechando la fuente energética del néctar en una época del año donde quizás no hay tanta disponibilidad de sus alimentos habituales. En este sentido, tanto el gran volumen de néctar como la numerosa cantidad de flores por planta (440 en promedio, Paiaro, 2011) de A. desideratum la convierten en un sistema altamente atractivo para estas especies de aves en la estepa Patagónica.
En cuanto a la composición de azúcares, tal como fuera postulado para las especies polinizadas por Passeriformes (Baker & Baker, 1983b; Proctor et al., 1996; Nicolson, 2002), el néctar de A. desideratum estuvo dominado por hexosas (glucosa y fructosa), mientras que presentó un porcentaje muy bajo de sacarosa (ca. 1%). Este resultado coincide con lo hallado en poblaciones de la especie Embothrium coccineum J.R. Forst.& G. Forst. polinizadas por aves Passeriformes en los Bosques Andino-Patagónicos de Chile (Smith-Ramírez & Armesto, 1998; Chalcoff et al., 2006). La dominancia de hexosas en el néctar de las especies polinizadas por Passeriformes podría estar asociada a la ausencia de la enzima intestinal responsable de la hidrólisis de sacarosa en glucosa y fructosa en este grupo de aves, que sólo sería capaz de digerir azúcares simples como las hexosas (Schondube & Martínez del Rio, 2004). Alternativamente, la ocurrencia de néctar rico en hexosas en A. desideratum podría tener una base filogenética (Baker & Baker, 1982), ya que su composición fue similar a la de otras Fabáceas de las regiones Chaqueña y Patagónica, y particularmente a la de la especie co-distribuida del mismo género, A. rigidum (Forcone et al., 1997; Bernardello et al., 1999; Galetto & Bernardello, 2003).
Variabilidad espacio-temporal del néctar
Si bien los atributos del néctar floral suelen ser característicos de una especie de planta con un determinado síndrome floral, la variabilidad espacial y temporal encontrada dentro de A. desideratum así como las relaciones entre los distintos rasgos de las poblaciones sugieren que las características del néctar varían a nivel intra-específico dentro de un mismo grupo funcional de polinizadores. La asociación positiva entre la concentración y el contenido de sacarosa y la asociación negativa entre la concentración y la proporción de hexosas (particularmente fructosa) de las poblaciones podrían responder tanto a procesos osmóticos en las plantas (Nicolson, 2002) como a presiones selectivas ejercidas por los Passeriformes con preferencias dependientes de la concentración (Schondube & Martínez del Rio, 2003). Sin embargo, es poco probable que las aves que visitan las flores de esta especie de manera oportunista tengan requerimientos específicos en cuanto al néctar que sean seleccionados diferencialmente en las distintas poblaciones. Por otro lado, contrariamente a lo esperado según la hipótesis de Nicolson (2002), la proporción de sacarosa y el volumen del néctar en las poblaciones no estuvieron significativamente correlacionados, y el volumen y la concentración estuvieron positivamente asociados. Esta última asociación podría deberse a que cuando aumenta la concentración del néctar, las aves nectarívoras disminuyen el volumen de la ingesta (Martínez del Río et al., 2001 y referencias allí citadas), lo que podría resultar en un incremento de la cosecha en pie con la concentración de néctar.
