










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
On-line version ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.48 no.1 Córdoba Jun./May 2013
FICOLOGÍA
Diatomeas de ambientes marinos costeros de Argentina. Especies del género Pleurosigma (Pleurosigmataceae)
Eugenia A. Sar1,2*, Frithjof A.S. Sterrenburg3, Andrea S. Lavigne1 e Inés Sunesen1,2
1 División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina.
2 Consejo Nacional de Investigaciones Científicas y Técnicas, CONICET, Argentina.
3 Stationsweg 158, 1852LN Heiloo, The Netherlands.
*easar@fcnym.unlp.edu.ar
Resumen: Este estudio está enfocado en el análisis de la morfología, taxonomía y distribución de las especies pertenecientes al género de diatomeas Pleurosigma W. Smith presentes en aguas marinas costeras de la Argentina. Las muestras de red fueron obtenidas en la capa superficial de la columna de agua en varias estaciones costeras de las provincias de Buenos Aires y Río Negro. Materiales procesados y no procesados fueron examinados con microscopía óptica y electrónica de barrido. Las especies determinadas fueron Pleurosigma decorum W. Smith, P. distinguendum Hustedt, P. inflatum Shadbolt, P. inscriptura M. A. Harper, P. normanii Ralfs y P. simonsenii Hasle. Dos nuevos taxa son erigidos. Pleurosigma patagonicum var. paucistriatum nov. var. es comparada con el material tipo de Pleurosigma patagonicum (Ferrario & Sar) Sterrenburg & Sar var. patagonicum y P. perlucidum nov. spec. es comparada con P. elongatum W. Smith, P. delicatulum W. Smith, P. amara Stidolph y P. brunii H. Peragallo sobre la base del análisis de materiales tipo y/o protologos. Nuevas observaciones sobre Pleurosigma decorum, P. distinguendum, P. inflatum, P. inscriptura y P. normanii son incluidas y las diferencias con especies relacionadas son discutidas. Pleurosigma inflatum es citada por primera vez para Argentina y P. distinguendum y P. inscriptura son citadas por primera vez para el Océano Atlántico Sur.
Palabras clave: Aguas marinas costeras; Pleurosigma; Provincia de Buenos Aires; Provincia de Río Negro; Argentina; Taxonomía; Morfología.
Summary: Diatoms from marine coastal environments of Argentina. Species of the genus Pleurosigma (Pleurosigmataceae). This study focuses on the analysis of the morphology, taxonomy and distribution of species belonging to the diatom genus Pleurosigma W. Smith found in marine coastal waters of Argentina. Net samples were collected from the surface layer of the water column at several coastal stations in Buenos Aires and Río Negro Provinces. Unprocessed and cleaned materials were examined by light and scanning electron microscopy. Species identified were Pleurosigma decorum W. Smith, P. distinguendum Hustedt, P. inflatum Shadbolt, P. inscriptura M. A. Harper, P. normanii Ralfs and P. simonsenii Hasle. Two new taxa are described. Pleurosigma patagonicum var. paucistriatum nov. var. is compared with type material of Pleurosigma patagonicum (Ferrario & Sar) Sterrenburg & Sar var. patagonicum and P. perlucidum nov. spec. is compared with P. elongatum W. Smith, P. delicatulum W. Smith, P. amara Stidolph and P. brunii H. Peragallo based on type materials and/or protologues. New observations on Pleurosigma decorum, P. distinguendum, P. inflatum, P. inscriptura and P. normanii are included and differences from related species are discussed. P. inflatum is recorded for the first time from Argentina and Pleurosigma distinguendum and P. inscriptura are recorded for first time from the South Atlantic Ocean.
Key words: Marine coastal waters; Pleurosigma; Buenos Aires Province; Río Negro Province; Argentina; Taxonomy; Morphology.
Introducción
Pleurosigma es un género de diatomeas que tiene un grupo grande de especies bentónicas, un grupo pequeño de especies planctónicas y aparece frecuentemente en ambientes marinos neríticos de todo el mundo. Pleurosigma W. Smith es un nombre conservado y el género presenta una compleja historia taxonómica que fue analizada en detalle por Sar et al. (2012). De acuerdo con VanLandingham (1978) Pleurosigma W. Smith sensu stricto contiene noventa taxa válidamente publicados y de acuerdo con Reid (2012) contiene 306 especies. Este género ha sido caracterizado por presentar estrías oblicuas y contorno valvar y rafe-sternum sigmoideos, sin embargo también incluye especies estrechamente lanceoladas o rómbico lanceoladas rectas o casi rectas con rafesternum recto o ligeramente excéntrico hacia los ápices. A partir de los trabajos de Cardinal et al. (1989) y Sterrenburg (1991a, b) en que se discuten los caracteres diagnósticos para separar especies de Pleurosigma varias investigaciones críticas de materiales tipo de diversas especies han ayudado a clarificar algunos problemas taxonómicos (e.g. Sterrenburg 1991b, 2001, 2003, Reid 2001, 2002, Stidolph 2002, Sar et al. 2012, Sterrenburg et al. enviado). Sin embargo, el género requiere aún de estudios morfológicos que incluyan microscopía electrónica, que han sido bastante escasos y/o fragmentarios hasta el presente.
Los primeros estudios sobre diatomeas de ambientes marinos costeros de las Provincias de Buenos Aires y Río Negro fueron realizados por Frenguelli (1928, 1930, 1938, 1939), posteriormente en las últimas tres décadas se realizaron varios estudios ultraestructurales de diferentes grupos de diatomeas (i.e. Lange et al. 1983, Gayoso 1988, 1989, Ferrario et al. 1999, Negri & Inza 1998, Sar 1989,1996a, b, Sar & Ferrario 1990, Sar & Sunesen 2003, Sar et al. 2001, 2002a, b, 2003, 2007a, b, 2010a, b, 2011, Sunesen & Sar 2004, 2007a, b y Sunesen et al. 2008, 2009). Sin embargo, excepto por el trabajo de Ferrario & Sar (1990) sobre un taxa puntual de Pleurosigma realizado en base a material procedente de Argentina, el género no ha recibido atención hasta el presente. Según Vouilloud (2003) veintiún taxa de Pleurosigma fueron citados para el país en listas o en trabajos florísticos sin análisis morfológico.
El objetivo del presente estudio es analizar las especies de Pleurosigma halladas en el área de estudio y proveer detalles sobre su morfología, taxonomía y distribución.
Materiales y Métodos
El material estudiado en el presente trabajo fue recolectado en tres áreas marinas costeras disjuntas (Fig. 1):

Fig. 1. Mapa del área de estudio y su localización en Argentina. Estaciones de muestreo, 1: San Clemente del Tuyú, 2: Santa Teresita, 3: Mar del Tuyú, 4: La Lucila del Mar; 5: Mar de Ajó, 6: Nueva Atlantis, 7: Pinamar, 8: Villa Gesell, 9: Mar Azul, 10: Los Pocitos, 11: Ría del Jabalí, 12: San Blas, 13: Punta Orengo, 14: Las Garzas, 15: San Antonio Oeste, 16: Banco Reparo, 17: Los Alamos, 18: Las Grutas, 19: Piedras Coloradas, 20: El Sótano, 21: El Fuerte.
en varios puntos del norte de la Provincia de Buenos Aires: San Clemente del Tuyú, Santa Teresita, La Lucila del Mar, Mar de Ajó, Nueva Atlantis, Pinamar, Villa Gesell y Mar Azul, desde noviembre de 1994 a septiembre de 2000 y desde enero de 2008 a septiembre 2011. en varios puntos del sur de la Provincia de Buenos Aires: Los Pocitos, Ría del Jabalí y Bahía San Blas desde mayo de 2008 a septiembre de 2011.
en varios puntos de la zona norte del Golfo San Matías (Provincia de Río Negro): Punta Orengo, Las Garzas, Banco Reparo, San Antonio Oeste, Los Álamos, Las Grutas, El Sótano y El Fuerte desde abril de 1998 a mayo de 2000 y desde marzo de 2006 a abril de 2007.
Los ambientes costeros poco profundos en que se realizó el muestreo están caracterizados por fuerte re-suspensión de materiales del fondo causada por las corrientes de marea, lo cual explica la frecuente ocurrencia de diatomeas bénticas en el fitoplancton.
Las muestras cualitativas fueron tomadas de la parte superficial de la columna de agua (entre 0 y 5 m) con red de 30 mm de apertura de malla y fijadas con formaldehido al 4%. En el laboratorio las muestras preservadas fueron lavadas con agua destilada para remover sales y el fijador y luego tratadas para oxidar la materia orgánica por los métodos de Hasle & Fryxell (1970) y Prygiel & Coste (2000). El material fue montado para su análisis con microscopio óptico (MO) y microscoscopio electrónico de barrido (MEB) según los métodos detallados en Ferrario et al. (1995). Los montajes permanentes fueron realizados con Hyrax o Naphrax.
El material fue depositado en el Herbario de la División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata bajo los números LPC 4250 a 4495, LPC 4550 a 4645, LPC 11001 a 11305, LPC 11601 a 11816 y LPC 13648 a 13685. Las observaciones fueron realizadas con microscopio Nikon Microphot-FX con contraste de fase y las microfotografías fueron obtenidas con Leica DM 2500 con contraste diferencial de interferencia (DIC) y con microscopio electrónico de barrido Jeol JSM 6360 LV previa metalización de la muestra con oro. La terminología general utilizada es la recomendada por Ross et al. (1979) y Round et al. (1990). La terminología particular empleada para el género Pleurosigma y los detalles sobre medición de ángulos de las estrías y del rafe siguen la propuesta de Sterrenburg (1991a, b) y Reid (2002, 2012).
Resultados
Pleurosigma decorum W. Smith (Figs. 2 A-G, 3 A-H, 4 A-G)
W. Smith 1853, p. 63, lám. 21, fig. 196; Hendey 1964, p. 242.
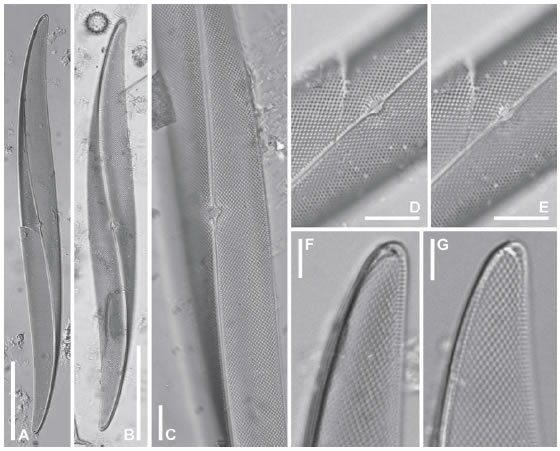
Fig. 2. Pleurosigma decorum. MO. A-B: Aspecto general de la valva, LPC 13665 y 13683 respectivamente. C: Valva abovedada mostrando el área central con dilatación bilateral, LPC 13683. D-E: Detalles de la valva en la zona del área central. Note terminaciones centrales del rafe, LPC 13665. F-G: Detalles de los polos, LPC 13668. F: Note área terminal. Escalas: A-B 50 μm, C-E 10 μm, F-G 5 μm. Fig. 3. Pleurosigma decorum. MEB, vistas externas. A-B: Aspecto general de la valva, LPC 11755. Note abovedamiento a nivel del rafe. C: Detalle de la valva en la zona central mostrando el patrón de estriación, LPC 11755. D-F: Detalles de la valva mostrando las fisuras centrales del rafe, D-E LPC 11755, F LPC 13671. G-H: Detalle del ápice mostrando la fisura terminal del rafe, LPC 11755. Escalas: A-B 50 μm; C, G-H 5 μm; D-F 2 μm.

