










Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.51 no.4 Córdoba dic. 2016
ANATOMÍA Y MORFOLOGÍA
Análise comparativa da anatomia da madeira de 41 espécies de Rubiaceae sob enfoque taxonômico
Talita Baldin¹, Anelise Marta Siegloch², José Newton Cardoso Marchiori³ e Luciano Denardi4
¹ Doutoranda no programa de Pós-graduação em Engenharia Florestal na Universidade Federal de Santa Maria, UFSM. talita.baldin@hotmail.com
² Doutoranda no programa de Pós-graduação em Engenharia Florestal na Universidade Federal de Santa Maria, UFSM. anesiegloch@yahoo.com.br
³ Professor Titular do Departamento de Ciências Florestais na Universidade Federal de Santa Maria, UFSM. marchiori@pq.cnpq.br
4 Professor Associado do Departamento de Ciências Florestais da Universidade Federal de Santa Maria, UFSM. lucianodenardi@yahoo.com.br
Resumo
Rubiaceae é uma das famílias mais complexas do ponto de vista taxonômico, fato comprovado pelas inúmeras alterações ocorrentes ao longo dos anos na sua organização interna. O presente estudo investiga a madeira de 41 espécies, com base na revisão de literatura e verifica através da análise multivariada a formação de grupos e posicionamento das espécies nas subfamílias Ixoroideae, Rubioideae e Cinchonoideae. Pode-se verificar que o grupo apresenta-se estruturalmente homogêneo, devido ao grande número de caracteres anatômicos compartilhados, como já havia sido afirmado por muitos pesquisadores da área. As espécies não se agruparam totalmente de acordo com suas subfamílias. A análise também se mostra favorável a passagem do gênero Calycophyllum para a subfamília Cinchonoideae e as espécies de Calycophyllum permanecem unidas no dendrograma, evidenciando a homogeneidade do gênero.
Palavras chave: Análise de agrupamento; Anatomia da madeira; Cinchonoideae; Lxoroideae; Rubioideae; Taxonomia.
Summary
Comparative analysis of wood anatomy in forty one species of Rubiaceae with taxonomic emphasis
Rubiaceae is one of the most complex families from a taxonomic point of view, a fact proven by the numerous changes occurring over the years in their internal organization. Based on a literature review, this study investigates the wood of 41 species and utilizes multivariate analysis to verify group formation and placement of species in the subfamilies Ixoroideae, Rubioideae and Cinchonoideae. It can be seen that the group presents structurally homogeneous due to the large number of shared anatomical characters, a fact that has often been stated by others. The species do not completely group according to their subfamilies. The analysis also suggests the change of Calycophyllum genera for Cinchonoideae subfamily while the species Calycophyllum remain united in the dendrogram, demonstrating the homogeneity of the genera.
Key words: Cluster analysis; Wood anatomy; Cinchonoideae; Lxoroideae; Rubioideae; Taxonomy.
Introdução
A família Rubiaceae Juss., é a quarta em número de espécies no mundo, superada apenas por Orchidaceae Juss., Asteraceae Bercht. & J.Presl e Fabaceae Lindl. (Judd et al., 2009). De grande representatividade entre as Angiospermas, reúne cerca de 650 gêneros e 13.000 espécies (Delprete & Jardim, 2012), localizadas nas regiões mais quentes do globo, principalmente nos trópicos, onde se encontram mais de 75% das mesmas (Chiquieri et al., 2004).
As inúmeras tentativas de reestruturação da família, verificadas ao longo dos anos, comprovam sua alta complexidade do ponto de vista taxonômico. A classificação sistemática define que a família Rubiaceae pertence ao grupo das Asteridae, subgrupo das Euasteridae I, ordem Gentianales (APG III, 2009). Pesquisas filogenéticas recentes, com base em dados moleculares, sugerem a sua segregação em três subfamílias: Rubioideae, Cinchonoideae e Ixoroideae, e, até o presente momento, 42 tribos foram catalogadas. Tais concepções são igualmente referidas por pesquisadores dedicados à resolução de problemas taxonômicos na família, tais como: Andersson (1995), Bremer et al. (1995), Delprete (1996), Lens et al. (2000), Jansen et al. (2002), Robbrecht & Manen (2006), Bremer & Eriksson (2009) e Delprete & Jardim (2012).
Apesar das numerosas pesquisas correlatas à família Rubiaceae, a interpretação dos relacionamentos entre as diferentes espécies ainda está longe de consenso, evidenciando a necessidade de estudos mais aprofundados. Corroborando o exposto, Bremer (2009) salienta que para uma classificação mais definitiva e o melhor entendimento das circunscrições dos subgrupos, torna-se necessária a análise de um número maior de gêneros, cabendo salientar que, até o presente, mais de 200 ainda não foram abordados em estudos moleculares, morfológicos e anatômicos.