La variación geográfica en los parámetros del néctar de A. desideratum podrían responder a los gradientes climáticos presentes en la estepa Patagónica (Bertsch, 1983; Freeman & Head, 1990; Devoto et al., 2006), especialmente tratándose de una especie de flores abiertas con néctar expuesto (Corbet et al., 1979; Nicolson, 2002). La ausencia de diferencias significativas en los caracteres del néctar entre las áreas ecológicas de la estepa indica que las variaciones macro-ambientales en clima, suelo y vegetación que diferencian estasáreas no serían un factor modelador importante del néctar floral en esta especie. Por el contrario, las importantes diferencias encontradas entre las poblaciones y el hecho de que éstas explicaran la mayor parte de la variabilidad espacial en todos los caracteres analizados sugieren que variaciones ambientales que ocurren a escala local afectando a las poblaciones podrían ser las responsables de la variación del néctar. En efecto, el porcentaje de glucosa del néctar aumentó a medida que descendió la temperatura de los sitios, contrariamente al patrón encontrado para Ipomopsis longiflora, donde la proporción de hexosas aumentó con la temperatura (Villarreal & Freeman, 1990). El mayor contenido de glucosa en el néctar de A. desideratum a menor temperatura podría deberse a un incremento de la actividad enzimática de hidrólisis de la sacarosa en el nectario con un consecuente aumento en la cantidad de hexosas hacia los sitios más fríos como resultado del estrés por bajas temperaturas, y no por altas temperaturas como se halló en I. longiflora (Villarreal & Freeman, 1990). Alternativamente, el patrón encontrado podría responder a los mayores requerimientos energéticos de las aves polinizadoras en ambientes más fríos (Calder, 1979). Por su parte, ni el volumen ni la concentración del néctar mostraron una asociación significativa con la temperatura o las precipitaciones en las distintas poblaciones, sugiriendo que estos rasgos no responderían a gradientes ambientales simples, a diferencia de lo que sucede con otros caracteres florales de la especie, como el color, el tamaño y la forma de la corola (Paiaro et al., en prensa). Nuestros resultados también difieren de lo encontrado para Embothrium coccineum en los Bosques Andino- Patagónicos, donde la concentración del néctar aumentó con el gradiente de aridez (Devoto et al., 2006). La variabilidad tanto inter- como intra-poblacional hallada en el volumen y la concentración del néctar de A. desideratum podrían ser el resultado de variaciones en la actividad de los polinizadores, ya que el método usado en la recolección de muestras ("standing crop") refleja la oferta natural de néctar, es decir, la cantidad producida menos la cantidad removida por los visitantes florales (Corbet, 2003). Así, se espera que la cosecha en pie acumulada en flores no visitadas sea mucho mayor que la de flores con una alta frecuencia de visitas, y que las trazas de néctar en flores recientemente visitadas se concentren mucho más rápido que los volúmenes más grandes de las flores no visitadas (Corbet, 2003). En el presente estudio, sólo el volumen de néctar disminuyó a medida que aumentó la frecuencia de visitas de las aves Passeriformes en las poblaciones, sugiriendo que la variación en la actividad de estos visitantes florales dentro del rango geográfico de la especie podría afectar la cantidad de néctar de las poblaciones, mientras que no contribuiría a explicar las diferencias en su concentración. Alternativamente, las variaciones encontradas en el volumen del néctar podrían ser consecuencia de una relación alométrica con el tamaño del nectario, ya que flores con nectarios más grandes producen más cantidad de néctar (Petanidou et al., 2000). El incremento en el volumen de néctar con el tamaño de las flores reportado para las poblaciones de A. desideratum (Paiaro, 2011) apoyaría esta hipótesis. A su vez, nectarios de mayor tamaño también podrían ser seleccionados indirectamente por los polinizadores a través de la selección de grandes volúmenes de néctar (Corbet et al., 1984), aunque el mayor volumen de néctar en las poblaciones menos visitadas descartaría esta idea.
Si bien el volumen y la concentración del néctar de las poblaciones variaron temporalmente entre años consecutivos, la proporción de los distintos azúcares fue constante en las tres poblaciones estudiadas, sugiriendo que la composición sería un carácter más conservativo que el volumen y la concentración. En parte, esto podría deberse a que la técnica de standing crop afecta los valores de volumen y concentración del néctar pero no la composición de azúcares. También se ha reportado que la composición de azúcares del néctar es menos afectada por el ambiente que la concentración y el volumen (Baker & Baker, 1982). Nuestros resultados difieren de lo hallado en Ipomopsis longifora, donde la composición de carbohidratos del néctar, y en particular el porcentaje de sacarosa, dentro de las poblaciones mostró variaciones a lo largo de la estación en asociación con los cambios de temperatura (Freeman & Head, 1990 y referencias allí citadas). Las diferencias interanuales en las precipitaciones y/o en la actividad de los polinizadores en los sitios estudiados podrían explicar las variaciones temporales halladas en el volumen y la concentración del néctar. En este sentido, el aumento en la evaporación en el año más seco (2006) podría haber estimulado la producción y secreción de néctar, llevando a un incremento del volumen y la concentración respecto del año más húmedo (2005) (Corbet et al., 1979; Devoto et al., 2006). Además, se ha sugerido que la sequía limitaría la actividad de los polinizadores de A. desideratum (Paiaro, 2011), lo que también podría contribuir al incremento de la cosecha en pie de las poblaciones en el año más seco. Sin embargo, la falta de diferencias significativas en la tasa de visitas entre los años 2005 y 2006 no apoyaría esta hipótesis. Adicionalmente, deben considerarse como causas alternativas las posibles diferencias en el momento de la floración en que los datos fueron tomados cada año.