Fig. 3. Pleurosigma decorum. MEB, vistas externas. A-B: Aspecto general de la valva, LPC 11755. Note abovedamiento a nivel del rafe. C: Detalle de la valva en la zona central mostrando el patrón de estriación, LPC 11755. D-F: Detalles de la valva mostrando las fisuras centrales del rafe, D-E LPC 11755, F LPC 13671. G-H: Detalle del ápice mostrando la fisura terminal del rafe, LPC 11755. Escalas: A-B 50 μm; C, G-H 5 μm; D-F 2 μm.
.
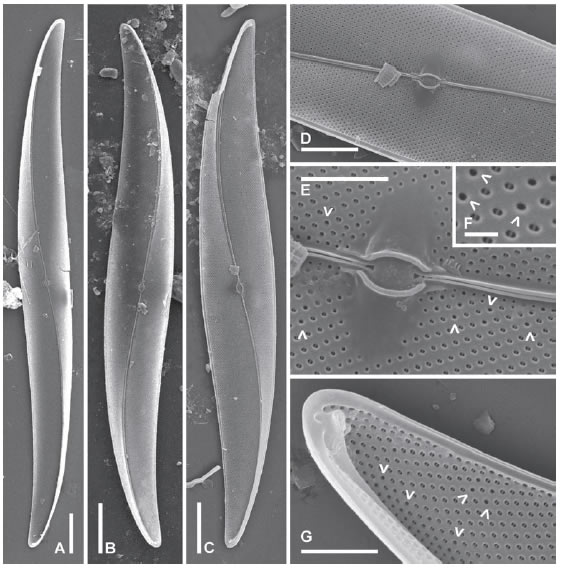
Fig. 4. Pleurosigma decorum. MEB, vistas internas. A-C: Aspecto general de la valva. Note abovedamiento a nivel del rafe-sternum, LPC 13665, 11755 y 13664 respectivamente. D-E: Detalle mostrando el área central en forma de silla de montar y el nódulo central, LPC 13664. F: Detalle de areolas con barras silíceas y pares de poros sin barras silíceas (flechas). G: Polo mostrando helictoglossa, LPC 13664. Las flechas señalan pares de poros sin barras silíceas. Escalas: A-C 20 μm, D 10 μm, E, G 5 μm, F 1 μm.
Microscopía óptica
Valvas lanceoladas, sigmoideas (Fig. 2 A-B), abovedadas (Fig. 2 C), lados entre el centro de la valva y cada polo uno convexo y el opuesto ligeramente cóncavo, con ápices sub-agudos, 185-272 μm de largo y 22-30 μm de ancho. Rafesternum sigmoideo, excéntrico, diagonal en la zona central y desviado gradualmente hacia el lado convexo de la valva hacia los polos (Fig. 2 A-C). Ángulo del rafe +13º a +16º. Área central con dilatación bilateral alrededor del nódulo central (Fig. 2 C-E), área terminal unilateralmente dilatada, infundibuliforme, en posición apical (Fig. 2 F). Estrías oblicuas 15-17 en 10 μm (Fig. 2 D-G), con un ángulo de intersección de 70-80º (Fig. 2 C-G), estrías transversas, 18-19 en 10 μm (Fig. 2 D-G).
Microscopía electrónica
Valvas abovedadas hacia los polos, elevadas en el área del rafe (Fig. 3 A-B, Fig. 4 A-C). Ramas del rafe externamente rodeadas por un área estrecha lisa (Fig. 3 C-F). Fisuras centrales del rafe onduladas en el mismo sentido, superpuestas en paralelo por alguna distancia, ligeramente divergentes y expandidas en los extremos (Fig. 3 D-F). Fisuras terminales del rafe en forma de gancho, desviadas en sentido opuesto hacia el lado cóncavo de la valva, aproximadamente paralelas al margen apical de la valva y terminadas cerca del margen valvar (Fig. 3 G-H). Rafe-sternum estrecho, internamente engrosado (Fig. 4 E, G). Área central en forma de silla de montar (Fig. 4 E), lisa, con un engrosamiento variable (Fig. 4 A-C). Nódulo central elíptico limitado por dos barras aproximadamente simétricas en forma de paréntesis (Fig. 4 D-E). Terminaciones centrales del rafe coaxiales, expandidas en forma de poro (Fig. 4 E), terminaciones polares finalizadas en helictoglossae pequeñas, torcidas hacia el margen cóncavo de la valva, bordeadas por un refuerzo silíceo (Fig. 4 G). Areolas loculadas dispuestas en hileras decusadas, con forámenes externos en forma de ranura (Fig. 3 C-H) y poros internos elípticos, ocluidos por un himen más o menos hundido (no visible en las figuras) atravesado por una sutil barra (Fig. 4 E-G). Areolas ubicadas alrededor del área central con poros ocluidos circulares, sin barra (Fig. 4 E). Esparcidos sobre la superficie valvar hay algunos poros, dispuestos de a pares, que son algo mayores en tamaño que los otros y que no presentan barra silícea (Fig. 4 E-G, flechas).
Comentarios: los especímenes analizados en el presente estudio (n = 28) fueron comparados con los procedentes del material tipo de la especie analizado en Sterrenburg et al. (enviado).
Material fotografiado: LPC 11755 Ría del Jabalí 12/10/10, LPC 13664 Las Grutas 26/05/06, LPC 13665 Piedras Coloradas 26/05/06, LPC 13668 Las Grutas 31/07/06, LPC 13671 Piedras Coloradas 05/08/06, LPC 13683 Piedras Coloradas 23/02/07.
Distribución: la especie fue erigida a partir de material procedente de Ipswich, Inglaterra. Cleve (1894) enlistó a Pleurosigma decorum como sinónimo de P. formosum W. Smith, y su criterio fue seguido por varios autores, de allí que es dificultoso determinar su distribución fehacientemente. Hendey (1964), quien analizó el material de W. Smith y lectotipificó el taxon, señaló que se trata de una especie común y ampliamente distribuida en aguas costeras del Mar del Norte. En el presente estudio P. decorum fue hallada en forma frecuente a ocasional en el plancton de Los Pocitos, Ría del Jabalí y Bahía San Blas en la Provincia de Buenos Aires y en el de Punta Orengo, San Antonio Oeste, Las Grutas, Piedras Coloradas y El Sótano en la Provincia de Río Negro.
Pleurosigma distinguendum Hustedt (Figs. 5 A-G, 6 A-G)
Hustedt 1955, p. 36, lám. 11, figs. 3-5; Sterrenburg et al. 2003, p. 297, figs. 14-20.

Fig. 5. Pleurosigma distinguendum. A: MO, B-G: MEB, vistas externas, LPC 11171. A-B: Aspecto general del frústulo en vista valvar. C: Detalle de la zona central de la valva. D: Fisuras centrales del rafe. E-F: Detalle de los polos. G: Detalle de la fisura terminal del rafe. Escalas: A-B 10 μm, C, E-F 5 μm, D, G 2 μm..
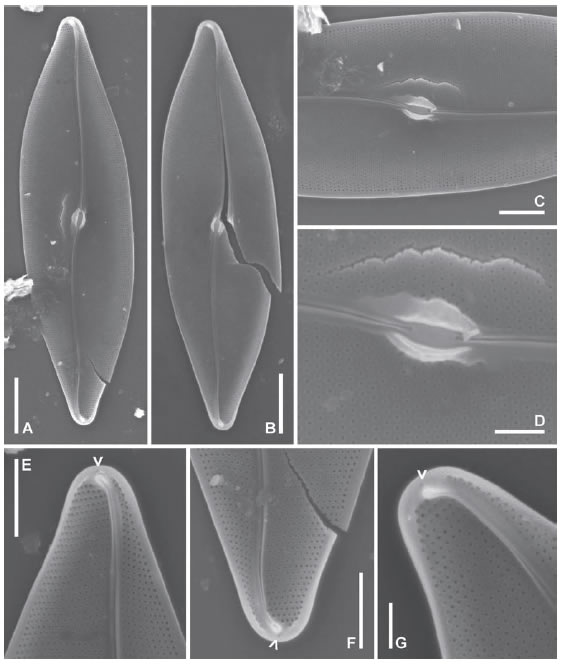
Fig. 6. Pleurosigma distinguendum. MEB, vistas internas, LPC 11171. A-B: Aspecto general de la valva. C: Detalle de la zona central de la valva. D: Detalle mostrando el área central alrededor de las barras que rodean al nódulo. E-G: Polo mostrando la helictoglossa. Note areola marginal aislada (flecha). Escala: A-B 10 μm, C, E-F 5 μm, D, G 2 μm
Microscopía electrónica
Valvas anchas, lanceoladas, no sigmoideas o muy ligeramente sigmoideas en los polos, aplanadas, con ápices sub-rostrados (Figs. 5 A-B, 6 A-B), 69-78 μm de largo, 19-20 μm de ancho. Rafe ligeramente ondulado, oblicuo en la parte central de la valva y abruptamente curvado hacia lados opuestos en la parte apical (Figs. 5 A-B, 6 A-B), externamente rodeado por un área estrecha lisa (Fig. 5 C, E-F). Ángulo del rafe +4º. Fisuras centrales del rafe desviadas en sentido opuesto, sin superposición (Fig. 5 D). Fisuras terminales desviadas hacia el lado cóncavo del rafe formando un ángulo casi recto con el último tramo de éste (Fig. 5 G). Rafe-sternum internamente asimétrico, más engrosado de un lado de la valva que del otro (Fig. 6 A-C, F). Nódulo central elíptico, oblicuamente ubicado, limitado por dos barras anchas y gruesas, aproximadamente simétricas (Fig. 6 C-D). Área central en forma de pequeña plataforma alrededor de las barras que rodean el nódulo (Fig. 6 C-D). Terminaciones centrales del rafe coaxiales, ligeramente expandidas en forma de poro (Fig. 6 D), terminaciones polares finalizadas en helictoglossae pequeñas, torcidas hacia el lado cóncavo del rafe, rodeadas apicalmente por un área lisa generalmente interrumpida por una areola marginal aislada (Fig. 6 E-G). Estrías oblicuas 24-25 en 10 μm y estrías transversas paralelas en el centro y convergentes en los polos, 24-28 en 10 μm tanto en el centro como en los polos. Ángulo de intersección de las estrías oblicuas 55-61º en el centro (Fig. 5 C). Areolas loculadas dispuestas en hileras decusadas, con foramen externo en forma de ojal (Fig. 5 C-G) y poros internos ocluidos, circulares y rebordeados (Fig. 6 C-G). Areolas de los polos con un patrón desordenado en vista externa (Fig. 5 E-G). Algunos forámenes de la zona central de la valva y de los ápices, tienen su eje mayor dispuesto en diferente posición a los del resto de la estría (Fig. 5 D, G).
Material fotografiado: LPC 11171 San Clemente del Tuyú 04/12/09.
Comentarios: los especímenes analizados en el presente estudio (n = 4) fueron comparados con los ilustrados en el protologo (Hustedt 1955) y con el material tipo designado por Simonsen (1987, pl. 624, figs. 3-6).
Distribución: Los datos de distribución de Pleurosigma distinguendum son escasos en la bibliografía, ella fue descripta de material procedente de sedimentos de la playa de Beaufort, Carolina del Norte, Estados Unidos de Norte América (Hustedt 1955) y posteriormente hallada por Sterrenburg et al. (2003) en muestras planctónicas procedentes de aguas costeras de Zihuatanejo, Estado de Guerrero y de Caleta de Campos, Estado de Michoacán, México y en material béntico de Mission Bay, California, Estados Unidos de Norte América. Durante el presente estudio la especie fue ocasionalmente detectada en muestras de plancton de San Clemente del Tuyú (Provincia de Buenos Aires), y es citada por primera vez para Argentina y para el Atlántico Sudoccidental.
Pleurosigma inflatum Shadbolt (Figs. 7 A-E, 8 A-G, 9 A-F)
Shadbolt 1853, 16, fig. 9; Sterrenburg 2001, p. 121, figs. 1-5, 11-14.

