










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.50 no.1 Córdoba mar. 2015
PALEOBOTÁNICA Y PALINOLOGÍA
Morphology and ultrastructure of Schizaea fistulosa (Schizaeaceae) spores from Chile
Juan P. Ramos Giacosa1, 3, Marta A. Morbelli1, 3 and Gabriela E. Giudice2
1 Cátedra de Palinología, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s/n°, B1900FWA, La Plata, Argentina. E-mail: jpramosgiacosa@hotmail.com
2 Cátedra de Morfología Vegetal
3 CONICET
Summary
The spores of Schizaea fistulosa from Chile were studied using light microscopy (LM), scanning (SEM) and transmission electron microscopy (TEM). The spores are monolete and elliptic in polar view. The major equatorial diameter is 71-85 μm and the polar diameter is 54-61 μm. The laesurae are 50-60 μm long and, in some cases, bifurcated. The sporoderm ultrastructure is first mentioned and described here. The exospore is two-layered in section, verrucate-tuberculate with single or fused elements forming short ridges. The perispore is single-layered, 10-30 nm thick and it is only visible under transmission electron microscopy. On the spore surface, numerous, single or fused, spheroids and nanospheroids of different sizes were observed attached to the perispore surface. The results are discussed and compared with previous studies in Schizaea.
Key words: Schizaeaceae; Schizaea fistulosa; Spores; Morphology; Ultrastructure.
Resumen
Morfología y ultraestructura de las esporas de Schizaea fistulosa (Schizaeaceae) de Chile. Se estudiaron las esporas de Schizaea fistulosa de Chile con microscopía óptica (MO), electrónica de barrido (MEB) y transmisión (MET). Las esporas son monoletes y elípticas en vista polar. El diámetro ecuatorial mayor es de 71-85 μm y el diámetro polar de 54-61 μm. Las lesuras son de 50-60 μm de longitud y en algunos casos están bifurcadas. La ultraestructura de la esporodermis se describe por primera vez. El exosporio en sección posee dos capas y es verrucoso-tuberculado con elementos aislados o fusionados formando lomos cortos. El perisporio posee una sola capa, de 10-30 nm de espesor y solamente es visible con microscopía electrónica de transmisión. Sobre la superficie de la espora, se observan numerosos esferoides y nanoesferoides de diferentes tamaños, adheridos a la superficie. Se discuten los resultados y se los compara con estudios previos en Schizaea.
Palabras clave: Schizaeaceae; Schizaea fistulosa; Esporas; Morfología; Ultraestructura.
Introduction
The spores of Schizaea fistulosa Labill were analyzed as part of the study of the Schizaeales spores from the southern South America (Ramos Giacosa et al., 2012; Ramos Giacosa et al., 2013; Ramos Giacosa 2014). The genus Schizaea Sm. is part of the Schizaeaceae. Previously, this family included the genera Anemia Sw., Mohria Sw., Lygodium Sw., Schizaea and Actinostachys Wall. However, according to recent phylogenetic studies (Smith et al., 2006), the Schizaeaceae is a monophyletic family that comprises only two genera: Schizaea and Actinostachys. Schizaea includes about 30 species distributed in tropical and south warm-temperate areas of the New and Old World (Wikstrom et al., 2002). It is distinct by its erect, apically annular sporangia on flag-like projections of the blade and grass-like to flabellate lamina (Mickel & Smith, 2004). Schizaea fistulosa was reported as growing in Argentina (Malvinas Islands), Chile, New Zealand, Auckland Islands, Australia, Madagascar, New Caledonia and Fiji (Moore, 1968; Large & Braggins, 1991; de la Sota et al., 1998; Zuloaga et al., 2008). Nevertheless, this species has not been seen in Malvinas Islands since it was reported by Gaudichaud-Beaupré in 1825 (as Schizaea australis) (Gaudichaud-Beaupré, 1825). Thus, this species could be considered extinct for the Argentinean flora (de la Sota et al., 1998).