CONCLUSIONES
Los resultados de este trabajo señalan que el volumen, la concentración y la composición de azúcares del néctar de A. desideratum estarían estrechamente asociados al síndrome floral de la especie, y podrían haber evolucionado en respuesta al grupo funcional de las aves Passeriformes. No obstante, las relaciones entre los distintos caracteres y la importante variación espacial y temporal observada dentro de esta especie sugieren que el grupo funcional de polinizadores no es la única fuerza que moldea los caracteres del néctar, los cuales estarían bajo el efecto de otros factores que actúan a nivel intra-específico. La composición de azúcares del néctar estaría afectada por los cambios de temperatura dentro del rango geográfico de la especie, mientras que las variaciones inter-poblacionales en el volumen y la concentración no responderían a los gradientes climáticos de la estepa Patagónica. Las diferencias entre poblaciones en el volumen estarían asociadas a la actividad de los visitantes florales. Sin embargo, la tasa de visitas de los polinizadores no explicaría las variaciones inter-anuales en la cantidad y calidad de la recompensa floral, las cuales podrían responder a las diferencias en las precipitaciones entre los años de muestreo. Nuestros resultados ponen en evidencia la multiplicidad y complejidad de factores que pueden actuar sobre los caracteres del néctar floral dando lugar a la variabilidad espacio-temporal observada. A su vez, estas variaciones intra-específicas pueden influenciar el comportamiento de los polinizadores (frecuencia de visitas, número de flores visitadas, etc.), determinando las probabilidades de cruzamiento y el éxito reproductivo de las plantas (Rathcke, 1992). Futuros estudios deberían evaluar el rol que juega la variabilidad del néctar sobre la actividad de los polinizadores en las diferentes poblaciones de A. desideratum, y si dicha variación se refeja en un éxito reproductivo diferencial.
AGRADECIMIENTOS
Agradecemos a la Dra. A. Cosacov, G. Humano, A. Mazzoni, C. Franchini, R. Kofalt, C. Lazarte y C. Romanutti por su colaboración en los muestreos. Al Dr. G. Bernardello por el asesoramiento acerca de la conservación de las muestras. A la Dra. M. A. Perillo por facilitar el uso de equipamiento, y a P. Clop por su ayuda en la lioflización de las muestras. Al Dr. L. Johnson (Brigham Young University) por proporcionar el material utilizado en las cromatografías gaseosas, y al Dr. J. Zou por su asesoramiento en esta técnica. A APN Argentina y CONAF Chile por permitir realizar los muestreos en sitios localizados en parques y reservas. A los dos revisores anónimos que han contribuido sustancialmente a mejorar la calidad de este trabajo. Este estudio fue realizado en el marco de la Tesis Doctoral de V. Paiaro en el Doctorado de Ciencias Biológicas de la Universidad Nacional del Córdoba, y fue financiado por FONCyT, CONICET y NSF-PIRE. Actualmente, V.P. es becaria posdoctoral del CONICET, y A.A.C. y A.N.S. son investigadores de la misma Institución.