Fig. 7. Pleurosigma inflatum. MO, LPC 4608. A-B: Aspecto general de la misma valva en diferentes focos. C-D: Detalle de la parte central de la valva en diferentes focos. C: Note fisuras centrales del rafe y las estría oblicuas ligeramente curvas. D: Note terminaciones centrales del rafe. E: Polo mostrando área terminal pequeña. Escalas: A-B 10 mm, C-E 5 mm.

Fig. 8. Pleurosigma inflatum. MEB, vistas externas. A: Aspecto general del frústulo en vista valvar inclinada mostrando la cintura, LPC 13665. Note depresión en forma de H sobre la superficie valvar en la parte media. B: Aspecto general de la valva, LPC 11002. C-D: Detalle de la parte central de la valva, LPC 4285 y 11002 respectivamente. E-F: Detalle de las fisuras centrales del rafe, LPC 11002 y 4644 respectivamente. G: Detalle de un polo mostrando la fisura terminal del rafe, LPC 4285. Note el poro apical aislado (flecha). Escalas: A-B 10 μm; C-D 5 μm, E-G 2 μm.

Fig. 9. Pleurosigma inflatum. MEB, vistas internas. A-B: Aspecto general de la valva, LPC 13672 y 4258 respectivamente. C: Detalle de la parte central de la valva, LPC 4258. D-E: Área central en forma de silla de montar alrededor de las barras que delimitan el nódulo central, LPC 4258. E: Note que las barras están interrumpidas en la parte media. F: Polo mostrando helictoglossa, LPC 4258. Note el poro apical aislado. Escalas: A-B 10 μm, C 5 μm, D-F 2 μm.
Microscopía óptica
Valvas ampliamente lanceoladas, con los polos apenas desviados en sentido opuesto, ápices redondeados, sub-rostrados (Fig. 7 A-B), 80-107 mm de largo, 15-22 mm de ancho. Rafe-sternum sigmoideo, excéntrico, diagonal en la zona central y desviado gradualmente hacia el lado convexo de la valva en el área de los polos (Fig. 7 A-B). Ángulo del rafe de alrededor de +10º a +13º. Área central grande, circular, con dilatación bilateral alrededor del nódulo central (Fig. 7 C-D), área terminal pequeña, apenas visible, en posición apical (Fig. 7 E). Estrías transversas paralelas (Fig. 7 D-E), 22-24 en 10 mm. Estrías oblicuas, ligeramente curvas en el centro (Fig. 7 C), 18-20 en 10 mm, con un ángulo de intersección de 60-64º semejante en el centro y en los polos (Fig. 7 C-E).
Microscopía electrónica
Valvas abovedadas hacia los ápices, más aplanadas en la zona central (Fig. 8 A-B), en la que se marca una depresión en forma de H (Fig. 8 A, D). Rafe externamente rodeado por un área lisa (Fig. 8 C-D, G). Fisuras centrales del rafe apenas onduladas, ligeramente desviadas en sentido opuesto o excepcionalmente en el mismo sentido, con terminaciones levemente expandidas (Fig. 8 C-F). Fisuras terminales del rafe oblicuamente desviadas en sentido opuesto hacia el lado cóncavo de la valva con el último tramo en forma de gancho, delimitando un área lisa hacia el lado convexo del rafe (Fig. 8 G). Rafe-sternum internamente engrosado hacia los polos (Fig. 9 F), rodeado por un área lisa de ancho regular (Fig. 9 A-F). Área central en forma de silla de montar, lisa, semejante a un stauros (Fig. 9 C-E). Nódulo central elíptico delimitado por dos delgadas barras simétricas, sub-paralelas entre sí (Fig. 9 C-D), raramente interrumpidas en la parte media (Fig. 9 E). Terminaciones centrales del rafe coaxiales, ligeramente expandidas en forma de poro (Fig. 9 C-E), terminaciones polares finalizadas en helictoglossae pequeñas, oblicuamente dispuestas (Fig. 9 F). Areolas loculadas en hileras decusadas, con forámenes externos sub-circulares en la zona central de la valva y en forma de ojal, alargados en sentido apical hacia los polos (Fig. 8 C, G), poros internos ocluidos, circulares y rebordeados (Fig. 9 C-F).
Material fotografiado: LPC 4258 Santa Teresita 27/03/95, LPC 4285 Villa Gesell 26/04/96, LPC 4608 El Sótano 30/04/99, LPC 11002 Santa Teresita 08/01/08, LPC 13672 Las Grutas 05/08/06. Comentarios: los especímenes analizados en el presente estudio (n = 19) fueron comparados con especímenes procedentes del material tipo y de otras áreas geográficas previamente ilustrados por Sterrenburg (2001, figs. 1-5, 11-16).
Distribución: Los datos de distribución de Pleurosigma inflatum son escasísimos en la bibliografía, ella fue descripta de material procedente de Port Natal, Durban, Sudáfrica (Shadbolt 1853) y posteriormente fue hallada por Sterrenburg (2001) en aguas marinas litorales del oeste de Europa, del Mar Caribe, del Mar Mediterráneo y del Mar del Norte. Durante el presente estudio la especie fue ocasionalmente detectada en muestras de plancton procedentes de todos los puntos de muestreo de las tres áreas y es citada por primera vez para Argentina.
Pleurosigma inscriptura M. A. Harper (Figs. 10AD, 11A-F)
Harper et al. 2009, p. 343, figs. 18-22.
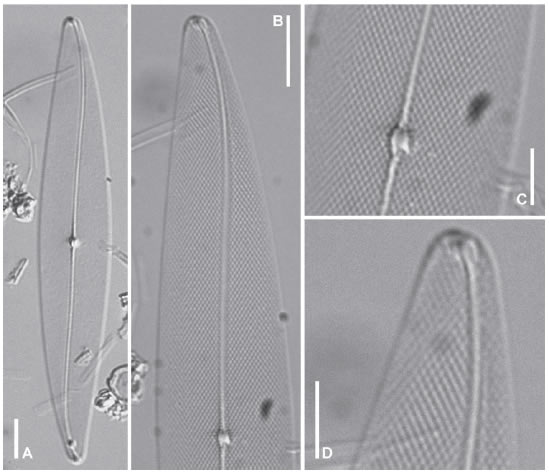
Fig. 10. Pleurosigma inscriptura. MO, LPC 4606. A: Aspecto general de la valva. B: Detalle de una valva mostrando el patrón de estriación. C: Detalle de la parte central de la valva mostrando las terminaciones del rafe. D: Polo mostrando el área terminal en forma de triángulo, de posición apical. Escalas: A-B 10 μm, C-D 5 μm.

Fig. 11. Pleurosigma inscriptura. MEB, A, C-D vistas externas, LPC 13668, B, E-F vistas internas, LPC 13668. A: Aspecto general de un frústulo fracturado y erosionado. B: Aspecto general de la valva. C: Detalle de la parte media de la valva mostrando las fisuras centrales del rafe. D: Detalle de un polo mostrando la fisura terminal del rafe. E: Nódulo central del rafe rodeado por dos barras simétricas. F: Detalle de un polo mostrando el final del rafe terminado en helictoglossa. Note el área lisa que rodea a la helictoglossa y la hilera de areolas en forma de ojal alrededor del margen apical. Escalas: A-B 20 μm; C-F 2 μm.
Microscopía óptica
Valvas lanceoladas con ápices ampliamente redondeados, contorno casi simétrico respecto del eje apical (Fig. 10 A), 115-146 μm de largo, 19-22 μm de ancho. Rafe-sternum sigmoideo, casi recto en la zona central de la valva y desviado gradualmente en sentido opuesto hacia los polos (Fig. 10 A). Ángulo del rafe de +3º a +4º. Área central pequeña, elíptica (Fig. 10 B-C), área terminal pequeña, apenas visible, en forma de triángulo, de posición apical (Fig. 10 D). Estrías transversas paralelas, 20-23 en 10 mm. Estrías oblicuas, 17-20 en 10 mm, con un ángulo de intersección de 62-70º (Fig. 10 B-D).
Microscopía electrónica
Valvas planas (Fig. 11 A-B). Rafe externamente rodeado por un área estrecha lisa (Fig. 11 C-D). Fisuras centrales del rafe onduladas en la misma dirección, con sus extremos apenas superpuestos (Fig. 11 C). Fisuras terminales del rafe en forma de gancho abierto, desviadas en sentido opuesto, delimitando un área lisa hacia el lado convexo del rafe-sternum (Fig. 11 D). Raphe-sternum internamente engrosado (Fig. 11 E-F), nódulo central pequeño, rodeado por dos barras simétricas en forma de paréntesis (Fig. 11E). Terminaciones centrales del rafe coaxiales, apenas dilatadas (Fig. 11 E). Terminaciones polares del rafe finalizadas en helictoglossae desviadas en sentido opuesto, lateralmente rodeadas por un área lisa delimitada hacia el margen valvar apical por una hilera de areolas en forma de ojal pervalvarmente alargado (Fig. 11 F). Areolas loculadas, ordenadas en estrías decusadas, con forámenes externos en forma de ranura ubicados en sentido apical (Fig. 11 C-D) y poros internos elípticos, ocluidos por hímenes con una gruesa barra central (Fig. 11 E-F). Poros internos ubicados alrededor de las barras que limitan el nódulo central, circulares, ocluidos por un himen sin barra (Fig. 11 E).
Material fotografiado: LPC 4606 Punta Orengo 13/04/99, LPC 13668 Las Grutas 31/07/06, LPC 13672 Las Grutas 05/08/06.
Comentarios: los especímenes analizados en el presente estudio (n = 10) fueron comparados con los ilustrados en el protologo (Harper et al. 2009, figs. 18-22).
Distribución: Hasta el presente Pleurosigma inscriptura sólo ha sido citada por Harper et al. (2009) para Nueva Zelanda procedente de la costa de Wellington, colectada junto con algas rojas, de Island Bay, hallada sobre macroalgas sublitorales de los géneros Herposiphonia, Halopteris y Gigartina y de Owhiro Bay, hallada sobre coralináceas. Durante el presente estudio P. inscriptura ha sido ocasionalmente encontrada en muestras planctónicas procedentes de Las Grutas y Piedras Coloradas, Provincia de Río Negro. Esta es la primera cita del taxon para Argentina y para el Atlántico Sud-Americano.
Pleurosigma normanii Ralfs in Pritchard (Figs. 12 A-G, 13 A-I, 14 A-I)
Pritchard 1861, p 919; Peragallo 1891, lám. IV, p, 10, figs. 6-7 [como P. affine Grunow var. normanii (Ralfs) H. Peragallo]; Sterrenburg 1990, p. 236, figs. 2-6, 12-14.

Fig. 12. Pleurosigma normanii. MO. A-B: Células mostrando plástidos, LPC 11745 y 11276 respectivamente. C: Aspecto general de la valva, LPC 4262. D: Detalle de la valva mostrando el patrón de estriación, LPC 4262. E: Detalle de la parte media de la valva mostrando las fisuras centrales del rafe, LPC 13675. F-G: Polo mostrando el área terminal infundibuliforme, LPC 13675 y 4262 respectivamente. Escalas: A-C 20 μm, D 10 μm, E-G 5 μm.

