










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.49 no.1 Córdoba mar. 2014
SISTEMÁTICA DE PLANTAS VASCULARES
La vegetación reófila del Nordeste argentino. Las comunidades vegetales con Podostemaceae de la Provincia de Misiones
José Luis Fontana1
1Laboratorio de Ecología Vegetal, Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, Av. Libertad 5470, 3400-Corrientes. E-mail: jlfontana@yahoo.com.ar
Resumen
En la provincia de Misiones, comunidades vegetales con Podostemáceas viven en rápidos y saltos de los arroyos pertenecientes a dos cuencas (Ríos Paraná y Uruguay). La Comunidad de Mourera aspera se encuentra sólo en las correderas río arriba de las Cataratas del Iguazú. Sobre la roca submersa en correderas crece una Comunidad con Apinagia yguazuensis, relativamente rara, encontrada en el Río Iguazú y en los arroyos Tabay y Yazá. En paredes y al pié de saltos y en determinados rápidos, comunidades con Podostemum spp. constituyen la vegetación reófila más llamativa: la Comunidad de Podostemum comatum en Cataratas, las Comunidades de Podostemum mulleri y la de Podostemum atrichum exclusivas de arroyos de la cuenca del Paraná, las Comunidades de Podostemum rutifolium y la de P. undulatum de amplia distribución en las cuencas de los Ríos Paraná y Uruguay. Comunidades reófilas con Podostemum uruguayense son exclusivas de la cuenca del Río Uruguay. La comunidad con Marathrum azarensis si bien en Misiones sólo fue encontrada en afluentes del río Uruguay, crece también en el Río Paraná en la provincia de Corrientes. Se analizaron las condiciones del hábitat y la distribución de las comunidades según cuencas.
Palabras clave: Vegetación reófila; Podostemaceae; Hábitat; Misiones; Argentina.
Summary
Northeastern Argentine rheophile vegetation. Plant communities with Podostemaceae of the Misiones Province. In the Misiones province, plants communities with Podostemaceae live in rapids and waterfalls of the streams beloging to two watersheds (Paraná and Uruguay rivers). The Mourera aspera Community is only found in the tracks going upstream the Iguazú Falls. On the rock submerged in tracks there grows a Community with Apinagia yguazuensis, relatively rare, and only found in the Iguazú river and in the streams Tabay and Yazá. On walls and at the foot of waterfalls and in certain rapids, communities with Podostemum spp. constitute the most singular rheophile vegetation: the Podostemum comatum Community in Cataratas, the Podostemum mülleri and Podostemum atrichum Communities peculiar to streams of the Paraná watershed, the Podostemum rutifolium and P. undulatum Communities of wide distribution in the watersheds of the Paraná and Uruguay rivers. The community Marathrum azarensis although it was only found in tributaries of the River Uruguay, also grows in the Parana River in the Corrientes province. Habitats conditions were analysed and the distribution of the communities as relates to watersheds was studied.
Key words: Rheophile vegetation; Podostemaceae; Habitat; Misiones; Argentine.
Introducción
Los arroyos de la provincia de Misiones forman correderas y saltos donde es posible encontrar comunidades vegetales constituidas por plantas reófilas, sometidas a la fuerte tracción del agua. El hábitat particular determina formas de crecimiento características, con predominancia de plantas de hojas divididas, largas y flexibles, que evitan roturas por la fuerza del agua.
Llaman la atención por su aspecto las Podostemáceas, morfológicamente muy variables, desde aquellas con grandes hojas rígidas (Mourera), hasta plantas parecidas a algas con hojas finamente divididas (Apinagia, Podostemum, Marathrum), y otras parecidas a musgos (Tristicha). Se trata de una familia de plantas esencialmente tropicales (Schnell in Landolt et al., 1998).
Por lo general de un verde oscuro, estas plantas cuelgan de las paredes de los saltos y represas o están fijas a rocas de las correderas, donde la corriente del agua orienta sus tallos y hojas río abajo. Podostemum es el género más común; cubre la roca o el cemento de las represas en forma de manto, constituyendo comunidades vegetales que en la mayoría de los sitios están representadas sólo por una o dos especies del género, acompañadas por Tristicha. Mourera crece actualmente sólo en el Río Iguazú; antes de la construcción de la represa de Yacyretá, también existía en Ituzaingó, provincia de Corrientes. Apinagia fue encontrada en cuatro sitios de la provincia de Misiones: en Cataratas, en el Salto del arroyo Urugua-í y en dos lugares del centro provincial (Fontana, 2008). Marathrum citado para Argentina por Tur (2003) vive en aguas corrientes más o menos tranquilas, por lo general poco profundas (10 a 15 cm, excepcionalmente hasta 30 cm) en arroyos del sur misionero, en el límite con la provincia de Corrientes, y al pié de la represa de Yacyretá (Fontana, 2007).
Para el nordeste argentino, los conocimientos actuales sobre la vegetación reófila y particularmente sobre comunidades con predominancia de Podostemaceae son muy generales y en algunos aspectos, como hábitat y composición florística, prácticamente inexistentes. Fontana (2008a) describe aspectos ecológicos de Apinagia yguazuensis; Fontana & Iriart (en prensa) analizan detalladamente la vegetación de correderas y de islotes en el Parque provincial Moconá (prov. de Misiones); Eskuche (1986) menciona la presencia de una comunidad vegetal formada por Paspalum lilloi y Podostemum atrichum en las correderas del Río Iguazú, sobre los saltos; Drehwald (1990) describe una comunidad con Podostemáceas y musgos, estableciendo una asociación, el Podostemetum undulati, en base a censos realizados en el Salto Berrondo (arroyo Salto), departamento Oberá, diferenciando las subasociaciones typicum y tristichietosum trifariae; Carnevali (1994) cita especies de los rápidos de Apipé y comunidades vegetales de la orilla inundable de arroyos del norte y este de Corrientes. Trabajos sistemáticos con breves comentarios ecológicos fueron publicados por Tur (1997, 1999, 2003). La morfología y la anatomía de plantas que viven bajo condiciones de fuerte tracción del agua fueron analizadas por Ancibor (1990).
Para otras regiones de América dan información Chodat & Vischer (1917) quienes describen plantas de cascadas y saltos del Paraguay y de las Cataratas del Iguazú. Para Brasil, Hoehne (1948) informa sobre la ecología de reófitos; Van Royen & Reitz (1971) describen las Podostemáceas de Santa Catarina con breves comentarios sobre condiciones de vida; Klein (1979) enumera los reófitos más comunes, con datos sobre hábitat y abundancia. Novelo & Philbrick (1997) describen las Podostemáceas de México con comentarios sobre la distribución. Fontana (2007) menciona por primera vez Marathrum azarensis para Paraguay, haciendo referencia al hábitat de la especie; Tur (2009) completa la lista de Podostemáceas para la Flora del mismo país.
Philbrick & Novelo (1995) dan un panorama sobre los estudios taxonómicos y ecológicos de las Podostemáceas, problemas de conservación y sugerencias para la colección de las mismas, y en 2004 presentan un trabajo monográfico sobre el género Podostemum. Schnell (1970) trata los reófitos de las regiones tropicales. Van Steenis (1981), analiza el hábitat, el problema de la evolución, la morfología y la ecología de los reófitos del mundo. Gruber (1991) estudió las fanerógamas presentes en los saltos de regiones tropicales y Schnell (1994) las adaptaciones de las plantas que viven en ríos torrentosos. Ruthishauser (1997) trata la diversidad estructural y del desarrollo de las Podostemáceas.
Con respecto a las condiciones químicas de las aguas y la relación con la distribución de las Podostemaceae, Quiróz et al. (1997) informan sobre arroyos mexicanos.
Las comunidades vegetales con Podostemáceas abrigan y dan alimento a diversos animales. Algunas especies de pirañas herbívoras (Tometes spp., Serrasalmidae) que se encuentran en arroyos, desde las Guayanas hasta Paraguay, dependen de estas plantas (Guillaume, 2003). En el Alto Paraná muchos peces herbívoros también se alimentan con Podostemum (Bechara, 2005, com.verbal). Silveira Vilella & al. (2002) consideran a las Podostemáceas como fuente importante de alimentación de Astyanax spp. (Characidae), peces omnívoros del SE de Brasil. En Misiones, moluscos y peces fueron observados escondidos en el césped submerso formado por Podostemáceas (Chatellenaz, 2007).