BIBLIOGRAFÍA
1. AGOSTINI, K., M. SAZIMA & L. GALETTO. 2011. Nectar production dynamics and sugar composition in two Mucuna species (Leguminosae, Faboideae) with different specialized pollinators. Naturwissenschaften 98: 933-942. [ Links ]
2. BAKER, H. G. 1975. Sugar concentrations in nectars from hummingbird flowers. Biotropica 7: 37-41. [ Links ]
3. BAKER H. G. & I. BAKER. 1975. Studies of nectar- constitution and pollinator-plant coevolution. In: GILBERT, L. E. & P. H. RAVEN (eds.), Coevolution of animals and plants, pp. 100-140. University of Texas Press, Austin. [ Links ]
4. BAKER, H. G. & I. BAKER. 1982. Chemical constituents of nectar in relation to pollination mechanisms and phylogeny. In: NITECKI, M. H. (ed.), Biochemical Aspects of Evolutionary Biology, pp. 131-171. University of Chicago Press, Chicago. [ Links ]
5. BAKER, H. G. & I. BAKER. 1983a. A brief historical review of the chemistry of floral nectar. In: BENTLEY, B. & T. S. ELIAS (eds.), The biology of nectaries, pp. 127-152. Columbia University Press, New York. [ Links ]
6. BAKER, H. G. & I. BAKER. 1983b. Floral nectar sugar constituents in relation to pollinator type. In: JONES, C. E. & R. J. LITTLE (eds.), Handbook of experimental pollination biology, pp.117-141. Van Nostrand Reinhold, New York. [ Links ]
7. BAKER, H. G., I. BAKER & S. A. HODGES. 1998. Sugar composition of nectars and fruits consumed by birds and bats in the tropics and subtropics. Biotropica 30: 559-586. [ Links ]
8. BERNARDELLO, G., L. GALETTO & A. FORCONE. 1999. Floral nectar chemical composition of some species from Patagonia II. Biochem. Syst. Ecol. 27: 779-790. [ Links ]
9. BERNARDELLO, L., L. GALETTO & I. G. RODRIGUEZ. 1994. Reproductive biology, variability of nectar features and pollination of Combretum fruticosum (Combretaceae) in Argentina. Bot. J. Linn. Soc. 114: 293-308. [ Links ]
10. BERTSCH, A. 1983. Nectar production of Epilobium angustifolium L. at different air humidities: nectar sugar in individual flowers and the optimal foraging theory. Oecologia 59: 40-48. [ Links ]
11. BOLTEN, A. B. & P. FEINSINGER. 1978. Why do hummingbird flowers secrete dilute nectar? Biotropica 10: 307-309. [ Links ]
12. BRINK, D. W. & J. M. J. DEWET. 1980. Interpopulation variation in nectar production in Aconitum columbianum (Ranunculaceae). Oecologia 47: 160-163. [ Links ]
13. BROWN, J. H. & A. KODRIC-BROWN. 1979. Convergence, competition, and mimicry in a temperate community of hummingbird pollinated flowers. Ecology 60: 1022-1035. [ Links ]
14. CALDER, W. A. 1979. On the temperature dependency of optimal nectar concentration for birds. J. Theor. Biol. 78: 185-196. [ Links ]
15. CHALCOFF, V. R., M. A. AIZEN & L. GALETTO. 2006. Nectar concentration and composition in 26 species from the temperate forest of South America. Ann. Bot. 97: 413-421. [ Links ]
16. CORBET, S. A. 2003. Nectar sugar content: estimating standing crop and secretion rate in the field. Apidologie 34: 1-10. [ Links ]
17. CORBET, S. A., C. J. C. KERSLAKE, D. BROWN & N. E. MORLAND. 1984. Can bees select nectar-rich fowers in a patch? J. Apic. Res. 23: 234-242. [ Links ]
18. CORBET, S. A., P. G. WILLMER, J. W. L. BEAMENT, D. M. UNWIN & O. E. PRŷS-JONES. 1979. Post- secretory determinants of sugar concentration in nectar. Plant Cell Environ. 2: 293-308. [ Links ]
19. CRUDEN, R. W., S. M. HERMANN-PARKER & S. PETERSON. 1983. Patterns of nectar production and plant pollinator co-evolution. In: BENTLEY, B. & T. ELIAS (eds.), Biology of nectaries, pp. 80-125. Columbia University Press, New York. [ Links ]