Fig. 13. Pleurosigma normanii. MEB, vistas externas. A: Frústulo en vista valvar ligeramente inclinada mostrando la cintura, LPC 13669. B-C: Aspecto general de la valva, LPC 4262. D: Parte media de la valva mostrando las fisuras centrales del rafe, LPC 13668. E: Detalle de la parte media de un frústulo en vista conectival inclinada para mostrar la valva, LPC 11078. Note las depresiones centrales paralelas al rafe. F-G: Detalle de las fisuras centrales del rafe, LPC 13667 y 4262 respectivamente. H-I: Polos mostrando la fisura terminal del rafe, LPC 4262. Note el poro aislado en el margen apical (flecha). Escalas: A-C 20 μm, D-E 5 μm, F-I 2 μm.
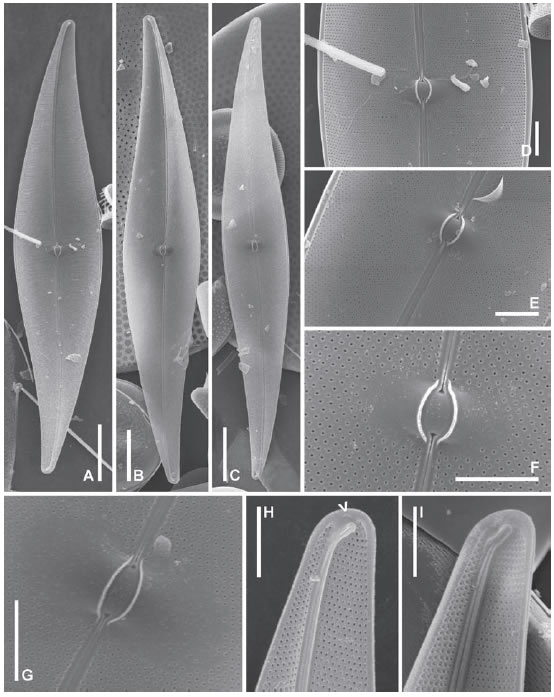
Fig. 14. Pleurosigma normanii. MEB, vistas internas. A-C: Aspecto general de la valva, LPC 4262. D-E: Detalle de la parte central de la valva. Note que la valva está inclinada, LPC 4262. F-G: Área central en forma de silla de montar alrededor del nódulo central delimitado por dos barras delgadas simétricas, LPC 4262. H-I: Polos mostrando las terminaciones del rafe-sternum finalizadas en helictoglossa y en H poro apical aislado (flecha), LPC 4262 y 11220 respectivamente. Escalas: A-C 20 μm, D-I 5 μm.
Microscopía óptica
Valvas lanceoladas a ampliamente lanceoladas, sigmoideas (Fig. 12 A-C), más o menos abovedadas a nivel del rafe-sternum (Fig. 12 D), con ápices sub-agudos a redondeados, 115-211 μm de largo, 21-35 μm de ancho. Rafe-sternum sigmoideo, aproximadamente central y recto en la porción media, desviado gradualmente en sentido opuesto hacia el lado convexo de la valva en la región polar (Fig. 12 C). Ángulo del rafe de alrededor de +4º a +7,8º. Área central circular, con dilatación bilateral alrededor del nódulo central (Fig. 12 C-E). Fisuras centrales del rafe muy evidentes, onduladas en el mismo sentido (Fig. 12 E). Área terminal pequeña, infundibuliforme, dilatada en posición apical (Fig. 12 F-G). Estrías transversas 20-24 en 10 μm, ligeramente más densas hacia los polos, 22-25 en 10 μm. Estrías oblicuas 18-22 en 10 μm en el centro de la valva y 20-23 en 10 μm en los polos. Ángulo de intersección de las estrías oblicuas 63-70º en el centro (Fig. E) y 54-60º en los polos (Fig. F-G).
Microscopía electrónica
Valvas aplanadas en la zona central (Fig. 13 A-B) o con dos cortas depresiones subcentrales paralelas al margen valvar en algunos especímenes (Fig. 13 C), abovedadas a nivel del rafe hacia los polos (Fig. 13 A-C). Rafe externamente rodeado por un área estrecha lisa limitada por una hilera de areolas densamente dispuestas, 20-25 en 10 μm (Fig. 13 D, F, H-I). Fisuras centrales del rafe simétricamente onduladas, curvadas en el mismo sentido (Fig. 13 D-G), con terminaciones levemente expandidas sólo en algunos especímenes (Fig. 13 D, F). Fisuras terminales del rafe oblicuamente desviadas en sentido opuesto hacia el lado cóncavo de la valva con el último tramo en forma de gancho, expandidas, delimitando un área lisa hacia el lado convexo del rafe con un poro aislado en su extremo (Fig. 13 H-I). Rafe-sternum estrecho, apenas engrosado en el centro (Fig. 14 F-G) y más engrosado hacia los extremos internamente (Fig. 14 H-I). Área central en forma de silla de montar, lisa, semejante a un stauros (Fig. 14 D-G). Nódulo central elíptico delimitado por dos barras delgadas simétricas en forma de paréntesis (Fig. 14 D-G). Terminaciones centrales del rafe coaxiales, ligeramente expandidas en forma de T (Fig. 14 F) o de poros (Fig. 14 G), terminaciones polares finalizadas en helictoglossae pequeñas, muy curvadas en sentido opuesto (Fig. 14 H-I). Areolas loculadas dispuestas en hileras decusadas, con forámenes externos en forma de ojal alargado en sentido apical (Fig. 13 D, F, H-I), a veces sub-circulares en el centro de la valva (Fig. 13 G), y poros internos circulares y rebordeados, ocluidos por un himen (Fig. 14 F, H-I).
Material fotografiado: LPC 4262 Nueva Atlantis 28/03/95, LPC 11078 Villa Gesell 01/09/08, LPC 11220 Villa Gesell 02/07/10, LPC 11276 Villa Gesell 21/03/11, LPC 11745 Los Pocitos 31/08/10, LPC 13667 Piedras Coloradas 14/06/06, LPC 13668 Las Grutas 31/07/06, LPC 13669 Piedras Coloradas 31/07/06, LPC 13675 Piedras Coloradas 01/09/06.
Comentarios: los especímenes analizados en el presente estudio (n = 42) fueron comparados con los ilustrados por Sterrenburg (1990) que de acuerdo con Sterrenburg (2003) coinciden morfológicamente con los hallados en el material tipo.
Distribución: Los datos de distribución de Pleurosigma normanii son abundantes en la bibliografía, la especie fue descripta a partir de material procedente de ambientes marinos de Europa, y hallada en prácticamente todas las muestras de aguas profundas según Ralfs in Pritchard (1861). Según Hasle & Syvertsen (1996) ésta es la especie del género más común y más ampliamente distribuida (desde el trópico a mares polares), frecuente en muestras de plancton. Sterrenburg (1990) encontró a P. normanii en materiales holocenos y actuales de Holanda, y en ambientes marinos de España, Nueva Zelanda y Puerto Rico. Durante el presente estudio la especie fue hallada frecuentemente en muestras de plancton de todas las estaciones en las tres áreas de muestreo, durante todo el año.
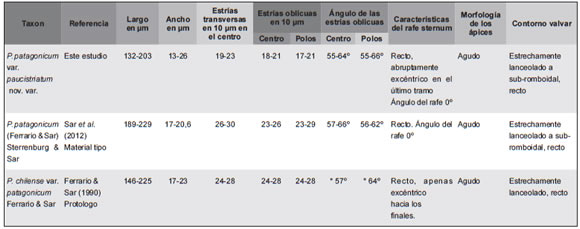
Pleurosigma patagonicum (Ferrario & Sar) Sterrenburg & Sar var. paucistriatum Sar, Sterrenburg & Sunesen nov. var. (Figs. 15 A-F, 16 A-F, 17 A-F, Tabla 1).

Fig. 15. Pleurosigma patagonicum var. paucistriatum nov. var. MO. A: Aspecto general de la valva, LPC 13671. B: Detalle de la valva mostrando el patrón de estriación y el rafe-sternum desviado abruptamente hacia el margen valvar en el polo, LPC 13665. C: Detalle de la parte central de la valva, LPC 13665. D: Detalle de las terminaciones centrales del rafe, LPC 13667. E: Polo mostrando el área terminal, LPC 13671. F: Polo mostrando la desviación del rafe-sternum, LPC 13670. Escalas: A 20 μm, B 10 μm, C-F 5 μm.

