










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versão On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.49 no.2 Córdoba jun. 2014
ANATOMÍA Y MORFOLOGÍA VEGETAL
Nueva variante cambial en el género Ephedra (Ephedraceae)
Pablo A. Cabanillas1,2, M. Lucía Borniego3 y Alcides A. Sáenz1
1 Cátedra de Morfología Vegetal, FCNyM, UNLP, Paseo del Bosque s/nro. 1900-La Plata.
2 Becario CIC, Pcia. Buenos Aires; Autor de correspondencia: cabanillaspablo@gmail.com .
3 Instituto de Fisiología Vegetal (INFIVE), FCAyF, UNLP. Becaria CONICET.
Resumen
En esta contribución se presentan los resultados de un estudio sobre el leño caulinar de la liana Ephedra tweediana Fisch. & C.A. Mey. emend. J.H. Hunz. (Ephedraceae), mediante técnicas anatómicas convencionales. Se da a conocer por primera vez para el género la presencia de la variante cambial "tallos lobulados"; y por primera vez para la especie la presencia de leño de reacción (expresado por la médula excéntrica), parénquima radial diagonal y radios agregados.
Palabras clave: Variante cambial; Tallo lobulado; Radios agregados; Parénquima radial diagonal; Leño de reacción; Ephedra tweediana.
Summary
New cambial variant in the genus Ephedra (Ephedraceae). This contribution presents the results of a study on the caulinar wood of the liana Ephedra tweediana Fisch. & C. A. Mey. emend. J. H. Hunz. (Ephedraceae), carried out through conventional anatomic techniques. The presence of the cambial variant called "lobed stem" is disclosed for the genus for the first time. The presence of reaction wood (expressed by eccentric medulla), diagonal and radial parenchyma, and aggregate rays is mentioned for the species for the first time.
Key words: Cambial variant; Lobed stem; Aggregate rays; Diagonal radial parenchima; Reaction wood; Ephedra tweediana.
Introducción
El género Ephedra L. (Gnetidae, Ephedrales) se distribuye en la región Mediterránea (sur de Europa, norte de África), las zonas templado-cálidas de Asia y del oeste de Norteamérica, y el centro-oeste de Sudamérica. Son plantas leñosas, generalmente xeromórficas, erectas (arbustos, rara vez arbolitos), trepadoras, o incluso rastreras, que crecen desde el nivel del mar hasta los 5000 m s.n.m. en los Andes y los Himalayas. El número de especies oscila entre 40 y 55, dado que existe discordancia entre las clasificaciones infragenéricas basadas en datos morfológicos y genéticos (Chase & Reveal, 2009; Christenhusz et al., 2011; Friedman, 1996; Hunziker, 1995; Ickert-Bond & Wojciechowski, 2004).
Ephedra tweediana Fisch. & C.A. Mey. emend. J.H. Hunz. es una trepadora escandente leñosa (liana) que desarrolla dos sistemas de ejes caulinares: rastreros y trepadores. Los tallos rastreros son leñosos, superficiales o subsuperficiales, con nudos enraizantes de los que surgen nuevos vástagos. Los ejes trepadores mantienen una relación estrecha con el soporte y pueden alcanzar los 10 cm de diámetro y varias decenas de metros de longitud (Cabanillas & Hurrell, 2012; Cabanillas et al., 2013). Se distribuye desde el sur de Brasil (Río Grande do Sul), Uruguay (Río Negro, Flores, Florida, San José, Rocha y Maldonado) hasta la Argentina (Salta, Catamarca, Tucumán, Corrientes, Entre Ríos y Buenos Aires); en la provincia de Buenos Aires crece en bosques tanto húmedos como secos, en médanos y bordes de caminos del noreste bonaerense, las sierras de Balcarce y la Isla Martín García (Hunziker, 1995; Hunziker & Novara, 1998; Giehl, 2012; Instituto de Botánica Darwinion, 2012; Souza, 2012).
El presente trabajo tiene por objetivo describir la anatomía xilológica del tallo de E. tweediana.
Materiales y Métodos
Se coleccionaron leños caulinares de 1-10 cm diám., verticales, horizontales e inclinados, provenientes de plantas que crecen en la Isla Martín García. Los ejemplares de referencia correspondientes se depositaron en el Herbario LP, bajo los números Cabanillas 13, 14, 15 (LP) y en la xiloteca XCD-UNLP, bajo los números Cabanillas T9, T24, T25, T26.