Los simúlidos encontrarían en estas plantas protección para sus larvas (Hamada, 2000; Sprada Tavares, 2006). Un elevado número de larvas pertenecientes a Ephemeroptera y Trichoptera, y en menor medida a Odonata y Megaloptera encuentran buenas condiciones de vida (alimento y protección) en las hojas de Mourera fluviatilis (Fleck, 2004). Zambiasio et al. (2011, 2012) analizaron en dos arroyos de la provincia de Misiones las comunidades de invertebrados asociados a Podostemum mulleri y P. atrichum, encontrando Megaloptera, Odonata, Plecoptera, Coleoptera, Ephemeroptera, Díptera, Trichoptera, Lepidoptera y Platelmintos (Tricladida, Planaria). Sobre las utilidades de las Podostemáceas informan Philbrick & Novelo (1995, 2004). Aborígenes de Amazonia usan las hojas disecadas y pulverizadas como sustituto de la sal en sus comidas. En México se utiliza Marathrum sp. para afecciones hepáticas. Los usos en cosmética de productos hechos a partir de Mourera fluviatilis como hidratantes de la piel, también son conocidos (es.clarins.com). Indígenas de Guayana hacen una tortilla con hojas de tabaco condimentada con Mourera para mascar (Wilbert, 2000).
Materiales y Métodos
Para el estudio de la vegetación se emplearon los métodos analíticos y sintéticos de la escuela fitosociológica de Braun-Blanquet descriptos entre otros por Braun-Blanquet (1979) y Dierschke (1994). Para la identificación de las plantas se consultaron trabajos sistemáticos, como los de Pontiroli (1955), Van Royen & Reitz (1971), Tur (1997, 1999, 2003, 2009) y Philbrick & Novello (2004). La nomenclatura de las especies sigue a Zuloaga et al. (2008). El material coleccionado se encuentra conservado en alcohol 70%, en el Herbarium Humboldtianum (CTESN).
Los resultados del trabajo sinsistemático se correlacionaron con mediciones hidrológicas y químicas, y con observaciones fenológicas. Para la determinación de los parámetros químicos básicos se utilizó un pH-metro portátil Orion 106 "Quikchek" con una resolución de 0.1 pH y compensación de temperatura; y un conductímetro portátil Orion 116 "Quikchek" con una resolución de 10 mS/cm.
La velocidad de la corriente del agua se determinó mediante la medición del tiempo que tardaba una boya plástica de pesca de 4 cm de diámetro en recorrer 10 m de distancia.
Condiciones ambientales del área de estudio
Hidrografía
Los arroyos misioneros pertenecen a dos grandes cuencas (Fig. 1), la del Río Uruguay al E y la del Río Paraná al W. El Río Iguazú, en el límite norte de la provincia, es el afluente más importante del Paraná. A los tres grandes ríos que rodean la provincia desembocan unos 800 cursos de agua; de ellos 120 van a los ríos Iguazú y San Antonio, 270 al Río Paraná y 400 a los ríos Uruguay y Pepirí Guazú. Los arroyos que desembocan en el Río Iguazú, a excepción del río San Antonio, son pequeños (Castellanos, 1975), y en algunos casos temporarios.

Fig. 1. Cuencas y principales subcuencas de la Provincia de Misiones, con indicación de los sitios relevados. Aº: Arroyo. Cuenca del Río Paraná: 1-Garupá, 2-S.Juan, 3-Sta. Ana, 4-Yabebiry, 5-Ñacanguazú, 6-Tabay, 7-Cuñapirú, 8-Capioví, 9-Garuhapé, 10-Paranay-Guazú, 11-Itacuruzú, 12-Piray Guazú, 13-Piray Miní, 14-Aguaray Guazú, 15-Urugua-í, 16-Mbocay. Subcuenca del Río Iguazú: 17-Sto.Domingo, 18-Yacuy, 19-S.Francisco, 20-Deseado, 21-S.Antonio. Cuenca del Río Uruguay: 22-Pepirí Guazú, 23-Pepirí Miní, 24-Paraíso, 25-Soberbio, 26-Chafariz, 27-Saltito, 28-Pindaití, 29-Canal Toro, 30-Acaraguá, 31-Ramón, 32-Chico Alférez, 33-Once Vueltas, 34-Guerrero López, 35-Itacaruaré, 36-Sta.María, 37-Persiguero, 38-Concepción, 39-Tunas, 40-Chimiray.
Los ríos Iguazú, Paraná y Uruguay son de régimen subtropical. Si bien las precipitaciones están distribuidas en todo el año, ocurren dos máximos, uno en otoño y otro en primavera, que determinan dos crecientes anuales. El régimen de escurrimiento de los arroyos y ríos del territorio provincial, debido a que poseen por lo común recorridos cortos, varía considerablemente, incluso en un mismo día, porque dependen de las lluvias que a veces se producen sólo en determinadas subcuencas.
Las Sierras del Imán y de Misiones constituyen la divisoria de aguas entre la cuenca del Paraná y la del Uruguay. Hacia uno u otro lado de la Sierra de Misiones, el relieve accidentado determina la formación de correderas y de numerosos saltos, algunos de ellos con alturas superiores a 25 m, como los saltos Paca (50 m), Encantado (48 m), Sak 1 (36 m), entre otros (Mercanti, 2007).
En el N de la provincia, la Sierra de la Victoria delimita la subcuenca del Río Iguazú. Dos accidentes geológicos destacan en la provincia, las Cataratas del Iguazú y el Salto del Moconá en el Río Uruguay. Ambos grupos de saltos constituyen ambientes propicios para el desarrollo de la vegetación reófila, particularmente de las Podostemáceas.
Clima
El clima de Misiones puede definirse como subtropical perhúmedo. Siete meses al año la temperatura media es superior a 20 ºC, con máximas absolutas de 42.1 ºC (Posadas, enero y febrero). Las temperaturas mínimas absolutas alcanzan en julio -3.9º C en Cerro Azul (Olinuck, 1998), con posibilidad de heladas entre junio y agosto, -4.9 ºC en Iguazú (SMN, 2010) y -5.6 ºC en Montecarlo, con heladas entre los meses de mayo y septiembre (Olinuck, 1995). Las nevadas, muy esporádicas, son sólo posibles en el extremo NE (Bernardo de Irigoyen) como la ocurrida en julio de 2000, o las de agua nieve en San Pedro (2010 y 2013).
Las precipitaciones son variables; en el norte de la provincia se registró un máximo de 2633 mm y un mínimo de 1655 mm (Parque Provincial Urugua-í, 1998 y 1999 respectivamente).
El invierno es más seco; los meses de julio y agosto registran las menores precipitaciones (Fig. 2). El período más húmedo del año ocurre en el verano, donde las precipitaciones mensuales pueden superar los 200 mm en el norte de la provincia; como límites extremos se registraron 504 mm en abril de 1990 (Montecarlo), 486 mm en octubre de 1998 (Puesto Uruzú, Parque Provincial Urugua-í) y 435 mm para Posadas en octubre de 2000. En general, las precipitaciones disminuyen de E a W y de N a S.
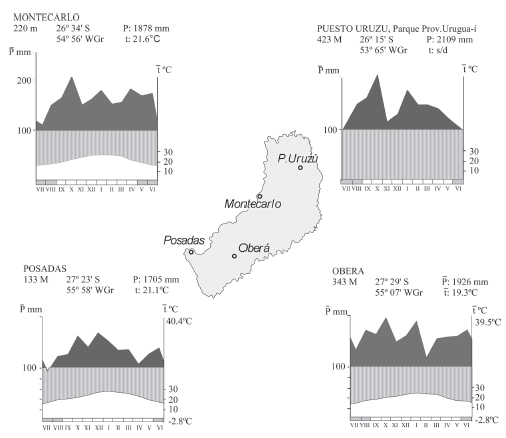
Fig. 2. Diagramas ombrotérmicos correspondientes a estaciones meteorológicas del área de estudio (datos del Servicio Meteorológico Nacional, Ministerio de Ecología y Recursos Naturales Renovables y del INTA).