Fig. 16. Pleurosigma patagonicum var. paucistriatum nov. var. MEB, vistas externas, LPC 13668. A: Aspecto general de la valva. B-D: Detalle de las fisuras centrales del rafe. E-F: Polos mostrando desviación abrupta del rafe, fisura terminal y dos calcars (flechas). Escalas: A 20 μm, B-F 2 μm.
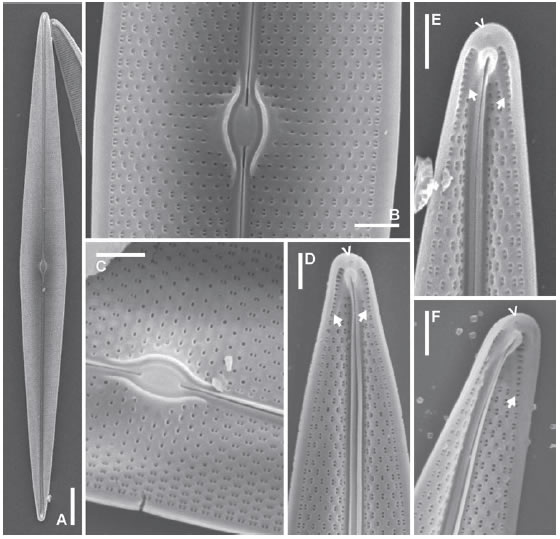
Fig. 17. Pleurosigma patagonicum var. paucistriatum nov. var. MEB, vistas internas, LPC 13668. A: Aspecto general de la valva. B-C: Detalle de la zona central de la valva. B: Note el área central en forma de pequeña plataforma alrededor de las barras que rodean el nódulo. D-F: Detalle de los polos con distinta inclinación. Note la helictoglossa y las dos áreas deprimidas con una hilera de areolas en forma de ojal (flechas rellenas) y un poro terminal (flecha). Escalas: A 10 μm, B-F 2 μm.
Tabla 1. Comparación de los parámetros morfométricos de Pleurosigma patagonicum var. paucistriatum nov. var. y P. patagonicum var. patagonicum. Abreviatura: *, dato obtenido por medición de las ilustraciones presentadas en el protologo.
Microscopía óptica
Valvas estrechamente lanceoladas a subromboidales, rectas, con ápices agudos (Fig. 15 A), 132-203 mm de largo, 13-26 mm de ancho. Rafesternum recto en prácticamente todo su recorrido, con una abrupta desviación hacia el margen valvar, de sentido contrario en ambos polos (Fig. 15 B, F). Áreas terminales bilateralmente dilatadas, simétricas, infundibuliformes, en posición apical, apenas perceptibles (Fig. 15 E). Ángulo del rafe 0º. Área central elíptica, simétrica (Fig. 15 C-D). Estrías con aproximadamente la misma densidad en el centro que en los polos, transversas 19-23 en 10 μm, oblicuas 17-21 en 10 μm, ángulo de intersección de las estrías oblicuas 55-66º (Fig. 15 C-F).
Microscopía electrónica
Valvas ligeramente abovedadas a nivel del rafe en los polos (Fig. 16 A, F). Rafe rodeado por un área lisa, más estrecha en el lado convexo y más ancha en el lado cóncavo de las ramas en los polos (Fig. 16 E-F). Fisuras centrales del rafe onduladas en el mismo sentido, terminadas a muy corta distancia o apenas superpuestas (Fig. 16 B-D). Fisuras terminales del rafe oblicuamente dispuestas en los ápices, terminadas en forma de gancho en la proximidad del margen valvar (Fig. 16 E-F). Rafesternum internamente engrosado (Fig. 17 B-F). Nódulo central elíptico, grande, limitado por dos barras simétricas en forma de paréntesis (Fig. 17 A-C). Área central en forma de pequeña plataforma alrededor de las barras que rodean el nódulo (Fig. 17 B). Terminaciones centrales del rafe coaxiales (Fig. 17 B-C). Helictoglossae ligeramente desviadas en sentido opuesto, rodeadas por un área proximal lisa, delimitada hacia el margen valvar por una hilera de areolas en forma de ojal, transapicalmente alargadas (Fig. 17 D-F flechas rellenas) localizadas en una ligera depresión, visibles externamente como dos pequeñas fisuras apicales, "calcars" (Fig. 16 E-F, flechas). Estructura apical con un poro aislado adyacente a las helictoglossae (Fig. 17 D-F, flecha). Areolas loculadas dispuestas en hileras decusadas, con forámenes externos en forma de ranura (Fig. 16 B, E-F) y poros internos elípticos ocluidos por un himen atravesado por una barra (Fig. 17 B-F). Areolas ubicadas en un rectángulo transapical alrededor del nódulo central y de un tramo corto del rafe-sternum, con poros ocluidos circulares sin barra (Fig. 17 B-C).
Holotipo: preparado LPC 13671 (3) aquí designado, rotulado "holotipo de Pleurosigma patagonicum var. paucistriatum, Piedras Coloradas, 05/08/06", depositado en el Herbario de la División Ficología, Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Isotipo: preparado LPC 13671 (4) aquí designado, rotulado "isotipo de Pleurosigma patagonicum var. paucistriatum, Piedras Coloradas, 05/08/06", depositado en el Herbario de la División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, La Plata, Argentina. Localidad tipo: Piedras Coloradas, 40º 50' 33''S-65º 06' 54''O, Prov. de Río Negro, Argentina.
Etimología del nombre: esta variedad de P. patagonicum recibió el epíteto "paucistriatum" del Latín pauci: poco (corto en número) y strãtum: estriada, para denotar que presenta menor densidad de estrías que la variedad tipo.
Material fotografiado: LPC 13665 Piedras Coloradas 26/05/06, LPC 13667 Piedras Coloradas 14/06/06, LPC 13668 Las Grutas 31/07/06, LPC 13670 Las Grutas 25/07/06, LPC 13671 Piedras Coloradas 05/08/06.
Comentarios: los especímenes estudiados (n = 29) fueron comparados con los hallados en el material tipo de P. patagonicum (Ferrario & Sar) Sterrenburg & Sar var. patagonicum recientemente analizado en Sar et al. (2012). Distribución: Pleurosigma patagonicum var. paucistriatum fue hallada ocasionalmente en el plancton de Piedras Coloradas y Las Grutas, Provincia de Río Negro, en muestras colectadas durante los años 2006 y 2007.
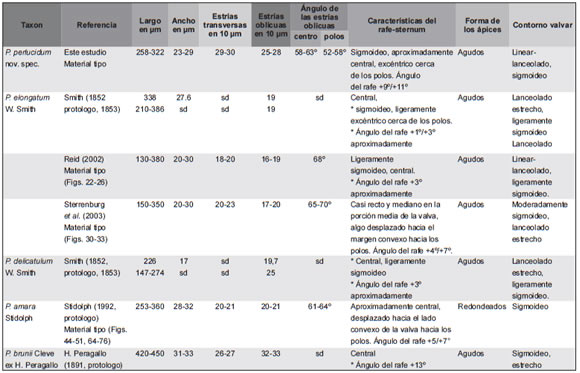
Pleurosigma perlucidum Sar, Sterrenburg & Sunesen nov. spec. (Figs. 18 A-H, 19 A-H, 20 A-F, Tabla 2).

Fig. 18. Pleurosigma perlucidum nov. spec. MO. A-C: Aspecto general de la valva, A-B LPC 4260 y C LPC 4606. D-E: Detalle de la parte central de la valva mostrando las terminaciones del rafe, LPC 4644 y 4260 respectivamente. E: Note el patrón de estriación. F-G: Detalle de un polo. F: Note el rafe-sternum y el área terminal de posición apical, LPC 4644. G: Note el patrón de estriación, LPC 4260. H: Detalle del área terminal, LPC 4260. Escalas: A-C 50 μm, D-H 5 μm.

Fig. 19. Pleurosigma perlucidum nov. spec. MEB, vistas externas, LPC 4261. A-B: Aspecto general de la valva. C: Detalle de la zona central de la valva. D-E: Detalle de las fisuras centrales del rafe. F: Área quebrada de la valva mostrando las areolas loculadas con forámenes externos y poros ocluidos atravesados por una barra. G-H: Polo mostrando fisura terminal en forma de gancho y poro apical aislado (flecha). Escalas: A-B 50 μm, C, G 5 μm, D-F, H 2 μm.

Fig. 20. Pleurosigma perlucidum nov. spec. MEB, vistas internas. A-C: Aspecto general de la valva, A-B LPC 4260 y C LPC 4262. D: Detalle del área central de la valva, LPC 4260. E: Detalle del nódulo central, LPC 4260. Note las areolas alrededor del nódulo mostrando poros ocluidos circulares sin barra. F-G: Polo mostrando la helictoglossa y poro apical aislado (flecha), LPC 4260 y LPC 4262 respectivamente. Escalas: A-B 50 μm, C-D 5 μm, E-G 2 μm.
Tabla 2. Comparación de los datos morfométricos de Pleurosigma perlucidum nov. spec. procedente del área de estudio, con datos obtenidos de los protologos e ilustraciones de los materiales tipo de P. amara, P. elongatum P. delicatulum y P. brunii. Abreviaturas sd, sin datos; *, dato obtenido por medición de las figuras en la literatura.
Microscopía óptica
Valvas muy delicadas, linear-lanceoladas, netamente sigmoideas, lados entre el centro de la valva y cada polo uno convexo y el opuesto cóncavo, ápices agudos (Fig. 18 A-C), 258-322 μm de largo y 23-29 μm de ancho. Rafe-sternum sigmoideo, aproximadamente central en la mayor parte de la valva y desviado en sentido opuesto hacia el lado convexo de la valva en el área de los polos (Fig. 18 A-C). Ángulo del rafe +9º a +11º. Área central circular, pequeña (Fig. 18 D-E), área terminal pequeña, infundibuliforme, de posición apical (Fig. 18 F-H). Estrías difícilmente visibles, aproximadamente de igual densidad en el centro y en los polos, transversas 29-30 en 10 μm, oblicuas 25-28 en 10 μm, ángulo de intersección de las estrías oblicuas a 58-63º en el centro y 52-58º en los polos. Estrías transversas más evidentes que las oblicuas.
Microscopía electrónica
Valvas abovedadas a nivel del rafe en la proximidad de los polos (Fig. 19 A-B). Rafe externamente rodeado por un área estrecha lisa en la zona central de la valva (Fig. 19 C-E) más ancha en la zona polar (Fig. 19 G-H). Fisuras centrales del rafe onduladas en sentido opuesto, más o menos superpuestas en el último tramo (Fig. 19 D-E). Fisuras terminales del rafe en forma de gancho, desviadas en sentido opuesto hacia el lado cóncavo de la valva (Fig. 19 G-H). Poros aislados ubicados uno en cada ápice, en el lado opuesto de las fisuras terminales (Fig. 19 G-H). Rafe-sternum estrecho, internamente engrosado (Fig. 20 A-G). Nódulo central elíptico, limitado por dos barras simétricas (Fig. 20 D-E). Terminaciones centrales del rafe, coaxiales, apenas expandidas en forma de poro (Fig. 20 E). Terminaciones polares finalizadas en helictoglossae, ligeramente desviadas en sentido opuesto hacia el lado cóncavo de la valva, rodeadas por un área lisa (Fig. 20 F-G). Areolas del margen valvar apical interrumpidas al final de la helictoglossa del lado cóncavo de la valva y por debajo de ésta en el lado convexo, excepto por una o dos areolas aisladas próximas a su extremo (Fig. 20 F-G). Areolas loculadas dispuestas en hileras decusadas, con forámenes externos en forma de ranura (Fig. 19 D-H) y poros internos elípticos, ocluidos por un himen más o menos hundido, atravesado por una barra (Fig. 20 E-G). Estrías oblicuas y transversas poco visibles en vista externa, con forámenes alineados en sentido apical. Areolas ubicadas alrededor del nódulo central y de un tramo corto del rafe-sternum con poros ocluidos circulares, sin barra (Fig. 20 E). Holotipo: preparado LPC 4260 (3) aquí designado, rotulado "holotipo de Pleurosigma perlucidum, Nueva Atlantis, 27/03/1995", depositado en el Herbario de la División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Isotipo: preparado LPC 4260 (4) aquí designado, rotulado "isotipo de Pleurosigma perlucidum, Nueva Atlantis, 27/03/1995", depositado en el Herbario de la División Ficología, Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Localidad tipo: Nueva Atlantis, 36º 45' 81''S-56º 40' 04''O, Prov. de Buenos Aires, Argentina. Etimología del nombre: esta especie recibió el epíteto específico "perlucidum" del Latín para denotar que las valvas se ven completamente transparentes cuando se las analiza con microscopio de luz en campo claro.
Material fotografiado: LPC 4260 Nueva Atlantis 27/03/95, LPC 4261 Nueva Atlantis 27/03/95, LPC 4262 Nueva Atlantis 27/03/95, LPC 4606 Punta Orengo 13/04/99, LPC 4644 Piedras Coloradas 26/05/00.
Comentarios: los especímenes estudiados (n = 38) fueron comparados con los ilustrados en los protologos y publicaciones más recientes de especies morfológicamente allegadas, P. elongatum W. Smith, P. delicatulum W. Smith, P. amara Stidolph y P. brunii H. Peragallo.
Distribución: durante el presente estudio Pleurosigma perlucidum fue hallada ocasionalmente en muestras planctónicas de Nueva Atlantis y Pinamar, Provincia de Buenos Aires y de Punta Orengo, Piedras Coloradas y El Sótano, Provincia de Río Negro.
Pleurosigma simonsenii Hasle (Fig. 21 A-J) Sinónimo: Pleurosigma planctonicum Simonsen.
Simonsen 1974: 46, lám. 30, figs. a-d; Hasle in Hasle & Syvertsen 1996: 339 (nombre correcto); Sar et al. 2012: 12, figs. 59-71.