Las muestras se analizaron de forma macroscópica (pulido de las superficies convencionales) y microscópica (macerados y secciones). Los macerados se realizaron con solución de Jeffrey y se tiñeron con safranina 1% en alcohol 80. Las secciones, de 50 μm, obtenidas con xilótomo de deslizamiento, fueron teñidas por doble coloración con safranina 1% en alcohol 80 y Fast Green 1% en alcohol 100.
La combinación de rasgos anatómicos observados no es abarcable con uno solo de los manuales de la IAWA (International Asociation of Wood Anatomists), por lo cual las descripción y mediciones fueron realizadas combinando los criterios para "Gimnospermas" (IAWA, 2004), Angiospermas (IAWA, 1989) y los aportes específicos de Carlquist (1989, 1992, 1996). El tipo de variante cambial fue determinado siguiendo las clasificaciones de Metcalfe (1983) y Carlquist (1988), dado que la IAWA no ofrece criterios aplicables.
Resultados
Los tallos hasta de 0,9 cm de diámetro presentan el patrón típico de las plantas leñosas, mientras los de mayor grosor presentan la variante cambial tallos lobulados (Fig. 1B, E).

Fig. 1. A: Esquema general en CT (solo se ilustran los radios multiseriados). B: Esquema detalle de la relación entre los diferentes anillos de crecimiento y radios (Dibujos P. A. Cabanillas). C: Pulido de la cara transversal de tallo vertical lobulado y con médula central. D: Pulido de la cara transversal de tallo inclinado con médula excéntrica (la flecha indica la vertical). E: Aspecto de los tallos trepadores verdes y leñosos (Foto: Daniel Bazzano). F: CT de la zona central: médula, xilema primario y anillos de crecimiento internos. G: Detalle de anillo interno, se observan los radios 1-3 seriados (Ru), los poros y el tejido de fondo formado por traqueidas. Referencias: Ae Anillos de crecimiento externos; Ai Anillos de crecimiento internos; Am Anillos de crecimiento intermedios; Fs Floema secundario; Me Médula; Pe Peridermis; Ra Radios agregados; Rm Radios multiseriados; Ru Radios 1-3 seriados; Xp Xilema primario.
Anillos de crecimiento demarcados por la reducción del diámetro radial de las traqueidas y, en algunos casos, por la ausencia de poros en el leño tardío (Fig. 1C, 2B).
Se observan tres tipos de anillos según su posición en el eje: anillos internos continuos circulares, anillos intermedios continuos sinuosos, y anillos externos quebrados sinuosos (Fig. 1B). Porosidad difusa. Poros exclusivamente solitarios, de contorno subanguloso, sin disposición especial (Fig. 1C, D).
Elementos de vaso de 574 μm long. y 34,18 (15,2- 37,8) μm de diámetro tangencial con paredes de 2,7 (1,89-4,05) μm; 251 vasos por mm2 en promedio. Placa de perforación terminal inclinada, foraminada, generalmente 10 perforaciones por placa organizadas en 2 series (a veces 3) alternas a subopuestas. Punteaduras intervasculares areoladas uniseriadas de contorno circular con torus.
Traqueidas cortas (560 μm long. en promedio), de 17,9 (13,5-24,3) μm de diámetro con paredes de 4,96 (1,89-7,29) μm. Punteaduras iguales a las de los vasos. Fibrotraqueidas (sensu Carlquist, 1989) cortas (550 μm long. en promedio), nucleadas, de distribución difusa. Punteaduras entre fibrotraqueidas simples uniseriadas. Punteaduras fibrotraqueida-traqueida, o vaso, semiareoladas uniseriadas.
Parénquima axial (sensu Carlquist, 1989) ausente. Radios homocelulares heterogéneos de tres tipos: 1-3 seriados, multiseriados y agregados.
Radios 1-3 seriados formados principalmente por células erectas; 571,4 (428,55-1400) μm alt. y 28,57 (19,9-30,2) μm lat. Presentes en los anillos internos e intermedios (Fig. 1D, 2A).