Habitat
Las comunidades vegetales reófilas con predominancia de Podostemáceas se desarrollan en el cauce de arroyos permanentes con agua de alta transparencia, corriente rápida y por lo general con fuerte turbulencia (Fig. 3).

Fig. 3. Ambientes donde se desarrollan las comunidades de Podostemáceas. A la izquierda, pié del Salto Berrondo. A la derecha, correderas del Río Uruguay en el Parque Provincial Moconá.
Mediante sus hápteros estas plantas se adhieren a la roca basáltica, a veces a piedras sueltas de distinto tamaño, a ladrillos y al cemento de las pequeñas represas construidas en el cauce de los arroyos. El mayor desarrollo se observó en Podostemum undulatum en ejemplares adheridos a paredes verticales de ladrillos (arroyo Salto) o al pié de las caídas (arroyo Persiguero) de los muros de contención del agua, con elevada turbulencia; sus ramas colgantes alcanzan 35-40 cm de longitud. Como parámetros químicos elementales, se realizaron mediciones del pH y de la conductividad en los sitios de relevamiento (Tabla 1). El pH varía entre 6.3 (arroyo Uruzú del Parque Provincial Urugua-í) y 8.6 (arroyo Tabay, en el salto). Los valores más bajos se registraron en sitios sin influencia del hombre, por ejemplo en el arroyo Uruzú que se encuentra dentro del Parque y no existe ningún tipo de explotación ni asentamiento humano en su recorrido; en cambio sobre el arroyo Tabay existen varios balnearios y en el sitio de la toma de muestras, incluso un camping. La conductividad mostró una situación equivalente; los valores extremos obtenidos fueron 20 mS en el arroyo Excelente, un pequeño tributario del arroyo Uruzú, y 130 mS en el arroyo Itacuruzú, aguas arriba del balneario de Colonia Caraguatay.
Tabla 1. Parámetros químicos básicos obtenidos en el agua de comunidades vegetales relevadas. Aº: Arroyo.
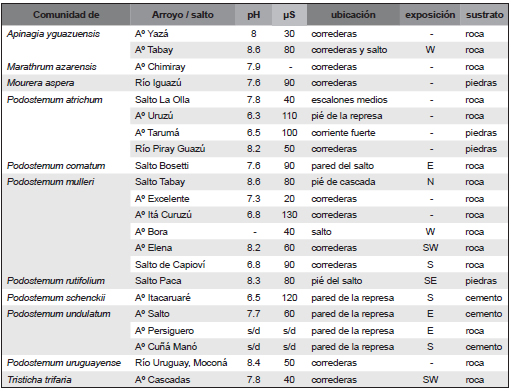
Aparentemente no existen relaciones entre los parámetros químicos básicos (pH y conductividad) y la distribución de las comunidades vegetales. Sí en cambio se observa con respecto a la turbulencia del agua, que estaría relacionada directamente con el contenido de oxígeno. No existen datos referidos a contenido de oxígeno disuelto en el agua. Este elemento tendría importancia para la existencia de estas comunidades vegetales pues se desarrollan en sitios con alta turbulencia y están ausentes en cursos de agua lentos o temporarios.
La velocidad de la corriente del agua varía considerablemente según el nivel del río y la pendiente; con estiaje, en sitios con roca plana y pendiente de 20 % (por ejemplo arroyo Uruzú en el puesto de guardaparques) la velocidad media es de 0,60 m/s, con reducida turbulencia. Con pendientes de hasta 40 %, la velocidad media de la corriente alcanza 1 m/s; la presencia de roca irregular y piedras grandes son responsables de una turbulencia mayor, y constituyen obstáculos que determinan variaciones grandes en la velocidad (por ejemplo Salto Encantado, Salto La Olla), provocando remolinos en el agua.
Resultados
La vegetación reófila
El término reófito fue propuesto por Van Steenis (1932) para las plantas restringidas a correderas y saltos. Estableció dos clases de reófitos: los pertenecientes a una "primera clase" en el que agrupó las plantas fijadas a la roca, que crecen en correderas y cascadas; y los pertenecientes a una "segunda clase", en su mayoría plantas leñosas que resisten correntadas fuertes, ubicadas sobre pequeñas islas o islotes en el cauce de ríos y arroyos y en el valle inundable de los mismos.
Las plantas que integran comunidades de Podostemáceas pertenecen a la primera clase de Van Steenis y las que crecen sobre los islotes a la denominada segunda clase (Fontana & Iriart, manuscrito).
La vegetación limitada a correderas y saltos, siempre bañadas por el agua, se caracteriza por la presencia de plantas especialistas, con predominancia de Podostemáceas, y en algunos sitios de gramíneas y arbustos que crecen en pequeñas islas o promontorios en medio de los cauces de ríos.
Las comunidades con predominancia de Podostemáceas, objeto de esta publicación, viven sobre rocas o piedras submersas y sobre las paredes de los saltos, en cursos de agua permanente. Las comunidades con Podostemum spp. se desarrollan en sitios con gran turbulencia del agua, en saltos, al pié de los mismos o en correderas donde la velocidad del agua es elevada. Las Comunidades de Mourera aspera y la de Apinagia yguazuensis crecen exclusivamente sobre piedras submersas en correderas, pero nunca en los saltos mismos, y rara vez quedan expuestas fuera del agua (Fontana, 2001, 2008).
En los grandes saltos de agua (Cataratas del Iguazú), se encuentra además una comunidad en la que predomina Panicum lilloi, en forma de grandes matas, permanentemente mojadas y sometidas a la fuerte correntada. Algunas comunidades de musgos viven sobre rocas que permanecen la mayor parte del tiempo submersas; estos musgos se caracterizan por la rigidez de sus talos, y por su color, generalmente verde muy oscuro. Drehwald (1990) describió una comunidad integrada por musgos en el arroyo Salto que denominó Sematophylletum riparioides. Sobre islas o islotes y en el valle fluvial de arroyos viven plantas que pasan la mayor parte del tiempo fuera del agua. En islotes periódicamente inundados, como por ejemplo en el Arroyo Piray Guazú, en el Río Uruguay, se encuentra una comunidad vegetal formada por arbustos entre los que predomina Phyllanthus sellowianus de cuyas ramas cuelgan llamativamente líquenes del género Ramalina. Los bordes rocosos de estos islotes albergan grandes matas de Dyckia brevifolia, una llamativa bromeliácea restringida a las cuencas del Río Uruguay y del Río Iguazú (Fontana & Iriart, 2007). En los bordes de los arroyos se desarrollan otras comunidades con gramíneas y alismatáceas, principalmente en los recodos de los arroyos, donde la corriente es más lenta y se depositan sedimentos finos. Echinodorus grandiflorus es la planta que destaca como integrante de una de estas comunidades, por sus grandes hojas e inflorescencias. Los arroyos que corren por valles estrechos o en quebradas tienen pequeñas barrancas donde se observan comunidades formadas por musgos y hepáticas. Las hepáticas predominan en las barrancas formadas por tierra colorada y los musgos principalmente sobre las rocas.
A lo largo de los valles inundables de los arroyos misioneros, la vegetación natural está constituida por franjas de bosque que periódicamente se inundan por horas o unos pocos días, luego de precipitaciones intensas. Esta vegetación arbórea sufrió alteraciones en su estructura y composición por la fuerte presión del hombre (balnearios, construcción de sitios de acampado, tala y fuego), salvo en los parques y reservas.
Las comunidades vegetales con Podostemáceas
1-Comunidad con Mourera aspera
Observada únicamente en las correderas del Ñandú, en el río Iguazú, río arriba de las Cataratas.
Esta comunidad vegetal uniespecífica se desarrolla submersa (rara vez sobresalen las hojas) a una profundidad variable entre ±20 cm durante el estiaje (excepcionalmente menos) y varios metros (3 m o más) durante el período de crecidas (Fig.4). En la bajante de 2006, las plantas estuvieron expuestas, en flor y con muchas hojas en descomposición.