Concerning palynological contributions, Heusser (1971) studied with light microscope (LM) the spores of Schizaea fistulosa from Chile and described the spore ornamentation as gemmate with gemmae of irregular size. When Morbelli (1980) analyzed material from Chile with LM, she described the ornamentation as scabrate with gemmae, verrucae and granules. This author asks some questions about the presence or absence of perispore and wonders if the granules are part of the exospore or the perispore.
Van Konijnenburg-van Cittert (1991), in a comparative study of fossil and extant spores of Schizaeaceae, illustrated with scanning electron microscope (SEM) the spores of Schizaea elegans (Vahl) Sw. and S. pectinata (L.) Sw. Tryon & Lugardon (1991) analyzed with SEM and transmission electron microscope (TEM) the spores of a few species of Schizaea. The authors illustrated with SEM, material of Schizaea fistulosa from Australia and described the spores as spheroidal, with a proximal face papillate and large spherules. Large & Braggins (1991) studied with LM and SEM the spores of Schizaea species from New Zealand. Schizaea fistulosa was one of the species studied by these authors and they described the spore ornamentation under SEM as granulate with large, rounded globular structures, irregularly distributed and frequently more common and larger in size around the laesura. They also mentioned that apparently the perispore was absent.
Later, Parkinson (1994, 1995 a, b) studied the spore wall development in Schizaea pectinata from South Africa. The author described the species sporoderm as having a two-layered exospore and a two-layered perispore. No studies on the spores of Schizaea fistulosa with TEM were carried out. The sporoderm ultrastructure of this species is still unknown. The aim of this work is to analyze the spore morphology and wall ultrastructure of Schizaea fistulosa from Chile in order to get a better knowledge of the spores within the genus.
Material and Methods
Spores were obtained from herbarium specimens from the following institutions (abbreviations according to Holmgren et al., 1990): División Plantas Vasculares, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata (LP) and Instituto de Botánica Darwinion (SI). The spores were studied using light (LM), scanning (SEM) and transmission (TEM) electron microscopy. For LM, the spores were treated with hot 3% sodium carbonate for two minutes in order to preserve the perispore (Morbelli, 1980) and acetolyzed according to the method of Erdtman (1960). For SEM, the material was treated with hot 3% sodium carbonate, washed, dehydrated, suspended in 96% ethanol and then transferred to acetate plates. After drying, they were coated with gold.
For studies with TEM, dry material from herbarium specimens was hydrated following the technique by Rowley & Nilsson (1972), by use phosphate buffer and alcian blue (AB). Then, the material was fixed with Glutaraldehyde + 1 % Alcian Blue in phosphate buffer for 12 h and postfixed with 1% OsO4 in water plus 1% Alcian Blue. The spores were dehydrated in an acetone series and then embedded in Spurr soft mixture. Three μm thick sections were stained with toluidine blue and observed with LM. Ultrathin sections were stained with 1% uranyl acetate for 15 min followed by lead citrate for three min. The observations were made with an Olympus BH2, a JEOL JSMT-100 scanning electron microscope, and a Zeiss T-109 transmission electron microscope.
Studied material: CHILE: Prov. Chiloé, III- 1924, Werdermann 298 (SI). Prov. Magallanes, Dpto. Ultima Esperanza, Isla Virtudes, 2-XII-1977, Roig y Méndez s/n° TBPA-FIT 2613 (LP).
Results
Morphology
The spores are monolete and elliptic in polar view. In equatorial view, the proximal face is plane to convex and the distal face is convex. The major equatorial diameter is 71-85 μm and the polar diameter is 54- 61 μm. The laesurae are 50-60 μm long and in some cases, they are bifurcated near their apices. Analyzed with SEM, the exospore is verrucatetuberculate. The verrucae and tubercles observed are single or partially fused forming short ridges, which are distributed in several directions (Fig. 1E-F). Among the ornamental elements some perforations are also visible (Fig. 1E).