Fig. 21. Pleurosigma simonsenii. A: MO, B, D-E, I: MEB, vista externa, C, F-H, J: MEB, vistas internas. A: Aspecto general de una célula con plástidos, LPC 11272. B: Frústulo en vista valvar, LPC 11100. C: Aspecto general de la valva, LPC 11100. D-E: Detalle de las fisuras centrales del rafe, LPC 11100. F: Detalle de la zona central de la valva, LPC 11100. G-H: Detalle del nódulo central, LPC 11100. I: Detalle de un polo mostrando la fisura terminal del rafe, LPC 11272. J: Detalle de un polo mostrando la helictoglossa, LPC 11100. Escalas: A-C 50 μm, F 10 μm, D-E, I-J 5 μm, G-H 2 μm.
Microscopía óptica
Valvas muy delicadas, lanceoladas, ligeramente sigmoideas en los polos, ápices agudos (Fig. 21 A), 250-598 mm de largo, 30-73 mm de ancho. Rafe-sternum recto excepto en los polos, en los que es excéntrico (Fig. 21 A). Ángulo del rafe de alrededor de +1,6º a +2,7º. Área central pequeña, redondeada, área terminal unilateralmente dilatada, infundibuliforme, asimétrica, de posición apical. Estrías oblicuas, 26-28 en 10 mm, con un ángulo de intersección de 56 a 60º. Estrías transversas, paralelas, 30-32 en 10 mm.
Microscopía electrónica
Valvas ligeramente abovedadas (Fig. 21 F). Rafe externamente rodeado por un área estrecha lisa (Fig. 21 D-E, I). Fisuras centrales del rafe onduladas en la misma dirección, con terminaciones coaxiales o ligeramente desplazadas (Fig. 21 D-E). Fisuras terminales del rafe desviadas oblicuamente en sentido opuesto, hacia el lado ligeramente cóncavo de la valva, delimitando un área terminal lisa, grande (Fig. 21 I). Rafe-sternum estrecho, engrosado internamente (Fig. 21 F-H, J). Nódulo central rodeado por dos barras simétricas, terminaciones centrales del rafe ligeramente dilatadas, coaxiales (Fig. 21 G-H). Terminaciones polares del rafe finalizadas en helictoglossae desviadas en sentido contrario (Fig. 21 J). Areolas loculadas, ordenadas en estrías decusadas, con forámenes externos en forma de ranura ubicados en sentido apical (Fig. 21 E, I) y poros internos elípticos, ocluidos por un himen atravesado por una barra central (Fig. 21 G-H, J). Poros internos de las areolas periféricas al nódulo central, circulares, ocluidos por un himen sin barra (Fig. 21 H).
Material fotografiado: LPC 11100 Mar Azul 12/01/09, LPC 11272 Villa Gesell 25/02/11.
Comentarios: los especímenes estudiados (n = 41) fueron comparados con los procedentes del material tipo previamente analizado por Sar et al. (2012).
Distribución: De acuerdo con Hasle & Syvertsen (1996) Pleurosigma simonsenii es probablemente cosmopolita. Ella ha sido citada para el Océano Indico por Simonsen (1974), para el Canal Inglés por Boalch & Harbour (1977) y para Port Hacking, New South Wales, Australia y Mar Azul, Provincia de Buenos Aires, Argentina por Sar et al. (2012). Esta especie ha sido hallada ocasionalmente en muestras de San Clemente del Tuyú, Santa Teresita, Nueva Atlantis, Pinamar, Villa Gesell y Mar Azul, en verano, otoño e invierno.
Discusión
El material de Pleurosigma decorum analizado en este estudio presenta algún grado de variación respecto del material tipo de la especie analizado en Sterrenburg et al. (enviado). En los especímenes hallados el área en forma de silla de montar varió entre muy engrosada como en el tipo (Fig. 4 C, E) a más sutilmente engrosada (Fig. 4 A-B) y el ángulo de intersección de las estrías oblicuas osciló entre 70 y 80º mientras que el del material tipo lo hizo entre 76 y 83º. Sin embargo, el material estudiado fue asignado a la especie sin duda porque comparte con el material tipo caracteres morfológicos tales como la superposición de las fisuras centrales del rafe, la forma de las fisuras terminales, la morfología apical en vista interna, los poros ocluidos de las areolas cruzados por barras hundidas y la presencia de pares de poros areolares esparcidos, más excavados que los contiguos y carentes de barras (Fig. 4 E-F, flechas).
Los especímenes de Pleurosigma distinguendum hallados en nuestro material son ligeramente más anchos que los descriptos e ilustrados en el protologo (Hustedt 1955, figs. 3-5) y que los fotografiados y designados como lectotipo y paralectotipo de la especie por Simonsen (1987), sin embargo ellos comparten con el material tipo el característico patrón de estriación irregular de los polos. El material del área de estudio guarda mayor similitud en el contorno valvar con los especímenes procedente de aguas costeras del Pacífico mexicano ilustrados por Sterrenburg et al. (2003, figs. 15, 17-18). P. distinguendum presenta similitud en el contorno valvar y patrón del rafe-sternum con P. ibericum Peragallo y con P. curiosa van den Heuvel. La comparación de las ilustraciones del lectotipo y paralectotipo de P. distinguendum con las del lectotipo de P. ibericum (designado por Sterrenburg et al. 2003, figs. 8-10), permite detectar una diferencia notable en el área central (que no está lateralmente expandida en P. distinguendum y lo está en P. ibericum) y en el ángulo del rafe-sternum (+4º en P. distinguendum y +2º en P. ibericum). Pleurosigma curiosa van den Heuvel in Sterrenburg (1991a, figs. 22-28) difiere de P. distinguendum por presentar mayor ángulo del rafe (+6-9º en la primera y +4º en la segunda), mayor ángulo de intersección de las estrías oblicuas (60-70º en la primera y 55-61º en la segunda), menor densidad de las estrías oblicuas (18-22 en 10 μm en la primera y 24-25 en 10 μm en la segunda) y menor densidad de las estrías transversas (21-24 en 10 μm en la primera y 24-28 en 10 μm en la segunda). A partir del análisis ultraestructural de P. distinguendum llevado a cabo en este estudio es posible establecer que, además de las diferencias morfométricas enunciadas, ambas especies muestran diferencias en la morfología de las barras simétricas que rodean el nódulo central (más sutiles en P. curiosa que en P. distinguendum), en la posición del nódulo (oblicuo entre las dos ramas del rafe en la segunda) y en la expansión del nódulo central en un área central (en forma de silla de montar en la primera y en forma de pequeña plataforma alrededor de las barras que limitan el nódulo en la segunda).
Sterrenburg (2001) analizó material de Pleurosigma inflatum procedente de Port Natal, Durban, Sudáfrica (a partir del cual Shadbolt 1853 erigió la especie), designó al preparado BM22088 como su lectotipo, amplió su descripción y determinó los caracteres diferenciales entre esta especie y P. diverse-striatum Meister. A partir del análisis comparativo del material de P. inflatum procedente del área de estudio y del ilustrado por Sterrenburg (2001) que incluye el material tipo y material procedente de otras áreas geográficas, fue posible detectar un carácter común a todos ellos, se trata de una depresión en forma de H en la zona central de la valva, que es constante en todos los especímenes de P. inflatum analizados. Esta depresión puede verse sutilmente con MO (Fig. 7 A), es muy evidente en vista externa con MEB (Fig. 8 B, D) y está posicionalmente relacionada con los límites del área central en forma de silla de montar. P. inflatum difiere de P. diverse-striatum por el ángulo del rafe, +10-13º en la primera y +17- 21º en la segunda, y por la presencia de un área deprimida en el centro de la valva en vista externa sólo presente en la primera.
Pleurosigma inscriptura fue comparada por Harper et al. (2009, tabla 1) con otras especies simétricas o apenas sigmoideas cuyos rafes son claramente sigmoideos. El material de P. inscriptura procedente del área de estudio puede distinguirse sencillamente de P. obtusum Mann (Stidolph 2002) y P. strigosum W. Smith (Sterrenburg 2003) porque las dos últimas tienen contorno sigmoideo y mayor ángulo del rafe y de P. rigidum W. Smith (Sterrenburg 2001) por el ángulo de intersección de las estrías oblicuas (mayor en la primera que en la última) y la forma del área central (no proyectada lateralmente a partir del nódulo en la primera y proyectada en forma de silla de montar en la última). La especie morfológicamente más parecida a Pleurosigma inscriptura es P. williamsii Reid (2002, figs. 9-17), sin embargo ambas pueden diferenciarse por el ángulo de intersección de las estrías oblicuas (alrededor de 65 a 73º en la primera y de 50º en la segunda) y por la dirección del rafe-sternum en la zona central de la valva, aproximadamente central en la primera y oblicuo en la segunda.
Pleurosigma normanii es una especie comúnmente citada en la literatura para ambientes marinos de diferentes latitudes, sin embargo la descripción original es somera y no existen estudios detallados sobre su morfología. Sterrenburg (1990) analizó el lectotipo de la especie (preparado BM 1376 según Sterrenburg et al. 2003) e ilustró un espécimen similar al tipo analizado con microscopía óptica (fig. 2) que presenta un ángulo de intersección de las estrías oblicuas en el centro de alrededor de 68º, con las fisuras del rafe simétricas, onduladas y desviadas en el mismo sentido. Hasta donde pudimos determinar el único análisis ultraestructural sobre la especie es el que figura en Sterrenburg (1990, figs. 12-14) que muestra la forma del área central en vista interna y un polo de la valva en vista externa e interna. El material de Argentina determinado como P. normanii muestra coincidencias en contorno, patrón del rafe-sternum y longitud de la valva con el descripto en el protologo, y en la forma del área central y de las fisuras centrales y terminales del rafe, en el ángulo de intersección de las estrías oblicuas y en la disposición de las areolas que limitan el área estrecha lisa alrededor del rafe, con el ilustrado por Sterrenburg (1990). Sobre la base del análisis morfológico llevado a cabo en este estudio ampliamos la descripción P. normanii y extendemos su rango de densidad de estrías.
El material tipo de Pleurosigma simonsenii ha sido críticamente examinado por Sar et al. (2012). En dicho trabajo, el análisis ultraestructural de la especie en vista interna fue realizado sobre el único espécimen hallado en el material sin procesar de la muestra correspondiente al holotipo y completado con el análisis de la vista externa de especímenes procedentes de aguas costeras del área correspondiente al presente estudio.
Pleurosigma patagonicum var. paucistriatum Sar, Sterrenburg & Sunesen nov. var. fue erigida como variedad de P. patagonicum por presentar similitudes en cuanto al contorno valvar, forma de los ápices, ángulo del rafe, ángulo de intersección de las estrías oblicuas, aspecto de los poros ocluidos de las areolas y presencia de calcars (ver Figs. 15-17 comparativamente con figs. 59-76 en Sar et al. 2012). La nueva variedad se diferencia de la variedad tipo por presentar menor densidad de estrías transversales y oblicuas (ver Tabla 1), una desviación abrupta del rafe-sternum en ambos polos, un área central en forma de pequeña plataforma alrededor de las barras que rodean el nódulo y una estructura apical con un poro aislado adyacente a las helictoglossae.