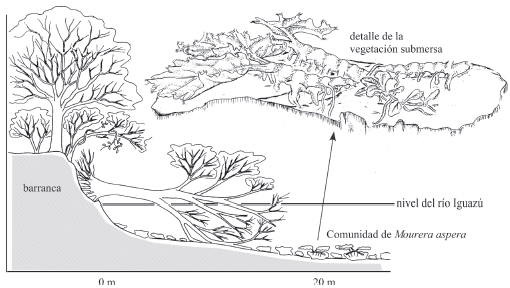
Fig. 4. Perfil de una sección del Río Iguazú ("Correderas del Ñandú") mostrando la ubicación de la Comunidad de Mourera aspera.
Las plantas crecen sobre piedras de distinto tamaño, que reposan sobre el fondo del río formado por arena gruesa. Los tallos rastreros que están fuertemente adheridos por medio de hápteros, muestran un crecimiento a primera vista en zig-zag, debido a las cicatrices alternas que quedaron de las hojas que fueron desprendiéndose. En ejemplares adultos de este hidrófito radicante submerso se observa el progreso de descomposición desde el extremo más viejo de los tallos y la multiplicación vegetativa de la planta a partir de tallos laterales más pequeños.
La cobertura de la comunidad vegetal es baja, no superando 20-30 %. En algunos sitios las plantas cubrían piedras medianas aisladas (30 cm x 15 cm x 5 cm) y en otros, individuos más jóvenes crecían sobre piedras pequeñas juntadas por la fuerte correntada. En uno de los censos (F477), en una superficie de 2.5 m2, se encontraron 7 piedras medianas cubiertas por numerosas plantas.
El área reducida a la que se halla limitada la comunidad vegetal en territorio argentino y la imposibilidad de acceso a otros sitios, no permitió la confección de un mayor número de censos. En los 4 censos fitosociológicos realizados (Tabla 2), Mourera aspera fue la única especie encontrada. La fuerte correntada del río y las características del fondo no permiten el desarrollo de otras plantas.
Tabla 2. Comunidad de Mourera aspera.

Localidad: Ñ, Correderas del Ñandú, Río Iguazú
Una colección de Mourera citada por Tur (1997) indica la presencia de la comunidad también en los rápidos de Apipé, Ituzaingó (provincia de Corrientes), hoy desaparecidos por la construcción de la represa de Yacyretá. La misma autora menciona que la especie sólo fue coleccionada en cuatro oportunidades en territorio argentino.
Sobre esta planta viven numerosos caracoles de reducido tamaño pertenecientes al género Potamolithus (J. Herrera, comunicación verbal), que utilizarían las hojas como alimento; este molusco también fue citado para los rápidos de Apipé, y para los brazos San José y Añacuá del Río Paraná (Varela & Bechara, 1980). Observaciones hechas por los guardaparques del PN Iguazú indicarían que los carpinchos (Hydrochaeris hydrochaeris) también se alimentan de las hojas de Mourera (Garzón, comunicación verbal).
La transparencia del agua depende de las lluvias; el fondo es visible durante períodos de sequía, pero el agua turbia (colorada) que baja después de las lluvias impide ver las plantas. Un pH de 7.6 y una conductividad de 90 mS fueron medidos en las aguas donde se encuentra esta comunidad. Pedralli & Mayer (1996) indican que Mourera aspera puede ser utilizada como bioindicador de agua de buena calidad; mencionan incluso el aumento considerable de ejemplares coincidente con un buen índice de calidad de agua, considerando demanda bioquímica de oxígeno, fosfatos, nitratos, turbidez y oxígeno disuelto.
2-Comunidad de Apinagia yguazuensis
Vegetación submersa que forma un manto denso y apretado de reducida superficie (no más de 0.5 m2) que cubre la roca ± plana, en sitios con fuerte correntada. La cobertura es variable; en algunos sitios cubre totalmente la superficie plana de la roca o bloques de cemento submersos, mientras que en otros crece en forma de plantas aisladas. La mayor cobertura (100%) se observó en parcelas expuestas directamente al sol la mayor parte del día (Salto Tabay, Arroyo Yazá). En estos sitios la planta predominante se observa con facilidad por la coloración amarillo verdoso brillante. En cambio, donde las plantas permanecen la mayor parte del día en la sombra pasan casi desapercibidas por su color verde oscuro, con talos anchos, muy flexibles, con aspecto de hepática. La planta tiene aspecto diferente según la velocidad de la corriente de agua a la que se halla sometida. En aguas rápidas y con insolación directa (arroyo Tabay), la planta es más rígida con hojas finamente divididas y orientadas a favor de la correntada. Las plantas que viven en arroyos de aguas menos turbulenta y corriente menos rápida (arroyo Yazá), tienen hojas anchas, menos rígidas y por lo común son de un verde más oscuro; hacia el borde del haz aparecen pequeños manojos de filamentos característicos de la especie (Fontana, 2008a).
La figura 5 muestra el aspecto de la comunidad. Es posible identificar tres estratos, un estrato sobresaliente que alcanza 5 cm constituido por Apinagia yguazuensis, un estrato intermedio que alcanza 2-3 cm de altura con Podostemum muelleri y P. rutifolium y un estrato muy bajo, pegado a la roca, con Tristicha trifaria, que tiene ramas ascendentes, orientadas en el sentido de la corriente.
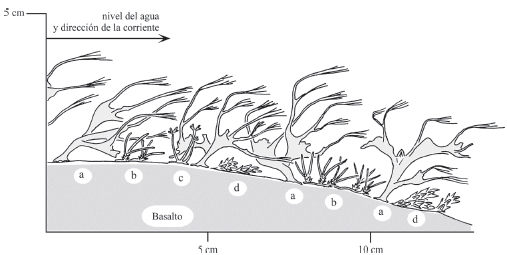
Fig. 5. Perfil de la Comunidad de Apinagia yguazuensis. Salto Tabay, provincia de Misiones. a: Apinagia yguazuensis; b: Podostemum muelleri, c: Podostemum rutifolium; d: Tristicha trifaria.
En los sitios que se encontró la comunidad predomina Apinagia yguazuensis, la única especie del género citada para Argentina (Tabla 3). Esta especie predominante crece junto con Podostemum muelleri, P. rutifolium y Tristicha trifaria.
Tabla 3. Comunidad de Apinagia yguazuensis.
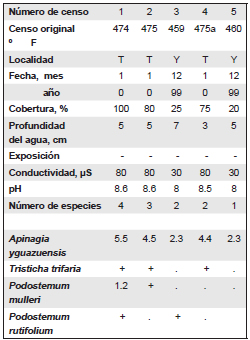
Localidad: T, Salto Tabay; Y, Aº Yazá y Ruta Nacional 14.
Tur (1997) circunscribe su distribución a la Cataratas del Iguazú, restringiendo la especie a la subcuenca del Iguazú. Los censos efectuados en la Provincia de Misiones, amplían su área de distribución aproximadamente 200 km al sur, encontrándose la comunidad en el arroyo Tabay (afluente del río Paraná) y en el arroyo Yazá, afluente del Río Uruguay (Fontana, 2008a). Siempre se observó la comunidad en arroyos con agua transparente, sobre rocas planas, en la parte superior de los saltos, pero nunca en las paredes rocosas o al pié de los saltos. El pH fue básico, variando entre 8 y 8.6. La conductividad en cambio mostró diferencias mayores: 80 mS en arroyo Tabay y 30 mS en el arroyo Yazá.
3-Comunidades con Podostemum spp. y Tristicha trifaria (Tabla 4)
Tabla 4. Comunidades con Podostemum spp. Aº: Arroyo.

Localidades: Arr: Salto Arrechea (PN Iguazú); Ber: A° Salto; Bos: Salto Bosseti; Cap: Salto de Capioví; Car: A° Caraguatá; Cat: Cataratas; Enc: Salto Encantado; Ex: A° Excelente; Pac: Salto Paca; Per: A° Persiguero; Tar: A° Tarumá; Uru: A° Uruzú; PA: A° Piray Guazú y RP 20; PG: A° Piray Guazú, antes de la confluencia con el A° Alegr'a; Bor: A° Bora (Piray).
Tabla 4 (Continuación). Comunidades con Podostemum spp. Aº: Arroyo.