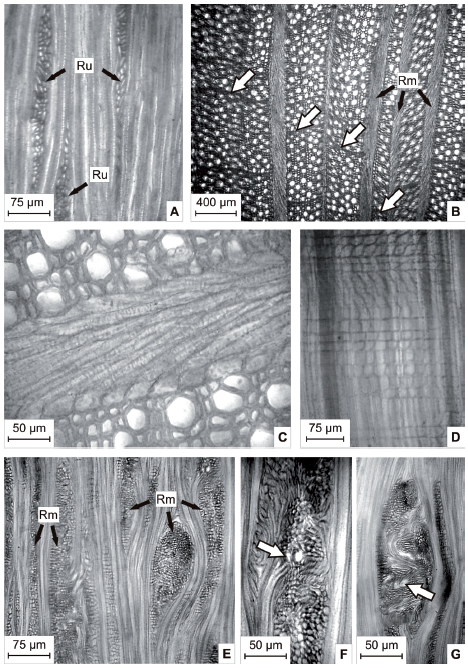
Fig. 2. A: CTg de anillo interno, se observan los radios 1-3 seriados (Ru). B: CT de anillos externos, se observa el desfase de los anillos de crecimiento (flechas blancas) y las diferencias en la orientación de los elementos axiales a ambos lados de los radios multiseriados (Rm). C: CT anillo externo donde se observa el parénquima diagonal de un radio multiseriado. D: CRd de un radio multiseriado donde se observan las hileras de células erectas, decumbentes, cuadradas y diagonales. E: CTg de anillo intermedio con radios multiseriados (Rm). F, G: CTg de radios agregados, se observan vasos y traqueidas de orientación radial (flechas blancas).
Radios multiseriados de 4 a 25 células de ancho, formados por hileras alternas de células erectas, cuadradas, procumbentes y diagonales. Radios de 3,43 mm (hasta más de 10 mm) alt., 0,11428 (0,22856) mm lat. Presentes en los anillos intermedios y externos (Fig. 2B, C, D, E).
Radios agregados de 571,4 μm alt. y 407,12 μm lat. (hasta de 514,26 μm). Presentes en los anillos externos, en especial en los surcos del tallo (Fig. 2F, G).
En los anillos intermedios y externos, los radios multiseriados delimitan sectores de elementos axiales que presentan diferencias leves de orientación unos respecto de los otros (Fig. 2B).
En los anillos externos, los vasos y traqueidas que quedan incluidos en los radios agregados presentan, en muchos casos, orientación radial (Fig. 2F, G). Los tallos horizontales e inclinados presentan la médula de posición excéntrica (Fig. 1 F) y espacios intercelulares dispersos por el leño, sin un patrón definido respecto a la orientación del tallo.
Discusión
El patrón de crecimiento secundario en grosor más común entre las plantas leñosas implica la actividad de un único cambium, cilíndrico y bifacial, que genera xilema centrípetamente y floema centrífugamente. Cualquier alteración de este patrón se denomina variante cambial, crecimiento secundario anómalo o bien estructura anómala. Estos patrones son constantes a nivel específico, es decir, "normales" para la especie, por lo cual se ha recomendado restringir el término anómalo a estructuras teratológicas (Metcalfe, 1983; Carlquist, 1988).
Por lo general, las variantes cambiales son evidentes en las primeras etapas del crecimiento secundario; sin embargo, en algunas especies sólo son evidentes en tallos añosos. Los ejemplares de herbario de leñosas, en general, contienen los órganos reproductivos, hojas y sólo los ejes jóvenes, muy rara vez incluyen leños mayores a 1 cm de diámetro (Lev-Yadun & Aloni, 1993; Tamaio & Somner, 2010). Carlquist (1989) estudió el leño de un único ejemplar de E. tweediana procedente de la Isla Martín García, leg. Pastore (SI 27998), de 9 mm de diámetro en el cual la actividad cambial concordaba con el patrón típico de las plantas leñosas. Dicha descripción sólo es válida, y de manera parcial, para las primeras etapas del crecimiento secundario, ya que los rasgos anatómicos cambian con la edad de la planta.
Según Carlquist (1989, 1992, 1996) Ephedra presenta un único cambium que puede tener una incipiente diferencia en las tasas de producción de xilema entre las áreas separadas por los radios multiseriados, mientras el parénquima radial diagonal acomodaría las diferencias entre dichas áreas.