20. DAFNI, A. 1992. Pollination Ecology. A practical approach. Oxford University Press, Oxford. [ Links ]
21. DEVOTO, M., N. H. MONTALDO & D. MEDAN. 2006. Mixed hummingbird: long-proboscid-fly pollination in 'ornithophilous' Embothrium coccineum (Proteaceae) along a rainfall gradient in Patagonia, Argentina. Austral Ecol. 31: 512-519. [ Links ]
22. DUPONT, Y. L., D. M. HANSEN, J. T. RASMUSSEN & J. M. OLESEN. 2004. Evolutionary changes in nectar sugar composition associated with switches between bird and insect pollination: the Canarian bird-flower element revisited. Funct. Ecol. 18: 670-676. [ Links ]
23. FAEGRI, K. & L. VAN DER PIJL. 1979. The principles of pollination ecology. Pergamon Press, New York. [ Links ]
24. FENSTER, C. B., W. S. ARMBRUSTER, P. WILSON, M. R. DUDASH & J. D. THOMSON. 2004. Pollination syndromes and floral specialization. Ann. Rev. Ecol., Evol. Syst. 35: 375-403. [ Links ]
25. FORCONE A., L. GALETTO & L. BERNARDELLO. 1997. Floral nectar chemical composition of some species from Patagonia. Biochem. Syst. Ecol. 25: 395-402. [ Links ]
26. FREEMAN, C. E. & K. C. HEAD. 1990. Temperature and sucrose composition of floral nectars in Ipomopsis longifora under field conditions. Southwest. Nat. 35: 432-426. [ Links ]
27. GALETTO, L. & G. BERNARDELLO. 2003. Sugar nectar composition in angiosperms from Chaco and Patagonia (Argentina): animal visitor's matter? Plant Syst. Evol. 238: 69-86. [ Links ]
28. HEINRICH, B. & P. H. RAVEN. 1972. Energetics and pollination ecology. Science 176: 597-602. [ Links ]
29. HERRERA, C. M. 1995. Microclimate and individual variation in pollinators: Flowering plants are more than their flowers. Ecology 76: 1516-1524. [ Links ]
30. HIJMANS R. J., S. E. CAMERON, J. L. PARRA, P. G. JONES & A. JARVIS. 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25: 1965-1978. [ Links ]
31. LANZA, J., G. C. SMITH, S. SACK & A. CASH. 1995. Variation in nectar volume and composition of Impatiens capensis at the individual, plant, and population levels. Oecologia 102: 113-119. [ Links ]
32. MARTíNEZ DEL RIO, C., J. E. SCHONDUBE, T. J. MCWHORTER & L. G. HERRERA. 2001. Intake responses in nectar feeding birds: Digestive and metabolic causes, osmoregulatory consequences, and coevolutionary effects. Amer. Zool.41: 902-915. [ Links ]
33. MITCHELL, R. J. & D. C. PATON. 1990. Effects of nectar volume and concentration on sugar intakerates of Australian honeyeaters (Meliphagidae). Oecologia 82: 238-246. [ Links ]
34. NICOLSON, S. W. 2002. Pollination by passerine birds: why are the nectars so dilute? Comp. Biochem. Phys. B 131: 645-652. [ Links ]
35. OLIVA, G. E., L. GONZÁLEZ, P. RIAL & E. LIVRAGHI. 2001. El ambiente en la Patagonia Austral. En: BORRELLI, P. & G. E. OLIVA (eds.), Ganadería sustentable en la Patagonia Austral, pp. 41-82. INTA, Río Gallegos. [ Links ]
36. PAIARO, V. 2011. Gradientes ambientales y márgenes de distribución: Patrones espaciales de variabilidad fenotípica, atributos poblacionales y caracteres reproductivos en Anarthrophyllum desideratum (DC) Benth., Universidad Nacional de Córdoba, Argentina. [ Links ]
37. PAIARO, V., G. E. OLIVA, A. A. COCUCCI & A. N. SÉRSIC. En prensa. Geographic patterns and environmental drivers of flower and leaf variation in an endemic legume of Southern Patagonia. Pl. Ecol. Diver. DOI:10.1080/17550874.2012.713403. [ Links ]