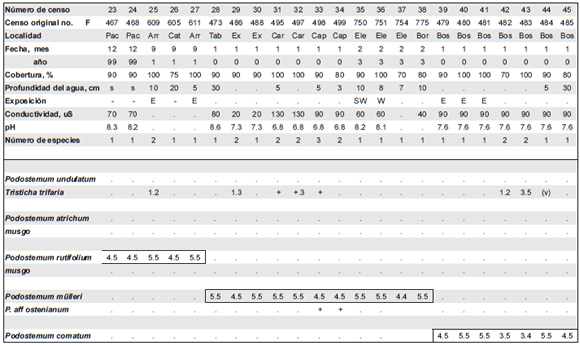
Localidades: Arr: Salto Arrechea (PN Iguazú); Ber: A° Salto; Bos: Salto Bosseti; Cap: Salto de Capioví; Car: A° Caraguatá; Cat: Cataratas; Enc: Salto Encantado; Ex: A° Excelente; Pac: Salto Paca; Per: A° Persiguero; Tar: A° Tarumá; Uru: A° Uruzú; PA: A° Piray Guazú y RP 20; PG: A° Piray Guazú, antes de la confluencia con el A° Alegr'a; Bor: A° Bora (Piray).
3.1- Podostemetum undulati U. Drehwald 1990
Esta comunidad fue encontrada en paredes y pié de las represas (arroyos Salto, Persiguero y Cuñá Manó), en piedras apretadas al borde de los saltos o en rocas planas al pié de saltos (Salto Berrondo) o en cascadas (arroyo Chimiray). Podostemum undulatum forma "matas" grandes con largos tallos verde oscuro y flexibles (30-45 cm) que cuelgan como una cabellera movida por la correntada (como en el Arroyo Salto) o se elevan desde el fondo impulsadas por la corriente ascendente como ocurre en el arroyo Persiguero, en el balneario de Concepción, al pié de la represa (Fig. 6). Siempre crece en arroyos con fuerte turbulencia, cubriendo entre 70 y 100% de la superficie.

Fig. 6. Comunidad de Podostemum undulatum al pié de una represa. Balneario sobre el arroyo Persiguero, Concepción de la Sierra.
En determinados sitios, donde en cortos períodos del año las plantas reciben agua por salpicadura (laterales de saltos), pueden encontrarse hepáticas que acompañan a Podostemum. Aquí es muy clara una estratificación, donde sobresalen las ramas y hojas colgantes de la podostemácea, cubriendo las hepáticas, pegadas a la pared del salto. Podostemum undulatum es la especie predominante y en la gran mayoría de los censos confeccionados, la única. En el 20 % de los censos se encontraron hepáticas, y musgos (Fissidens sp.) que llegan de las comunidades vecinas de la roca húmeda.
Drehwald (1990) describió esta asociación en base a censos realizados únicamente en el arroyo Salto (Salto Berrondo), cuenca del Río Paraná, estableciendo una subasociación típica y una subasociación con Tristicha trifaria. La existencia de la especie en los Arroyos Persiguero, Cuñá Manó (Fontana, 1998) y Chimiray, amplía el área de la comunidad vegetal a la cuenca del Río Uruguay.
En un solo sitio (arroyo Salto) se midieron parámetros químicos. El pH fue neutro a levemente básico (7.5 a 7.9), y la conductividad alcanzó 60 mS.
3.2- Comunidad con Podostemum atrichum
Esta comunidad cubre la roca plana submersa bajo 5 a 10 cm de agua transparente, formando un césped denso y relativamente duro. La planta predominante crece erecta, con el aspecto de muy ramificada por sus hojas rígidas laterales, alcanzando unos 5 (6) cm de altura, con una cobertura variable entre 80 y 100 % (Fig.7). También se observó creciendo sobre piedras de distinto tamaño sumergidas a unos 30 cm de profundidad, con corriente fuerte (arroyo Caraguatay). Los sitios donde se encuentra con mayor frecuencia la comunidad son las rocas planas al pié de represas, siempre sometidos a fuerte correntada, con turbulencia relativamente reducida.

Fig. 7. Perfil de la Comunidad de Podostemum atrichum. Correderas del arroyo Uruzú.
Expuestas todo el día al sol, las plantas tienen un color marrón claro, a excepción de las hojas más nuevas, que son de un verde claro.
Presenta un estrato principal de no más de 6-7 cm de altura con Podostemum atrichum y un estrato inferior, no siempre presente, con Tristicha trifaria, que no supera los 4 cm. En algunas ocasiones forman un estrato intermedio Podostemum rutifolium y P. mulleri, pero con baja cobertura. Podostemum atrichum (tabla 4) es la especie dominante. En el 50 % de los censos (6 censos) también apareció Tristicha trifaria y en algunas ocasiones musgos (Salto Encantado) y rara vez algas verdes.
El pH varió entre levemente ácido (6.3) y levemente básico (7.8). La conductividad mostró una variación mayor, entre 40 mS (arroyo Cuñapirú) y 110 mS (arroyo Uruzú).
Según lo observado hasta el presente, Podostemum atrichum es común en arroyos de la cuenca del Río Paraná, y más raro en la cuenca del Río Uruguay; fue observado en el curso superior de los arroyos del Medio y Acaraguá, afluentes del Uruguay (Zambiasio, 2012).
3.3- Comunidad de Podostemum muelleri
Con aspecto de césped denso, esta comunidad crece submersa, preferentemente sobre la roca plana, en sitios con corriente muy fuerte, aunque nunca se observó en las paredes de los saltos ni al pié de los mismos. Se distingue por el color marrón claro de la especie predominante, caracterizada por hojas filiformes simples inclinadas en dirección de la corriente del agua.
Forma manchas que cubren la roca entre 70 y 100 %, a una profundidad variable entre 5 y 30 cm. En la mayor parte de los sitios observados, la comunidad crecía expuesta al sol casi todo el día. Se pudieron diferenciar dos estratos (Fig. 8), uno que alcanza 5 cm formado por Podostemum muelleri y un segundo estrato más bajo con plantas que tienen 2-3 cm de altura, entre ellas Tristicha trifaria, de baja cobertura. La tabla 4 muestra a P. muelleri como la especie predominante; en el salto Capioví además se observó Podostemum aff ostenianum. Tristicha trifaria apareció en el 70 % de los censos confeccionados.

Fig. 8. Comunidad de Podostemum muelleri. arroyo Excelente, Parque Provincial Urugua-í, Provincia de Misiones. "a": Podostemum mulleri; "b": Tristicha trifaria.
Hasta el presente, la comunidad se observó sólo en arroyos de la cuenca del Río Paraná (Arroyos Excelente, Tabay, Tarumá, Itá Curuzú, Capioví). El pH varió entre 8.6 (arroyo Tabay) y 6.8 (arroyos Itá Curuzú y Capioví) y la conductividad entre 130 mS (balneario sobre el arroyo Itá Curuzú) y 20 mS (arroyo Excelente, en el Parque Provincial Urugua-í, sin influencia humana).
3.4- Comunidad de Podostemum comatum
Sobre la pared de los saltos de las Cataratas del Iguazú se desarrolla una comunidad vegetal con Podostemum comatum que cubre el basalto de la pared vertical.
Allí la comunidad, que es uniespecífica, muestra rizomas que crecen pegados a la roca en posición descendente. Las ramas que nacen de estos rizomas tienen unas pocas hojas verde-oscuro que se extienden en forma más o menos horizontal, fuera de la cortina de agua, sobresaliendo 5 a 8 cm. La comunidad cubre la pared del salto hasta en un 100 %, desarrollándose entre las matas aisladas de Panicum lilloi. Al pié del salto, sobre bloques planos de basalto submersos, junto a Podostemum comatum aparece también Tristicha trifaria. La fuerte turbulencia del agua en estos sitios provoca oleaje, variando la profundidad entre unos pocos cm y 30 cm.
La figura 9 muestra un perfil de la comunidad en la pared del Salto Bosseti, relevado en enero de 2000, durante una bajante considerable del Río Iguazú que registraba 11.90 m en el hidrómetro de Puerto Iguazú.

Fig. 9. Comunidad de Podostemum comatum. Salto Bosseti, Cataratas del Iguazú.
Los valores de pH y de conductividad obtenidos (7.6 y 90 mS) fueron idénticos a los medidos río arriba de la Garganta del Diablo (ver Comunidad de Mourera aspera). Podostemum comatum que da el nombre a la asociación, parece ser la única especie endémica de Argentina (Tur, 1997).