Lev-Yadun & Aloni (1993) dieron a conocer la primer variante cambial para el género. En los tallos añosos de E. campylopoda G. A. Mey., especie de las montañas y planicies costeras de Israel, los radios son muy anchos (de varias decenas de células), por lo general agregados, y pueden presentar una zona central parenquimática con aspecto de médula. Los sistemas radial y axial cambian sus orientaciones típicas, se entremezclan, adquieren trayectos sinuosos, incluso segmentos del sistema axial aparecen con orientación radial. Así, el patrón tridimensional producido es complejo y no responde a la actividad típica del cambium.
El leño de reacción se forma en ejes que se alejan de la posición vertical. El leño de compresión gimnospérmico se caracteriza por una mayor producción de xilema en la cara inferior, la presencia de traqueidas más cortas que el leño normal, de contorno redondeado, con espacios intercelulares. Las fibras gelatinosas son comunes en el leño de tensión angiospérmico y, aunque están ausentes en el xilema de las "Gimnospermas", son abundantes en la peridermis y el floema secundario de Gnetum gnemon L. (Gnetales) y de Ephedra. En G. gnemon se ubican en la cara superior de los ejes inclinados y funcionan como estructura de tensión; en Ephedra, no se ha detallado su distribución (Carlquist, 1989; Lev-Yadun, 1999; Tomlinson, 2003).
Conclusiones
Algunos aspectos del xilema secundario de E. tweediana han sido estudiados previamente (Carlquist, 1989); sin embargo, el estudio de tallos de diámetro mayor nos permite, en esta contribución, dar a conocer por primera vez para la especie la presencia de radios agregados (registrados previamente en E. campylopoda; Lev-Yadun & Aloni, 1993); parénquima radial diagonal (presente en la mayoría de las especies; Carlquist, 1989, 1992); variante cambial y leño de compresión.
E. tweediana presenta un cambium único que genera xilema centrípetamente y floema centrífugamente. En los primeros años de crecimiento secundario, las tasas de producción de xilema son iguales a lo largo de todo el cambium (anillos internos). Luego, dichas tasas aumentan en ciertas zonas respecto de las vecinas. Estas diferencias se expresan en anillos continuos de contorno sinuoso (anillos intermedios). Posteriormente, se generan desfases entre los segmentos axiales delimitados por los radios anchos, y en la orientación de los elementos axiales entre dichos segmentos. El cambium se vuelve acanalado, los anillos de crecimiento adquieren un contorno quebrado, se desarrollan los radios agregados y el parénquima diagonal en los radios multiseriados (anillos externos), por lo que el tallo adquiere un contorno lobulado.
En E. tweediana los espacios intercelulares, característicos del leño de reacción gimnospérmico, se hallan dispersos por el xilema sin un patrón específico respecto a la orientación del eje. Al igual que en E. campylopoda (Lev-Yadun, 1999), el leño de reacción se expresa por una mayor generación de xilema en la cara inferior, es decir como leño de compresión, a pesar de que no se observaron diferencias anatómicas entre la cara inferior y superior. De lo expuesto previamente podemos concluir que el leño de E. tweediana presenta una combinación de rasgos anatómicos particulares que permiten distinguirlo de las otras especies del género: Tallos lobulados, leño de compresión, espacios intercelulares dispersos, tres tipos de radios (uniseriados, multiseriados y agregados) y parénquima radial diagonal.
Agradecimientos
Los autores desean expresar su agradecimiento al Dr. Julio Hurrell por la cuidadosa lectura del manuscrito; a Daniel Bazzano por las fotografías y las observaciones en el campo; al Prof. Emilio Ulibarri, por su invalorable ayuda en los trabajos de campo; y a la Lic. Stella Maris Rivera por la lectura crítica del manuscrito.