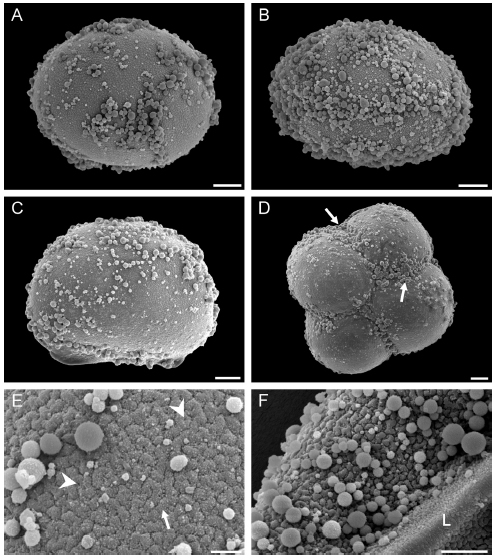
Fig. 1. Spores of Schizaea fistulosa with SEM. A: Distal view of an elliptic spore. B: The high density of spheroids, makes the visualization of the spore surface difficult in some areas. C: Equatorial view of a planeconvex spore. D: Tetrad of spores. The globules are especially abundant in the area of contact between adjacent spores (arrows). E, F: Detail of the spore surface. The exospore is verrucate-tuberculate. The verrucae and tubercles are single or partially fused forming short ridges (arrow). Some perforations are also visible (arrowheads). Numerous single spheroids or spheroids fused to others are observed. The laesurae (L) is evident. Scale bars: A-D: 10 μm. E: 2 μm. F: 5 μm.
The perispore is not evident under SEM. On the spore surface, numerous single or fused spheroids are evident. In some spores the high density of spheroids fused to the spore make it surface visualization difficult (Fig. 1 A-D).
Ultrastructure
Analyzed with TEM, the exospore is 1.2-3 μm thick and two-layered: having an inner layer of 100- 150 nm thick and an outer layer of 1.2-3μm thick. The outer exospore layer forms the elements of the ornamentation which is composed of irregular, partially fused verrucae and tubercles (Fig. 2 A, B). Radial channels with dark content are observed in the outer layer of the exospore (Fig. 2 E, F). The perispore is 10-30 nm thick and more contrasted than the exospore. This wall is adhered to the exospore and covers the outer surface. Nanospheroids of 55-120 nm of diameter are attached to the perispore surface (Fig. 2 E, F).

Fig. 2. Spores of Schizaea fistulosa with TEM. A: Section through the sporoderm. The exospore is twolayered. The inner exospore layer (arrow) is thin and frequently difficult to differentiate. The outer exospore layer (Eo) forms the elements of the ornamentation. Numerous globules are evident on the spore surface. On the globules surfaces, nanospheroids (arrowheads) are seen. B: Section through a laesurae. The perispore is thin and continuous in its surface. C: The outer exospore forms the verrucae and tubercles of the spore ornamentation. The perispore (arrow) is very thin, it is more contrasted than the exospore and it covers the outer surface. On the spore surface, the globules are fused (double arrow). D: Few globules are fused to the exospore surface (arrowhead). Channels with dark content are seen in the exospore (arrows) E, F: Detail of the sporoderm. Nanospheroids (arrowheads) are observed on the perispore surface. Channels are also seen (arrows). Scale bars: A, B: 1 μm. C, D: 0.4 μm. E, F: 200 nm.
On the spore surface, there are spheroids of 0.4- 3.5 μm in diameter. The spheroids are single, fused to one another or to the perispore. Exceptionally, they are fused to the exospore. They have a structure similar to the exospore but with parallel lines which traverse the spheroids all throughout. They are covered by a perispore layer with abundant nanospheroids (Fig. 2 A-D).