3.5- Comunidad de Podostemum rutifolium
La comunidad se observó creciendo sobre las caras superior y laterales de bloques de basalto en el pié de los saltos, salpicados permanentemente por el agua, ubicados en los costados de las caídas principales. En los momentos de creciente, el agua cae directamente sobre los bloques, pero con menor fuerza que en la parte central del salto.
Forma un césped denso y duro que puede alcanzar hasta 8 cm de altura, constituido por los tallos erectos, coronados por hojas pequeñas y obovadas, poco divididas, que dan a la comunidad un aspecto singular (Fig. 10).
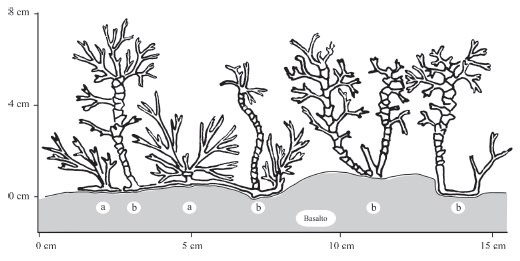
Fig. 10. Comunidad de Podostemum rutifolium. Salto del Moconá. Obsérvese los ejemplares en distinto grado de desarrollo: a, ejemplar juvenil; b, ejemplares adultos.
Predomina Podostemum rutifolium, con 90 a 100% de cobertura, acompañado rara vez por musgos, aún indeterminados (Tabla 4).
Se efectuaron mediciones de los parámetros químicos básicos sólo en el Salto Paca, con los siguientes resultados: pH 8.3 y conductividad 70 a 80 mS. La comunidad fue observada en afluentes del Río Paraná (arroyo Tarumá), en el Río Uruguay (Saltos del Moconá) y algunos de sus afluentes (Salto Paca, Río Yabotí) y en un afluente del Iguazú (Salto Arrechea). Esta especie es la única del género que también crece en ambientes lóticos; fue encontrada en la Laguna Dos Patos en Río Grande Do Sul (Irgang et al., 2003). P. rutifolium también se encuentra presente en el embalse de Yacyretá sobre rocas de los muros de contención (Fontana, 2008).
3.6- Comunidad de Podostemum uruguayense
Esta comunidad es frecuente en los saltos del río Uruguay (por ejemplo Salto del Moconá). También se encuentra esporádicamente en el cauce del A° Yabotí.
Podostemum uruguayense forma un césped duro, muy denso. Crece sobre roca plana y sobre los laterales de piedras sometidas a diferentes velocidades de corriente, desde débil a muy fuerte; sobre roca plana puede alcanzar extensiones superiores a los 4 m2. Parece resistir mejor la exposición al aire que los otros Podostemum, perdiendo las hojas, pero manteniendo vivo los tallos. En esta comunidad crecen también Podostemum rutifolium y Tristicha trifaria.
Fenología
Las podostemáceas que caracterizan las comunidades aquí descriptas florecen durante el período de bajante, cuando las plantas quedan parcialmente expuestas al aire; nunca se observaron plantas en flor en la pared de las caídas (aguas permanentes). En cambio fue común observar plantas submersas con frutos.
En una ocasión (febrero 2003), Podostemum atrichum florecía y fructificaba submerso en agua transparente a 10 cm de profundidad, con el arroyo crecido por precipitaciones ocurridas el día anterior.
En Podostemum spp., la floración y la fructificación parecen realizarse en muy cortos períodos, activadas cuando las plantas quedan fuera del agua. Estas etapas se observaron en el arroyo Salto en noviembre de 1999 durante un período de sequía; las flores, aparentemente anemógamas, rápidamente desaparecieron (1-2 días) dando lugar al desarrollo de los frutos. Como en Podostemum, en Apinagia se observaron botones en enero, cuando el agua bañaba las plantas, pero con el río bajo; las ramas y hojas, permanentemente húmedas por el salpicado, se encontraban por encima de la superficie del agua, cuya profundidad no superaba 5 cm. El ejemplar tipo coleccionado por Chodat & Vischer presentaba botones ya en octubre (Tur, 1997).
Durante las visitas hechas al Parque Nacional Iguazú, nunca observamos a Mourera aspera en flor. En el río Iguazú tendría dos floraciones anuales (información verbal del Sr. Justo Herrera), una a principios del otoño y otra a principios de la primavera, coincidiendo con los períodos de estiaje. Durante la bajante extraordinaria del Río Iguazú en octubre de 2006, varias poblaciones de Mourera quedaron expuestas, produciéndose entonces una profusa floración.
Distribución de las comunidades vegetales
La distribución de estas comunidades vegetales está relacionada directamente con la dispersión de las podostemáceas, las plantas predominantes. La forma de dispersión en las podostemáceas sigue siendo desconocida. Varios autores especularon sobre las posibles formas en que estas plantas dispersan sus semillas. Hoehne (1948) menciona la dispersión de los Podostemum por aves. Las pequeñas semillas se pegarían a las patas y serían así transportadas. Schnell (1969, p.265) opina a favor de una diseminación epizoócora por aves, debido a la presencia de una misma especie en cuencas diferentes, aunque en un trabajo posterior (Schnell in Landolt et al., 1998), lo pone en duda y se inclina por una diseminación anemócora. Varios autores citados por Van der Pijl (1982) mencionan la presencia de mucílago que se forma en torno a la semilla con la humedad y que permitiría adherirse a las patas de aves para ser transportadas; este mucílago también ayudaría al anclaje de la semilla a la roca. Van Royen & Reitz (1971) piensan en una diseminación anemófila, mediante un "hasta floral" que al secarse por efecto del sol se vuelve elástico, y a la menor brisa expele las semillas. Esta suposición es coherente con la diseminación restringida de especies a determinados ríos o cuencas.
Van Steenis (1981, p.340) dice que en primer lugar son plantas hidrócoras; también piensa que aves y el viento pueden contribuir a la dispersión. La dispersión por el agua sería el medio usado por las especies restringidas a una cuenca determinada. La presencia de algunas especies con distribución más amplia, sobre cuencas diferentes estaría explicada por el transporte involuntario de las semillas por aves. En el Parque Provincial Urugua-í, observamos pavas de monte (Penelope sp.) alimentándose en el cauce del arroyo Uruzú, donde Podostemum atrichum forma densas "carpetas" que cubren la roca a muy poca profundidad (5-10 cm). Según el guardaparque M.Chudy es habitual la presencia de estas aves en el sitio.
El relevamiento permitió determinar la existencia de comunidades vegetales limitadas a determinados ríos (Tabla 5). La comunidad de Mourera aspera se observó solamente en el río Iguazú, en el área de cataratas y las correderas río arriba de la Garganta del Diablo. En el alcance actual de nuestros estudios, las comunidades con Podostemum mulleri y de Podostemum atrichum están restringidas a otros afluentes del Río Paraná; la Comunidad de Podostemum muelleri a afluentes del centro-W misionero (Arroyos Capioví, Caraguatá, Tarumá y Tabay); la Comunidad de Podostemum atrichum en el centro y en el N de la provincia (Salto Encantado, Arroyos Uruzú y Tarumá).
Tabla 5. Distribución de comunidades vegetales en los tres grandes ríos y sus afluentes.

En Misiones sólo la comunidad con Podostemum uruguayense es exclusiva de la cuenca del Río Uruguay. Las comunidades de Apinagia yguazuensis, de Podostemum rutifoli y de Marathrum azarensis son las únicas que fueron halladas en afluentes de las dos cuencas, incluyendo la subcuenca del Río Iguazú.
Adaptaciones de las Podostemáceas a las corrientes de agua
Según Ancibor (1990), las especies de Podostemum no tienen espacios aeríferos, poseen escaso xilema, los cloroplastos están ubicados sólo en la epidermis, hay abundancia de sílice, no se observan estomas, tampoco pelos. En la hoja es posible ver un único nervio medio formado sólo por floema. El tejido mecánico es escaso, y está representado por esclerénquima constituido por fibras largas no lignificadas y con paredes gelatinizadas; la presencia de estas fibras dan elasticidad a la planta, cualidad necesaria para resistir la fuerza de tracción a la que están sometidas permanentemente.