Bibliografía
1. CABANILLAS , P. A. & J. A. HURRELL . 2012. Plantas trepadoras: tipo biológico y clasificación. Revista Ci. Morfol. (La Plata) 14: 1-15. [ Links ]
2. CABANILLAS , P. A., M. L. BORNIEGO, A. A. SAENZ & J. A. HURRELL. 2013. una nueva estrategia de ascenso en plantas trepadoras: Entramado caulinar en Ephedra tweediana (Ephedraceae). Bol. Soc. Arg. Bot. (Aceptado). [ Links ]
3. CARLQUIST , S. 1988. Comparative wood anatomy: systematic, ecological and evolutionary aspects of dicotyledon wood. 448 pp. Springer-Verlag. Berlín. [ Links ]
4. CARLQUIST , S. 1989. Wood and bark anatomy of the New World species of Ephedra. Aliso 12: 441-483. [ Links ]
5. CARLQUIST , S. 1992. Wood, bark, and pith anatomy of Old World species of Ephedra and summary for the genus. Aliso 13: 255-295. [ Links ]
6. CARLQUIST , S. 1996. Wood, bark, and stem anatomy of Gnetales: a summary. Int. J. Plant Sci. 157 (Suppl.): S58-S76. [ Links ]
7. CHASE, M. W. & J. L. REVEAL. 2009. A phylogenetic classification of the land plants to accompany APG III. Bot. J. Linn. Soc. 161: 122-127. [ Links ]
8. CHRISTENHUSZ, M. J. M., J. L. REVEAL, A. FARJON, M. F. GARDNER, R. R. MILL & M. W. CHASE. 2011. A new classification and linear sequence of extant gymnosperms. Phytotaxa 19: 55-70. [ Links ]
9. FRIEDMAN , W. E. (ed.). 1996. Biology and Evolution of the Gnetales. 125 pp. Int. J. Plant Sci. 157 (Suppl. [ Links ]).
10. GIEHL , E. L. (coord.). 2012. Ephedra. En Flora Digital do Rio Grande do Sul e Santa Catarina. Disponible: <http//ufrga.br./floradigital> [Consulta: 15-VII- 2013]. [ Links ]
11. HUNZIKER , J. H. 1995. Ephedraceae. En Hunziker, A.T. (ed.), Flora Fanerógama Argentina 4: 15-23. [ Links ]
12. HUNZIKER, J. H. & L. J. NOVARA. 1998. Ephedraceae. En Novara, L. J. (ed.), Flora del Valle de Lerma. Aportes Bot. Salta, Ser. Flora 5: 1-14. [ Links ]
13. IAWA 1989. List of microscopic features for hardwood identification. IAWA Bull. n.s. 10: 219-332. [ Links ]
14. IAWA. 2004. List of microscopic features for softwood identification. IAWA J. 25: 1-70. [ Links ]
15. INSTITUTO DE BOTÁNICA DARWINION. 2012. Flora del cono sur. Disponible: <http://www2.darwin.edu.ar/> [Consulta: 15-VII-2012]. [ Links ]
16. ICKERT-BOND, S. M. & M. F. WOJCIECHOWSKI. 2004. Phylogenetic Relationships in Ephedra (Gnetales): Evidence from Nuclear and Chloroplast DNA Sequence Data. Syst. Bot. 29: 834-849. [ Links ]
17. LEV-YADÚN, S. & R. ALONI. 1993. Variant secondary growth in old stems of Ephedra campylopoda G. A. Mey. Bot. J. Linn. Soc. 112: 51-58. [ Links ]
18. LEV -YADÚN , S. 1999. Eccentric deposition of secondary xylem in stems of the climber Ephedra campylopoda (Gnetales). I.A.W.A. Journal 20: 165-170. [ Links ]
19. METCALFE, C. R. 1983. Anomalous structure. En Metcalfe, C.R. & L. Chalk (eds.), Anatomy of the Dicotyledons, ed. 2. 330 Pp. Clarendon Press, Oxford. [ Links ]
20. SOUZA, V. C. 2012. Gimnospermas. En Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. Disponible: <http://floradobrasil.jbrj.gov.br/2012/FB035363> [Consulta: 15-VII-2012]. [ Links ]
21. TAMAIO, N. & G. V. SOMNER . 2010. Development of corded vascular cylinder in Thinouia restingae Ferruci & Somner (Sapindaceae: Paullinieae). J. Torrey Bot. Soc. 137: 319-326. [ Links ]
22. TOMLINSON, P. B. 2003. Development of gelatinous (reaction) fibers in stems of Gnetum gnemon (Gnetales). Am. J. Bot. 90: 965-972. [ Links ]
Recibido el 6 de agosto de 2013,
aceptado el 27 de febrero de 2014.