38. PATE, J. S., M. B. PEOPLES, P. J. STORER & C. A. ATKINS. 1985. The extrafloral nectaries of cowpea (Vigna unguiculata (L.) Walp.). II. Nectar composition, origin of nectar solutes, and nectar functioning. Planta 166: 28-38. [ Links ]
39. PERCIVAL, M. S. 1961. Types of nectars in angiosperms. New Phytol. 60: 235-281. [ Links ]
40. PETANIDOU, T., V. GOETHALS, E. SMETS. 2000. Nectary structure of Labiatae in relation to their nectar secretion and characteristics in a Mediterranean shrub community - Does flowering time matter? Pl. Syst. Evol. 225:103-118. [ Links ]
41. PINHEIRO, J., D. BATES, S. DEBROY, D. SARKAR and the R Core team 2009. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3, pp. 1-96. [ Links ]
42. PLEASANTS, J. M. 1981. Bumblebee responses to variation in nectar availability. Ecology 62:1648-1661. [ Links ]
43. PLEASANTS, J. M. 1983. Nectar production patterns in Ipomopsis aggregate (Polemoniaceae). Amer. J. Bot. 70: 1468-1475. [ Links ]
44. PLEASANTS, J. M. & S. J. CHAPLIN. 1983. Nectar production rates of Asclepias quadrifolia: causes and consequences of individual variation. Oecologia 59: 232-238. [ Links ]
45. PROCTOR, M. P., P. YEO & A. LACK. 1996. The natural history of pollination. Oregon Timber Press, Portland. [ Links ]
46. PYKE, G. H. & N. M. WASER. 1981. The production of dilute nectars by hummingbird and honeyeater flowers. Biotropica 13: 260-270. [ Links ]
47. RATHCKE, B. J. 1992. Nectar distributions, pollinator behavior, and plant reproductive success. In: HUNTER, M. D., T. OHGUSHI & P. W. PRICE (eds.), Effects of resource distribution on animal-plant interactions, pp. 113-138. Academic Press, New York. [ Links ]
48. ROCCA, M. A. & M. SAZIMA. 2010. Beyond hummingbird-flowers: The other side of ornithophily in the Neotropics. Oecologia Australis 14: 67-99. [ Links ]
49. ROIG, F. A. 1998. La vegetación de la Patagonia. In: CORREA, M. N. (ed.), Flora Patagónica. Vol 8, Parte 1, pp. 48-166. INTA. [ Links ]
50. ROJAS-ESCUDERO, E., A. L. ALARCÓN-JIMÉNEZ, P. ELIZALDE-GALVÁN & F. ROJO-CALLEJAS. 2004. Optimization of carbohydrate silylation for gas chromatography. J. Chromatogr. A 1027: 117-120. [ Links ]
51. SCHONDUBE, J. E. & C. MARTÍNEZ DEL RIO. 2004. Sugar and protein digestion in flowerpiercers and hummingbirds: a comparative test of adaptive convergence. J. Comp. Physiol. B 174: 263-273. [ Links ]
52. SCHONDUBE, J. E. & C. MARTÍNEZ DEL RIO. 2003. Concentration-dependent sugar preferences in nectar-feeding birds: mechanisms and consequences. Funct. Ecol. 17: 445-453. [ Links ]
53. SIMPSON, B. B. & J. L. NEFF. 1983. Evolution and diversity of floral rewards. En: JONES, C. E. & R. J. LITTLE (eds.), Handbook of experimental pollination biology, págs. 142-159. Van Nostrand Reinhold, New York. [ Links ]
54. SMITH-RAMÍREZ, C. & J. J. ARMESTO. 1998. Nectarivoría y polinización por aves en Embothrium coccineum (Proteaceae) en el bosque templado del sur de Chile. Rev. Chil. Hist. Nat. 71: 53-65. [ Links ]
55. SOKAL, R. R. & F. J. ROHLF. 1981. Biometry. Freeman. [ Links ]
56. SORARÚ, S. 1974. Revisión de Anarthrophyllum, género argentino-chileno de Leguminosas. Darwiniana 18: 453-488. [ Links ]
57. TAMM, S. & C. L. GASS. 1986. Energy intake rates and nectar concentration preferences by hummingbirds. Oecologia 702: 20-23. [ Links ]
58. VILLARREAL, A. G. & C. E. FREEMAN. 1990. Effects of temperature and water stress on some floral nectar characteristics in Ipomopsis longifora (Polemoniaceae) under controlled conditions. Bot. Gaz. 151: 5-9. [ Links ]
Recibido el 15 de diciembre de 2011
Aceptado el 25 de octubre de 2012.