Las hojas de las especies argentinas de Podostemum son laciniadas, con numerosas divisiones dicotómicas (en P. undulatum) más o menos aciculares, relativamente rígidas (en P. mulleri y P. uruguayense) pequeñas y obovadas (en P. rutifolium). Los órganos de fijación de estas plantas reófilas son particularmente llamativos por su tamaño y forma en Mourera aspera. Los hápteros o grampones se forman probablemente a partir de ramificaciones de las raíces constituyendo una suerte de "deformación funcional" de raíces laterales (Ancibor, op.cit.). La adhesión a la roca es tal que resulta muy difícil extraer la planta completa. Las hojas de Mourera son grandes, rígidas y muy frágiles; se despliegan en forma paralela al fondo, ofreciendo así un mínimo de resistencia a la corriente del agua (Fig. 11B). Se desconoce la forma de fijación de las semillas de esta especie. Se conoce que en Mourera fluviatilis, los frutos caen sobre la roca desnuda cuando el nivel del agua se encuentra lo más bajo y con la humedad de las primeras lluvias luego del período seco, forman un mucílago fijador (Schnell in Landolt et al., 1998). En Apinagia yguazuensis las adaptaciones consisten en tallos rígidos dorsiventrales apretados contra la roca, de crecimiento paralelo a la misma, apenas elevados o en algunos casos totalmente erectos. Según la velocidad de la corriente, sus hojas son laciniadas, con numerosas divisiones pequeñas en los extremos en sitios con corriente fuerte (Salto Tabay), y más anchas, semejantes a pequeñas hojas con bordes laciniados en los sitios de menor correntada (arroyo Yazá).

Fig. 11. Formas de las hojas en especies de Podostemaceae de Misiones. Hojas aplanadas divididas en Marathrum azarensis (A); hojas latifoliadas anchas (B) en Mourera aspera (obsérvese la posición de las hojas para ofrecer una menor resistencia a la corriente de agua); hojas laciniadas con numerosas divisiones dicotómicas en Podostemum atrichum (C y D) y en Podostemum undulatum (F); hojas aciculares en P. mulleri. (E).
Conservación
Quiroz et al. (1997) determinaron en México que las Podostemaceae son sensibles a la contaminación de las aguas, por lo que su desaparición de un arroyo o un mal crecimiento, podrían ser indicadores de la modificación de las condiciones químicas del agua. En arroyos con aguas contaminadas (vertido de aguas servidas) o con transparencia disminuida (fábrica de ladrillos) se observó el deterioro de estas comunidades vegetales, incluso la desaparición localizadas de las plantas, que aguas arriba y a cierta distancia aguas abajo del foco de contaminación crecen en condiciones normales (arroyo Salto, aguas arriba del salto Berrondo). La posibilidad del uso de estas plantas como indicadores biológicos de contaminación de aguas y de impacto ambiental de represas ya fue propuesto por Meijer (1976).
La construcción de pequeñas represas para la instalación de pequeños generadores de electricidad domiciliarios convirtieron a algunos tramos de pequeños arroyos en aguas lénticas con la consecuente muerte en esos sectores de las comunidades de reófitos. La construcción de la represa de Urugua-í determinó la desaparición del salto del mismo nombre así como la formación de un embalse de varios kilómetros en el valle del arroyo del mismo nombre, en una zona rica en vegetación reófila. Westeiniola apipensis, sólo conocida de su localidad típica (Rápidos de Apipé, provincia de Corrientes), no volvió a observarse después de la construcción de la represa de Yacyretá.
En otros países algunas podostemáceas son objeto de intensa recolección, poniendo en peligro el futuro no sólo de la planta sino de numerosas especies de animales asociadas. Fleck (2004) menciona a Mourera fluviatilis recolectada en Guayana francesa para la producción comercial de cosméticos.
Agradecimientos
La presente publicación es un resultado de los estudios realizados en el marco de los proyectos de investigación sobre la vegetación reófila y sus condiciones ambientales acreditados por la Secretaría de Ciencia y Técnica de la Universidad Nacional del Nordeste. Se agradece a las autoridades del Parque Nacional Iguazú por permitir el relevamiento y las colecciones en el área Cataratas, a los Guardaparques y especialmente a J. Herrera (CIES, Centro de Investigaciones Ecológicas Subtropicales) por su compañía y los datos sobre ubicación de las comunidades. Al Ministerio de Ecología y Recursos Naturales Renovables de la Provincia de Misiones que autorizó la permanencia y la colección en sus reservas. A Ernesto Krauczuck y a los guardaparques por la colaboración en la colección de plantas en distintas épocas del año. A Claudia, esposa y compañera incansable en los viajes.
Bibliografía
1. ANCIBOR, E. 1990. Anatomía de las especies argentinas de Podostemum Mich. (Podostemaceae). Parodiana 6: 31-47. [ Links ]
2. BRAUN-BLANQUET, J. 1979. Fitosociología. Bases para el estudio de las comunidades vegetales. H.Blume, Madrid. [ Links ]
3. CARNEVALI, R. 1994. Fitogeografía de la Provincia de Corrientes. Gob. de la Pcia. de Corrientes y INTA. Edic. del autor, Corrientes. [ Links ]
4. CASTELLANOS, A. 1975. Cuenca potamográfica del Río de la Plata. Geografía de la República Argentina 7. pp. 1-159. [ Links ]
5. CHATELLENAZ, M. L. 2007. Fauna asociada a la vegetación reófila del Parque Provincial Moconá (Misiones, Argentina). FACENA 23: 41-54. [ Links ]
6. CHODAT, R. & W. VISCHER. 1917. La végetation du Paraguay. VI.Podostémacées. Bull. Soc. Bot. Genève ser. 2: 165-196. [ Links ]
7. DIERSCHKE, H. 1994. Pflanzensoziologie, Grunlagen und Methoden. UTB Verlag Edit. Stuttgart. [ Links ]
8. DREHWALD, U. 1990. Pflanzensoziologische Untersuchungen an Fliessgewasser-Gesellschaften in NO-Argentinien. Tuexenia 10: 147-151. [ Links ]
9. ESKUCHE, U. 1986. Bericht über die 17.Internationale Pflanzengeographische Exkursion durch Nordargentinien. In: ESKUCHE, U. & E. LANDOLT (eds.). Contribuciones al conocimiento de flora y vegetación del norte de la Argentina. Veröff. Geobot. Inst. ETH 91: 12-117. [ Links ]
10. FLECK, G. 2004. Contribution à la connaissance des Odonates de Guyane française. Les larves de Macrothemis pumila Karsch, 1889 et de Brechmorhoga praedatrix Calvert, 1909. Notes biologiques et conséquences taxonomiques (Anisoptera: Libellulidae). Ann. Soc. Entomol. Fr.(n.s.) 40: 177-184. [ Links ]
11. FONTANA, J. L. 1998. Análisis sistemático-ecológico de la flora del sur de Misiones (Argentina). Candollea 53: 211-301. [ Links ]
12. FONTANA, J. L. 2001. La vegetación reófila del Nordeste argentino. 1. Las comunidades vegetales con Mourera aspera y Apinagia yguazuensis. [online]. Disponible en: http://www.unne.edu.ar/unnevieja/Web/cyt/cyt/2001/cyt.htm [Acceso: 27 septiembre 2013]. [ Links ]
13. FONTANA, J. L. 2007. Marathrum (Podostemacea), un nuevo género para el Paraguay. Candollea 62: 145-148. [ Links ]