Discussion and Conclusions
In the present work, we defined the spore ornamentation of Schizaea fistulosa as verrucatetuberculate. These two elements are single or partially fused and form short ridges distributed in several directions. These results are similar to the descriptions of spores from Chile mentioned by Heusser (1971) and Morbelli (1980) and they are also in accordance with the observations made by Large & Braggins (1991) of spores from New Zealand. In some spores, the laesurae is bifurcated near their apices. This characteristic was also mentioned by Heusser (1971). Tryon & Lugardon (1991) observed with SEM a spore of Schizaea fistulosa from Australia and mentioned the presence of spherules. The authors defined the spherules as small bodies with a concentric arrangement of the different perisporal materials.
However, based on the information provided here, the ultrastructural analysis of Schizaea fistulosa performed with TEM has demonstrated that the abundant spheroids on the spore surface have in their centre a similar structure to that of the exospore but with parallel lines that traverse the spheroids all throughout. They are covered by a perispore layer which is more contrasted than the exospore. These spheroids were resistant to the acetolysis method and, according to the characteristics mentioned above, they would be "globules" as defined by Lugardon (1981). These globules may be homologous to the Ubisch bodies of the Spermatophytes. Large & Braggins (1991) mentioned that, in the analyzed specimens, the globules were frequently more common and larger in size around the laesura. These characteristics were not observed in the material analyzed in the present study. However, when tetrads were observed, the globules were especially abundant in the contact area with the adjacent spores.
The sporoderm ultrastructure for the species is defined here for the first time. The ultrastructural analysis has demonstrated the presence of a single-layered perispore in this species that could only be visible under TEM. The two-layered exospore is the thickest wall of the sporoderm and the one that bears the spore ornamentation. The thin perispore covers the outer surface of the exospore. On the other hand, spore development studies in other species of this genus as Schizaea pectinata from South Africa (Parkinson, 1994, 1995a), described a two-layered exospore and also a two-layered perispore. In the present work, abundant nanospheroids attached to the surfaces of the perispore and of the globules were observed. The presence of nanospheroids on the perispore and the globules could mean that they transport material from the tapetum to complete the development of the perispore.
Although the type of tapetum is unknown for the species studied here, previous work on Schizaea pectinata (Parkinson, 1995 b) reported a combined tapetum consisting of a cellular, parietal and a plasmodial component. This author also suggested that some spherical bodies play a role in the development of the perispore. Further studies on other species of the genus, which analyze the development of the spore wall, could help to understand if the palynological data given in the present work could be relevant for systematic purposes, and also if these characteristics are representative of the family Schizaeaceae.
Acknowledgments
The authors thank Lisandro Anton, from the TEM Unit, Instituto de Biología Celular, Facultad de Medicina, Universidad de Buenos Aires. This work was supported by grants from Universidad Nacional de La Plata (project 584) and Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) for project PICT 0661.