Pleurosigma perlucidum Sar, Sterrenburg & Sunesen nov. spec. presenta alguna similitud en el aspecto general de la valva con P. elongatum W. Smith (Smith 1852, Stidolph 1992, Reid 2002, Sterrenburg et al. 2003), P. delicatulum W. Smith (Smith 1852), P. amara Stidolph (Stidolph 1992) y P. brunii H. Peragallo (Peragallo 1891) (ver Tabla 2). Sin embargo, las valvas de P. perlucidum son más sigmoideas que las de P. elongatum, P. delicatulum y P. amara y menos sigmoideas que las de P. brunii. La nueva especie difiere de P. elongatum, P. delicatulum y P. amara en la densidad de las estrías transversas (29-30 vs. 19-23, 19-25 y 20-21 en 10 μm respectivamente), en la densidad de las estrías oblicuas (25-28 vs. 16-20, 19,7-25 y 20-21 en 10 μm respectivamente), y en el ángulo del rafe (+9-11º vs. +3-7º, +3º y +5-7º). Adicionalmente, el ángulo de intersección de las estrías oblicuas es menor en P. perlucidum que en P. elongatum (52-63º vs. 65-70º), mientras que no hay diferencia entre P. perlucidum y P. amara (52-63º vs. 61-64º) y no hay datos sobre este carácter en los protologos de P. delicatulum y P. brunii. Además de las diferencias morfométricas señaladas, P. perlucidum presenta diferencias apreciables en la ultraestructura apical interna con P. elongatum y P. amara, en la nueva especie las areolas que rodean las helictoglossae son semejantes a las areolas marginales del resto de la valva (Fig. 20 F-G), mientras que en P. elongatum las areolas tienen forma de flor y en P. amara las areolas tienen forma de ojal y se hallan en un área deprimida (ver Stidolph 1992). El material tipo de P. delicatulum no ha sido analizado aún, por lo que no se pudieron establecer comparaciones más allá del contorno y de los parámetros morfométricos. Pleurosigma brunii es una especie mal conocida, con un ángulo del rafe mayor y una mayor relación largo/ancho de la valva que P. perlucidum. El material tipo de P. brunii no pudo ser localizado ya que no aparece mencionado en el catálogo de la Colección de diatomeas del mundo entero de Tempère & Peragallo (1907-1915) a pesar de que fue descripta por Peragallo (1891).
Descripción de los nuevos taxa
Recientemente Smith et al. (2011) presentó el texto del Art. 36.3 del Código Internacional de Nomenclatura para algas, fungi y plantas (International Code of Nomenclature for algae, fungi, and plants, ICN) aprobado durante la Sección de Nomenclatura del XVIII Congreso Internacional de Botánica que tuvo lugar en Melbourne, Australia. Dicho artículo puntualiza que a fin de que el nombre de un nuevo taxon de cualquier organismo cubierto por el Código esté válidamente publicado, a partir del 1º de Enero de 2012 debe estar acompañado por una descripción o diagnosis en Latín o en Inglés. De acuerdo con la prescripción del Art. 36.3 del ICN, las descripciones formales son presentadas debajo, en Inglés.
Pleurosigma patagonicum (Ferrario & Sar) Sterrenburg & Sar var. paucistriatum Sar, Sterrenburg & Sunesen nov. var. (Figs. 15 A-F, 16 A-F, 17 A-F, Table 1)
Description
Light-microscopy
Valves narrowly lanceolate to subrhomboidal, straight, with acute apices slightly vaulted at the level of the raphe-sternum, 132-203 mm long, 13-26 mm wide. Raphe sternum straight with an abrupt deflection to the valvar margin, in opposite directions, at the poles. Terminal areas bilaterally dilated, symmetric, funnel-shaped, in apical position. Raphe angle 0º. Central area elliptical, approximately symmetric. Striae transverse 19-23 in 10 μm, oblique 17-21 in 10 μm, intersection angle of the oblique striae 55-66º.
Scanning-electron microscopy
The raphe only becomes eccentric in its most distal portion. Central raphe fissures undulating in the same sense, with very approximate or just overlapping terminations. Apical terminations of the raphe obliquely positioned, ending as hooks near the valve margin. Raphe sternum internally thickened. Central node elliptical, bordered by two symmetric bars. Central area in the form of a small platform around the bars that surround the central node. Central raphe endings coaxial. Helictoglossae slightly oppositely deflected, surrounded by a smooth proximal area, delimited towards the valvar margin by a slightly depressed area with a row of eyelet-shaped transapically enlarged areolae, externally visible as two small apical fissures ("calcars"). Apical structure with an isolated pore next to the helictoglossae. Areolae loculate arranged in decussate rows, with slitshaped external foramina and elliptic internal pores occluded by a hymen crossed by a bar.
Areolae located in a transapical rectangle around the central node and a short portion of the raphe sternum with occluded, circular, pores without bars. Holotype: slide LPC 13671 (3) here designated, labelled "holotipo de Pleurosigma patagonicum var. paucistriatum, Piedras Coloradas, 05/08/06", deposited in the Herbarium of the División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Isotype: slide LPC 13671 (4) here designated, labelled "isotipo de Pleurosigma patagonicum var. paucistriatum, Piedras Coloradas, 05/08/06", deposited in the Herbarium of the División Ficología, Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Type locality: Piedras Coloradas, 40º 50' 33''S-65º 06' 54''W, Prov. de Río Negro, Argentina. Etymology of the name: this variety of P. patagonicum received the epithet "paucistriatum" from the Latin, pauci: few (short in number) and strãtum: striated, to denote that has lower stria density than the type variety.
Pleurosigma perlucidum Sar, Sterrenburg & Sunesen nov. spec. (Figs. 18 A-H, 19 A-H, 20 A-F, Table 2).
Description
Light-microscopy Valves
very delicate, vaulted at the level of the raphe sternum, linear-lanceolate, clearly sigmoid, with acute apices, 258-322 μm long, 23-29 μm wide. Raphe sternum sigmoid, approximately central, deflected oppositely near the poles. Raphe angle +9º to +11º. Central area round, small, terminal areas small, funnel-shaped, in apical position. Striae transverse 29-30 in 10 μm, oblique 25-28 in 10 μm, intersection angle of the oblique striae 58-63º at the centre and 52-58º at the poles.
Scanning-electron microscopy
Central raphe fissures undulating in opposite directions, more or less overlapping in the terminal portions. Apical raphe fissures hook-shaped, oppositely deflected towards the concave side of the valve. Isolated pore placed at the apices on the opposite side of the terminal raphe fissures. Raphe sternum straight, internally thickened. Central nodule elliptic, bordered by two symmetric bars. Central raphe endings coaxial, slightly dilated in the form of a pore. Apical raphe endings in the form of helictoglossae, slightly oppositely deflected, surrounded by a smooth area. Marginal areolae interrupted at the end of the helictoglossa on the concave side of the valve and below it on the convex side, except for one or two isolated areolae near to its end. Areolae loculate in decussate rows, external foramina slit-shaped and internal pores elliptic, occluded by a more or less depressed hymen crossed by a bar. Areolae located around the central node and a short portion of the raphe sternum with circular occluded pores, without bars.
Holotype: slide LPC 4260 (3) here designated, labelled "holotipo de Pleurosigma perlucidum, Nueva Atlantis, 27/03/1995", deposited in the Herbarium of the División Ficología "Dr. Sebastián A. Guarrera", Facultad de Ciencias Naturales y Museo, La Plata, Argentina. Isotype: slide LPC 4260 (4) here designated, labelled "isotipo de Pleurosigma perlucidum, Nueva Atlantis, 27/03/1995", deposited in the Herbarium of the División Ficología, Facultad de Ciencias Naturales y Museo, La Plata, Argentina.
Type locality: Nueva Atlantis, 36º 45' 81''S-56º 40' 04''W, Prov. de Buenos Aires, Argentina.
Etymology of the name: this species received the epithet "perlucidum" from the Latin, to denote that the valves are completely transparent under light microscopy with brightfield illumination.
Agradecimientos
Esta investigación fue financiada mediante subsidios de la Universidad Nacional de La Plata 11/N640 y del CONICET PIP0067.
Bibliografía
1. BOALCH, G. T. & D. S. HARBOUR. 1977. Observations on the structure of a planktonic Pleurosigma. Nova Hedwigia, Beih. 54: 275-280. [ Links ]
2. CARDINAL, A., M. POULIN & L. BÉRARDTHERRIAULT. 1989. New criteria for species characterization in the genera Donkinia, Gyrosigma and Pleurosigma (Naviculaceae, Bacillariophyceae). Phycologia 28: 15-27. [ Links ]
3. CLEVE, P. T. 1894. Synopsis of the Naviculoid Diatoms, Part I. Bih. Kongl. Svenska Vetensk.-Akad. Handl. 26: 1-194. [ Links ]
4. FERRARIO, M. E. & E. A. SAR. 1990. Ultraestructura de Pleurosigma chilensis var. patagonica nov. var. (Bacillariophyceae). Darwiniana 30: 201-204. [ Links ]
5. FERRARIO, M. E., E. A. SAR, C. CASTAÑOS & F. HINZ. 1999. Potentially toxic species of the diatom genus Pseudo-nitzschia in Argentinian coastal waters. Nova Hedwigia 68: 131-147. [ Links ]
6. FERRARIO, M. E., E. A. SAR & S. E. SALA. 1995. Metodología básica para el estudio del fitoplancton con especial referencia a las diatomeas. En: ALVEAL, K., M. E. FERRARIO, E. C. OLIVEIRA & E. SAR (eds.), Manual de Métodos Ficológicos, pp. 1-23. Universidad de Concepción, Concepción, Chile. [ Links ]
7. FRENGUELLI, J. 1928. Diatomeas del Océano Atlántico frente a Mar del Plata (República Argentina). Anales Mus. Nac. Hist. Nat. "Bernardino Rivadavia" 34, Protistología 1: 497-572. [ Links ]
8. FRENGUELLI, J. 1930. Contribución al conocimiento de las diatomeas argentinas. VI Diatomeas marinas de la costa atlántica de Miramar (Provincia de Buenos Aires). Anales Mus. Nac. Hist. Nat. "Bernardino Rivadavia" 36, Protistología 2: 243-311. [ Links ]
9. FRENGUELLI, J. 1938. XII Contribución al conocimiento de las diatomeas argentinas. Diatomeas de la Bahía de San Blas. (Provincia de Buenos Aires). Revista Mus. La Plata (n. s.) 1, Bot. 5: 251-337. [ Links ]
10. FRENGUELLI, J. 1939. XV Contribución al conocimiento de las diatomeas argentinas. Diatomeas del Golfo de San Matías (Río Negro). Revista Mus. La Plata (n. s.) 2, Bot. 10: 201-226. [ Links ]
11. GAYOSO, A. M. 1988. Variación estacional del fitoplancton de la zona más interna del estuario de Bahía Blanca (Prov. Buenos Aires, Argentina). Gayana Bot. 45: 241-247. [ Links ]
12. GAYOSO, A. M. 1989. Species of the diatom genus Thalassiosira from a coastal zone of the South Atlantic (Argentina). Bot. Mar. 32: 331-337. [ Links ]