14. FONTANA, J. L. 2008a. Aspectos ecológicos y nuevas citas de Apinagia yguazuensis (Podostemaceae). Bol. Soc. Arg. Bot. 43: 269-272. [ Links ]
15. FONTANA, J. L. 2008b. Alteraciones ambientales y comunidades de Podostemáceas en la represa de Yacyretá, Argentina y Paraguay. [online]. Disponible en: http://www.unne.edu.ar/unnevieja/investigacion/com2008/biologicas.php [Acceso: 27 septiembre 2013]. [ Links ]
16. FONTANA, J. L. & D. E. IRIART (en prensa). La vegetación reófila del Parque Provincial Moconá (Misiones, Argentina). Comunidades vegetales de islotes, correderas y saltos. FACENA. [ Links ]
17. GOBIERNO DE LA PROV. DE MISIONES. 1978. Atlas general de la Provincia de Misiones. Secretaría de Planificación y Control. Dirección General de Estadísticas y Censos. Posadas. [ Links ]
18. GRUBER, M. 1991. Ecología de las fanerógamas de saltos tropicales adaptadas en forma extrema. Natura (Caracas) 91: 54-61. [ Links ]
19. GUILLAUME, M. 2003. New herbivorous pirahnas discovered in french Guiana. Scientific Bulletins 168: 1-2. [ Links ]
20. HAMADA, N. 2000. Simulium (Psaroniocompsa) tergospinosum New Species (Diptera: Simuliidae) in siolii Group from the Southern Part of the State of Amazonas, Brazil. Anais do Istituto O. Cruz 95: 819-828. [ Links ]
21. HOEHNE, F.C. 1948. Plantas aquaticas. Publicaçao da serie "D". Inst. de Bot. Secret. da Agricultura. Sao Paulo. [ Links ]
22. IRGANG, B. E., C. V. SENNA GASTAL JR., C. T. PHILBRICK & A. NOVELLO. 2003. A ocorrência inédita de uma Podostemaceae nas costas de uma laguna (Laguna Dos Patos) no Rio Grande Do Sul, Brasil. Caderno de Pesquisa, Ser. Bio. 15: 7-12. [ Links ]
23. KLEIN, R. M. 1979. Reófitas no Estado de Santa Catarina. Anais XXX Congresso Nacional de Botanica. Soc. Bot. do Brasil. pp.159-169. [ Links ]
24. LANDOLT, E., I. JAGER-ZURN & R. A. A. SCHNELL. 1998. Extreme adaptations in Angiospermous Hydrophytes. Handbuch der Pflanzenanatomie 13: 1-290. [ Links ]
25. MEIJER, W. 1976. A note on Podostemum ceratophyllum Michx. As indicator of clean streams in and around the Appalachian Mountains. Castanea 41: 319-324. [ Links ]
26. MERCANTI, J. A. 2007. 100 Saltos de agua. Provincia de Misiones. Altuna impresores, Buenos Aires. [ Links ]
27. NOVELO, R. A. & C. T. PHILBRICK. 1997. Taxonomy of Mexican Podostemaceae. Aquatic Botany 57: 275-303. [ Links ]
28. OLINUCK, J. A. 1995. El clima de la localidad de Montecarlo. INTA Cerro Azul.Informe técnico N ° 62: 1-46. [ Links ]
29. OLINUCK, J. A. 1998. El clima de la localidad de Cerro Azul. INTA Cerro Azul.Informe técnico N° 68: 1-73. [ Links ]
30. PEDRALLI, G. & S. T. MEYER. 1996. Levantamento da vegetaçao aquatica (macrofitas) e das florestas de galeria na área de Usina Hidrelétrica de Nova Ponte, Minas Gerais. Cadernos do Departamento de Ciencias Biológicas da PUC-Minas 4 : 49-60. [ Links ]
31. PHILBRICK, C. T. & A. NOVELO. 1995. New World Podostemaceae: ecological and evolutionary enigmas. Brittonia 47 : 210-222. [ Links ]
32. PHILBRICK, C. T. & A. NOVELO. 2004. Monograph of Podostemum (Podostemaceae). Systematic Botany Monographs 70: 1-99. [ Links ]
33. PONTIROLI, A. 1955. Podostemáceas argentinas. Bol. Soc.Argent.Bot. 6: 1-20. [ Links ]
34. QUIROZ, A., A. NOVELLO & C. T. PHILBRICK. 1997. Water chemistry and the distribution of Mexican Podostemaceae: a preliminary evaluation. Aquatic Botany 57: 201-212. [ Links ]
35. RUTISHAUSER, R. 1997. Structural and developmental diversity in Podostemaceae (river-weeds). Aquatic Botany 57: 29-70. [ Links ]
36. SCHNELL, R. 1969. Contribution à l'étude des Podostémacées de Guyane. Adansonia, ser. 2, 9: 249-271. [ Links ]
37. SCHNELL, R. 1970. Introduction à la Phytosociologie des pays tropicaux. T1: Les flores, les structures. Collect. Góebiologie-Ecologie-Amenagement. Gauthier-Villars Edit, Paris. [ Links ]
38. SCHNELL, R. 1994. Les stratégies végètales. Essai de morphologie évolutive. Edit. Masson, Paris. [ Links ]
39. SERVICIO METEOROLOGICO NACIONAL. 2010. Estadísticas meteorológicas [online]. Disponible en: http://www.smn.gov.ar/?mod=clima&id=30&provincia=Misiones&ciudad=Iguaz%FA [Acceso: 23 septiembre 2013]. [ Links ]
40. SILVEIRA VILELLA, F., F. GERTUM BECKER & S. M. HARTZ. 2002. Diet of Astyanax species (Teleostei, Characidae) in an Atlantic Forest River in Southern Brazil. Brazilian Archives of Biology and Technology 45: 223-232. [ Links ]
41. SPRADA TAVAREZ, A., O. ODNETZ & A. ENRICONE. 2006. A familia Podostemaceae em rios amazônicos e comunidades de insetos associados. Insula 35: 19-50. [ Links ]
42. TUR, N. M. 1975. Nueva especie de Podostemácea para Argentina. Wettsteiniola apipensis. Bol. Soc. Argent. Bot. 16: 320-324. [ Links ]
43. TUR, N. M. 1997. Taxonomy of Podostemaceae in Argentina. Aquatic Botany 57: 213-241. [ Links ]
44. TUR, N. M. 1999. Podostemaceae. In: SPICHIGER, R. & L. RAMELLA (eds.), Flora del Paraguay. Cons. et Jard. Bot. de la ville de Genéve y Miss.Bot.Gard 29: 1-35. [ Links ]
45. TUR, N. M. 2003. Una nueva especie de Marathrum (Podostemaceae) y nueva cita del género para la Argentina. Hickenia 3: 151-198. [ Links ]
46. VAN DER PIJL, L. 1982. Principles of Dispersal in Higer Plants. 3a. edic. Springer-Verlag, Berlín. [ Links ]
47. VAN ROYEN, P. & P. R. REITZ. 1971. Podostemáceas. Flora Ilustr. Catarinense. Fasc. PODOS pp. 1-36. [ Links ]
48. VAN STEENIS, C. G. 1932. Report of a botanical trip to the Anambas and Natoena Island. Bull. Jard. Bot. Buitenzorg 3: 151-211. [ Links ]
49. VAN STEENIS, C. G. 1981. Rheophytes of the world. Sijthoff y Noordhoff. Holanda. [ Links ]
50. VARELA, M. E. & J. A. BECHARA. 1980. Estructura, distribución y variaciones de la comunidad bentónica en el Alto Paraná y ambientes relacionados. En: CECOAL, Estudios ecológicos en el área Yacyretá. Informe de avance Nº 4: 121-146. [ Links ]
51. WILBERT, J. 1999. The cultural significance of tobacco use in South America. In: SEAMAN, G & J.S.DAY (eds.), Ancient traditions: Shamanism in Central Asia and the Americas, pp. 47-76. Univ. Press of Colorado & Denver Museum of Nat. History, Denver. [ Links ]
52. ZAMBIASIO, V.A., J.L. FONTANA & E. LAFFONT. 2011. Podostemáceas e invertebrados asociados en dos arroyos de la Provincia de Misiones, Argentina. Bol. Soc. Arg. Bot. 46: 213. [ Links ]
53. ZAMBIASIO, V.A., J.L. FONTANA & E. LAFFONT. 2012. Comunidades Vegetales con Podostemáceas y Comunidades asociadas de Invertebrados en dos arroyos de la Provincia de Misiones, Argentina. XX Jornadas de Jovens Pesquisadores da AUGM. UFPR, Curitiba. [ Links ]
54. ZULOAGA F. O., O. MORRONE & M. J. BELGRANO (eds.). 2008. Catálogo de las Plantas Vasculares del Cono Sur (Argentina, sur de Brasil, Chile, Paraguay y Uruguay). Monogr. Syst. Bot. Missouri Bot. Gard. 107: 1-3348. [ Links ]
Recibido el 4 de junio de 2013,
aceptado el 2 de noviembre de 2013.