Bibliography
1. DE LA SOTA, E. R., M. M. PONCE, M. A. MORBELLI & L. A. CASSÁ DE PAZOS. 1998. Pteridophyta. En: M. N. CORREA (dir.). Flora Patagónica. Colecc. Ci. Inst. Nac. Tecnol. Agropecu. 8: 282-369. [ Links ]
2. ERDTMAN, G. 1960. The acetolysis method. A revised description. Svenk Botanisk Tidskrift 54: 561-564. [ Links ]
3. GAUDICHAUD-BEAUPRÉ, C. 1825. Rapport sur la Flore des îles Malouines. Ann. Sci. Nat. Paris 5: 89-110. [ Links ]
4. HEUSSER, C. J. 1971. Pollen and spores of Chile. Modern types of the Pteridophyta, Gymnospermae and Angiospermae. The University of Arizona Press. 167 p. [ Links ]
5. HOLMGREN, P. K., N. H. HOLMGREN & L. C. BARNETT. 1990. Index Herbariorum. Part I: The herbaria of the world. New York Botanical Garden, New York. 120 p. [ Links ]
6. LARGE, M. F. & J. E. BRAGGINS. 1991. Spore atlas of New Zealand ferns and fern allies. N. Z. J. Bot. (suppl. 29): 1-168. [ Links ]
7. LUGARDON, B. 1981. Les globules des Filicinées homologues des corps d'Ubisch des Spermatophytes. Pollen et Spores 23: 93-124. [ Links ]
8. MICKEL, J. T. & A. R. SMITH. 2004. The Pteridophytes of Mexico. Mem. New York Bot. Gard. 88: 48-60. [ Links ]
9. MOORE, D. M. 1968. The vascular flora of the Falkland Islands. British Antartic Survey, Scientific Report 60: 49-51. [ Links ]
10. MORBELLI, M. A. 1980. Morfología de las esporas de las Pteridophyta presentes en la Región Fuego- Patagónica. República Argentina. Opera Lilloana 28: 1-138. [ Links ]
11. PARKINSON, B. M. 1994. Morphological and ultrastructural variations in Schizaea pectinata (Schizaeaceae: Pteridophyta). Bothalia 24: 203-210. [ Links ]
12. PARKINSON, B. M. 1995 a. Spore wall development of Schizaea pectinata (Schizaeaceae: Pteridophyta). Grana 34: 217-228. [ Links ]
13. PARKINSON, B. M. 1995 b. The tapetum in Schizaea pectinata (Schizaeaceae) and a comparison with the tapetum in Psilotum nudum (Psilotaceae). Plant Syst. Evol. 196: 161-172. [ Links ]
14. RAMOS GIACOSA, J. P., M. A. MORBELLI & G. E. GIUDICE. 2012. Spore morphology and wall ultrastructure of Anemia Swartz species (Anemiaceae) from Argentina. Rev. Palaeobot. Palynol. 174: 27-38. [ Links ]
15. RAMOS GIACOSA, J. P., M. A. MORBELLI & G. E. GIUDICE. 2013. Comparative palynological analysis of Lygodium venustum Sw. and L. volubile Sw. (Lygodiaceae). An. Acad. Bras. Cienc. 85: 699-707. [ Links ]
16. RAMOS GIACOSA, J. P. 2014. Abnormal spore morphology and wall ultrastructure in Anemia tomentosa var. anthriscifolia and A. tomentosa var. tomentosa (Anemiaceae). Plant Syst. Evol. 300: 1571- 1578. [ Links ]
17. ROWLEY, J. R. & S. NILSSON. 1972. Structural stabilization for electron microscopy of pollen from herbarium specimens. Grana 12: 23-30. [ Links ]
18. SMITH A. R., K. M. PRYER, E. SCHUETTPELZ, P. KORALL, H. SCHNEIDER & P. WOLF. 2006. A classification for extant ferns. Taxon 55: 705-731. [ Links ]
19. TRYON A. F. & B. LUGARDON. 1991. Spores of Pteridophyta. Springer- Verlag, New York. 648 pp. [ Links ]
20. VAN KONIJNENBURG-VAN CITTERT J. H. A. 1991. Diversification of spores in fossil and extant Schizaeaceae. In: BLACKMORE S. & S. H. BARNES (eds.). Pollen and Spores, Patterns of Diversification. Publ. Syst. Assoc. spec. vol. 44: 103-118. [ Links ]
21. WIKSTRÖM N., P. KENRICK & J. C. VOGEL. 2002. Schizaeaceae: a phylogenetic approach. Rev. Palaeobot. Palynol. 119: 35-50. [ Links ]
22. ZULOAGA F. O., O. MORRONE & M. BELGRANO (eds.) 2008. Catálogo de las plantas vasculares del Cono Sur.Vol.1, Pteridophyta, Gymnospermae y Monocotyledonae. Monogr. Syst. Bot. Missouri. Bot. Gard. 107. 983 p. [ Links ]
Recibido el 23 de octubre de 2014,
aceptado el 11 de diciembre de 2014.