13. HARPER, M. A., J. E. PATTERSON & J. F. HARPER. 2009. New diatom taxa from the world's first marine Bioblitz held in New Zealand: Skeletomastus a new genus, Skeletomastus coelatus nov. comb. and Pleurosigma inscriptura a new species. Acta Bot. Croat. 68: 339-349. [ Links ]
14. HASLE, G. R. & G. A. FRYXELL. 1970. Diatoms: cleaning and mounting for light and electron microscopy. Trans. Amer. Microscop. Soc. 89: 469- 474. [ Links ]
15. HASLE, G. R. & E. E. SYVERTSEN. 1996. Marine diatoms. In: TOMAS, C. R. (ed.), Identifying Marine Phytoplankton, pp. 5-385. Academic Press, San Diego, California. [ Links ]
16. HENDEY, N. I. 1964. An introductory account of the smaller algae of British coastal waters. Part V. Bacillariophyceae (Diatoms). Her Majesty Stationery Office, London. [ Links ]
17. HUSTEDT, F. 1955. Marine littoral diatoms of Beaufort, North Carolina. Bull. Duke Univ. Mar. Sta. 6: 1-67. [ Links ]
18. LANGE, C., R. M. NEGRI, H. R. BENAVIDES. 1983. Algunas especies del género Thalassiosira (Bacillariophyceae) del Mar Argentino. (Parte 1). Iheringia, Bot. 31: 9-30. [ Links ]
19. NEGRI, R. M. & D. INZA. 1998. Some potentially toxic species of Pseudo-nitzschia in the Argentine Sea (35º-39ºS). In: REGUERA, B., J. BLANCO, M.L. FERNANDEZ & T. WYATT (eds.), Harmful Algae, pp. 84-85. Xunta de Galicia and IOC of UNESCO, Santiago de Compostela. [ Links ]
20. PERAGALLO, H. 1891. Monographie du genre Pleurosigma et des genres alliés. Extrait du Diatomiste 1890-1891. Tempère, J., Paris. [ Links ]
21. PRITCHARD, A. 1861. A history of Infusoria, including the Desmidiaceae and Diatomaceae British and foreign. Fourth edition, enlarged and revised by Arlidge, J. T., W. Archer, J. Ralfs, W. C. Williamson and the author. Whittaker & Co, London. [ Links ]
22. PRYGIEL, J. & M. COSTE. 2000. Guide Méthodologique pour la mise en ouvre de l´Indice Biologique Diatomées. Agences de l'Eau, Ministère de l'Aménagement du Territoire et de l'Environnement, Direction de l'Eau & CEMAGREF, France. [ Links ]
23. REID, G. 2001. On the identity of Pleurosigma angulatum (Bacillariophyta) and related species. Bull. Nat. Hist. Mus. London, Bot. 31: 107-117. [ Links ]
24. REID, G. 2002. Four new species of Pleurosigma (Bacillariophyta) from Alexandria, Egypt. Bot. J. Linn. Soc. 140: 77-92. [ Links ]
25. REID, G. 2012. A revision of the family Pleurosigmataceae (Bacillariophyta). Diatom Monographs 14. A.R.G. Gantner Verlag K. G. Ruggell, Liechtenstein. [ Links ]
26. ROSS, R., E. J. COX, N. I. KARAYEVA, D. G. MANN, T. B. B. PADDOCK, R. SIMONSEN & P.A. SIMS. 1979. An amended terminology for the siliceous components of the diatom cell. In: SIMONSEN, R. (ed.), Proceedings of the Fifth Symposium on Recent and Fossil Marine Diatoms. Nova Hedwigia, Beih. 64: 513-533. [ Links ]
27. ROUND, F. E., R. M. CRAWFORD & D. G. MANN. 1990. The Diatoms. Morphology and Biology of the Genera. Cambridge University Press, London. [ Links ]
28. SAR, E. A. 1989. Ultrastructure of Odontella rostrata (Hustedt) Simonsen, a new record for the South Atlantic Ocean. Bot. Mar. 32: 187-189. [ Links ]
29. SAR, E. A. 1996a. Flora diatomológica de Bahía San Antonio (Prov. de Río Negro, Argentina). O. Centrales I. Revista Mus. La Plata (n. s.) 14, Bot. 106: 365-400. [ Links ]
30. SAR, E. A. 1996b. Flora diatomológica de Bahía San Antonio (Prov. de Río Negro, Argentina). O. Pennales I. Revista Mus. La Plata (n. s.) 14, Bot. 107: 399-432. [ Links ]
31. SAR, E. A. & M. E. FERRARIO. 1990. Licmophora flabellata: ultrastructure and taxonomy I. Implication. Diatom Res. 5: 403-408. [ Links ]
32. SAR, E. A. & I. SUNESEN. 2003. Nanofrustulum shiloi (Bacillariophyceae) from Gulf of San Matías (Argentina): morphology, distribution and comments about nomenclature. Nova Hedwigia 77: 399-406. [ Links ]
33. SAR, E. A., D. U. HERNÁNDEZ BECERRIL & I. SUNESEN. 2002a. A morphological study of Chaetoceros tenussimus Meunier, a little known planktonic diatom, with a discussion of the Section Simplicia, Subgenus Hyalochaete. Diatom Res. 17: 327-335. [ Links ]
34. SAR, E. A., F. HINZ, F. A. S. STERRENBURG, A. S. LAVIGNE, S. LOFEUDO & I. SUNESEN. 2012. Species of Pleurosigma (Pleurosigmataceae) with lanceolate or slightly sigmoid valve outline. Analysis of type materials. Diatom Res. 27: 237-253. [ Links ]
35. SAR, E.A., O. ROMERO & I. SUNESEN. 2003. Cocconeis Ehrenberg and Psammococconeis Garcia (Bacillariophyta) from the Gulf of San Matías, Patagonia, Argentina. Diatom Res. 18: 79-106. [ Links ]
36. SAR, E. A., I. SUNESEN & C. CASTAÑOS. 2001. Marine diatoms from Buenos Aires coastal waters (República Argentina). I. Thalassiosiraceae. Nova Hedwigia 73: 199-228. [ Links ]
37. SAR, E. A., I. SUNESEN & A. S. LAVIGNE. 2002b. The diatom genus Thalassiosira: species from the northern San Matías Gulf (Río Negro, Argentina). Nova Hedwigia 74: 373-386. [ Links ]
38. SAR, E.A., I. SUNESEN & P.V. FERNÁNDEZ. 2007a. Marine Diatoms from Buenos Aires coastal waters (Argentina). II. Thalassionemataceae and Rhaphoneidaceae. Rev. Chil. Hist. Nat. 80: 63-79. [ Links ]
39. SAR E. A., I. SUNESEN & A. S. LAVIGNE. 2010a. Cymatotheca, Tryblioptychus, Skeletonema and Cyclotella (Thalassiosirales) from Argentinian coastal waters. Description of Cyclotella cubiculata sp. nov. Vie Milieu 60: 135-156. [ Links ]
40. SAR, E. A., I. SUNESEN & R. JAHN. 2010b. Coscinodiscus perforatus revisited and compared with Coscinodiscus radiatus (Bacillariophyceae). Phycologia 49: 514-524. [ Links ]
41. SAR, E. A, I. SUNESEN, R. JAHN, W. H. KUSBER & A. S. LAVIGNE. 2007b. Revision of Odontella atlantica (Frenguelli) Sar comb. et stat. nov. with comparison to two related species, O. rhombus (Ehrenb) Kütz. and O. rhomboides R. Jahn et Kusber. Diatom Res. 22: 341-353. [ Links ]
42. SAR, E. A., I. SUNESEN, A. S. LAVIGNE & S. LOFEUDO. 2011. Thalassiosira rotula Meunier a heterotypic synonym of Thalassiosira gravida: morphological evidence. Diatom Res. 26: 109-119. [ Links ]
43. SHADBOLT, G. 1853. A short description of some new forms of Diatomaceae from Port Natal. Trans. Microscop. Soc. London (n. s.) 2: 13-18. [ Links ]
44. SIMONSEN, R. 1974. The diatom plankton of the Indian Ocean Expedition of R/V "Meteor" 1964- 1965. "Meteor Forschungsergebnisse". Gebrüder Borntraeger, Reihe D, 19. Berlin, Stuttgart. [ Links ]
45. SIMONSEN, R. 1987. Atlas and Catalogue of the Diatom Types of Friedrich Hustedt. J. Cramer, Berlin, Stuttgart. [ Links ]
46. SMITH, G. F., E. FIGUEIREDO & G. MOORE. 2011. English and Latin as alternative languages for validating the names of organisms covered by the International Code of Nomenclature for algae, fungi, and plants: The final chapter?. Taxon 60: 1502-1503. [ Links ]
47. SMITH, W. 1852. Notes on the Diatomaceae; with description of British species included in the genus Pleurosigma. Ann. Mag. Nat. Hist. (ser. 2) 9: 1-12. [ Links ]
48. SMITH, W. 1853. A synopsis of the British Diatomaceae; with remarks on their structure, functions and distribution; and instructions for collecting and preserving specimens. Vol. 1. John Van Voorst, London. [ Links ]
49. STERRENBURG, F. A. S. 1990. Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae). A new phenomenon: co-existence of dissimilar raphe structure in population of several species. In: RICARD, M. (ed.), Ouvrage dédié à la Memoir du Professeur Henry Germain (1903-1989), pp. 235- 242. Koeltz, Koeningstein. [ Links ]
50. STERRENBURG, F. A. S. 1991a. Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae). Light microscopical criteria for taxonomy. Diatom Res. 6: 367-389. [ Links ]
51. STERRENBURG, F. A. S. 1991b. Studies on the Genera Gyrosigma and Pleurosigma (Bacillariophyceae). The typus generis of Pleurosigma, some presumed varieties and imitative species. Bot. Mar. 34: 561- 573. [ Links ]
52. STERRENBURG, F. A. S. 2001. Studies on the Genera Pleurosigma and Gyrosigma. The types of Shadbolt and related taxa. P. Acad. Nat. Sci. Phila. 151: 121- 127. [ Links ]
53. STERRENBURG, F. A. S. 2003. Studies on the diatom genera Gyrosigma and Pleurosigma (Bacillariophyceae): Pleurosigma strigosum W. Smith and some presumptive relatives. Micropaleontology 49: 159-169. [ Links ]
54. STERRENBURG, F. A. S., M. E. MEAVE DEL CASTILLO & M. A. TIFFANY. 2003. Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae): Pleurosigma species in the plankton from the Pacific coast of Mexico, with the description of P. gracilitatis sp. nov. Cryptogamie, Algol. 24: 291-306. [ Links ]
55. STERRENBURG, F. A. S., I. SUNESEN & E. A. SAR. (enviado). Revision of Pleurosigma formosum and P. decorum (Pleurosigmataceae, Bacillariophyta) using type material. Phycol. Res. [ Links ]
56. STIDOLPH, S. R. 1992. Observations and remarks on the morphology and taxonomy of the diatom genera Gyrosigma Hassall and Pleurosigma W. Smith III. Gyrosigma sterrenburgii sp. nov., and Pleurosigma amara sp. nov. Diatom Res. 7: 345-366. [ Links ]
57. STIDOLPH, S. R. 2002. Observations and remarks on the morphology and taxonomy of the diatom genera Gyrosigma Hassall and Pleurosigma W. Smith. V. Pleurosigma types of A. Mann (1925): a critical reinvestigation. Micropaleontology 48: 273-284. [ Links ]
58. SUNESEN, I. & E. A. SAR. 2004. Thalassiosira fryxelliae nov. spec. (Bacillariophyceae) from Argentinian coastal waters. Bot. Mar. 47: 238-247. [ Links ]
59. SUNESEN, I. & E. A. SAR. 2007a. Diatomeas marinas de aguas costeras de la Provincia de Buenos Aires (Argentina). III Géneros potencialmente nocivos Asterionellopsis, Cerataulina, Ceratoneis y Leptocylindrus. Rev. Chil. Hist. Nat. 80: 493-507. [ Links ]
60. SUNESEN, I. & E. A. SAR. 2007b. Marine diatoms from Buenos Aires coastal waters (Argentina). IV. Rhizosolenia s. str., Proboscia, Pseudosolenia, Neocalyptrella. Phycologia 46: 628-643. [ Links ]
61. SUNESEN, I., A. BÁRCENA & E. A. SAR. 2009. Diatomeas potencialmente nocivas del Golfo San Matías (Argentina). Rev. Biol. Mar. Oceanogr. 44: 67-88. [ Links ]
62. SUNESEN, I., D. U. HERNÁNDEZ-BECERRIL & E. A. SAR. 2008. Marine diatoms from Buenos Aires coastal waters (Argentina). V. Species of the genus Chaetoceros. Rev. Biol. Mar. Oceanogr. 43: 303-326. [ Links ]
63. TEMPÈRE, J. & H. PERAGALLO. 1907-1915. Diatomées du Monde Entier. Collection Tempère et Peragallo. Edition 2. Arcachon, Grez-sur-Loing. [ Links ]
64. VANLANDINGHAM, S. L. 1978. Catalogue of the fossil and recent genera and species of Diatoms and their synonyms. Part VI: Neidium through Rhoicosigma. J. Cramer, Vaduz. [ Links ]
65. VOUILLOUD, A. A. 2003. Catálogo de diatomeas continentales y marinas de Argentina. Asociación Argentina de Ficología, La Plata. ISBN nº 987- 21041-0-7. [ Links ]
Recibido el 11 de abril de 2012,
aceptado 25 de septiembre de 2012.