Vale lembrar que a anatomia da madeira tem sido empregada em diversos sistemas de classificação vegetal, tais como os de Takhtajan, como os de Gleason & Cronquist (1968), Takhtajan (1980) e Cronquist (1988), que se valeram de caracteres anatômicos como complemento ao estudo da taxonomia e filogenia das plantas. Em Rubiáceas, investigações dessa natureza demonstraram, claramente, a importância do lenho para a separação dos taxa (Lens et al., 2000). No presente estudo foi analisada a anatomia da madeira de 41 espécies de Rubiaceae, com o uso da análise de agrupamento (taxonomia numérica), para verificar a formação de grupos de afinidades anatômicas, enfatizando a opinião da anatomia da madeira nas classificações recentes da família.
Material e Métodos
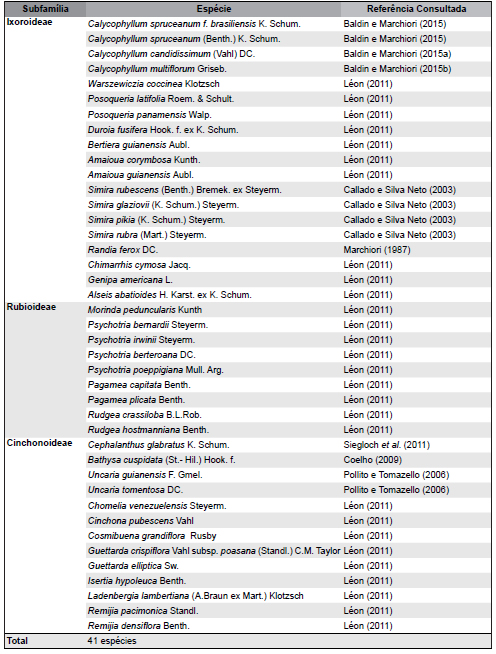
Para o estudo foram compiladas características anatômicas do lenho de 41 espécies de Rubiaceae (10 espécies brasileiras e 31 espécies venezuelanas), das subfamílias Ixoroideae (19 sp.), Rubioideae (9 sp.), e Cinchonoideae (13 sp.), disponíveis em periódicos científicos (Tabela 1), em publicações de Marchiori (1987), Callado & Silva Neto (2003), Pollito & Tomazello (2006), Coelho et al. (2012), Léon (2011), Siegloch et al. (2011) e Baldin & Marchiori (2015, 2015a, b).
Tabela 1. Espécies de Rubiaceae utilizadas na análise de agrupamento e suas respectivas subfamílias.
Table 1. Rubiaceae species used in the cluster analysis and their subfamilies.
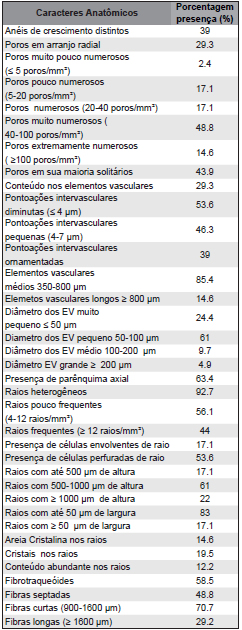
Para comparar e reunir tais espécies em grupos de afinidade anatômicas, realizou-se a análise de agrupamento hierárquica aglomerativa, com base nas características com ocorrência entre as espécies, de valor taxonômico e considerados como típicos de Rubiaceae por Record & Hess (1949), Metcalfe & Chalk (1972), Koek-Noorman (1977) e Jansen et al. (2002). Salienta-se que os caracteres ocorrentes em 100% dos exemplares foram excluídos da análise (Tabela 2).
Tabela 2. Caracteres anatômicos de ocorrência nas espécies investigadas.
Table 2. Anatomical characters occurrence in species.
Tal análise foi gerada a partir de uma matriz de distância de dissimilaridade de Jaccard, utilizando-se para a geração dos grupos o método de peso proporcional (Unweigthed Pair-Group Method using Arithmetical Averages - Upgma). Para a validação do dendrograma, calculou-se o coeficiente de correlação cofenético (r). Os dados e análises foram processados no software R (R Development Core Tean 2011), pacote vegan (Oksanen et al., 2012).
Para verificar quais são as variáveis anatômicas de cunho quantitativo que mais explicam a variância dos dados e importantes para a separação das espécies, foi realizado uma análise multivariada discriminante, por meio do software The Unscrambler. Os dados anatômicos quantitativos foram submetidos ao processo de normalização, e, posteriormente, submetidos à análise de componentes principais (ACP), analisando-se as covariâncias e as correlações baseadas nas raízes (ou valores) das variáveis e em seus vetores, considerando as duas primeiras raízes extraídas, por apresentaram os maiores autovalores (Ayres et al., 2000).
Todos os nomes científicos referenciados na pesquisa foram consultados no Ipni, The International Plant Names Index – www.ipni.org (acessado em 17.10.2015) e a classificação Botânica utilizada segue o APG III (2009).
Resultados
Características anatômicas do lenho de 41 espécies de Rubiaceae
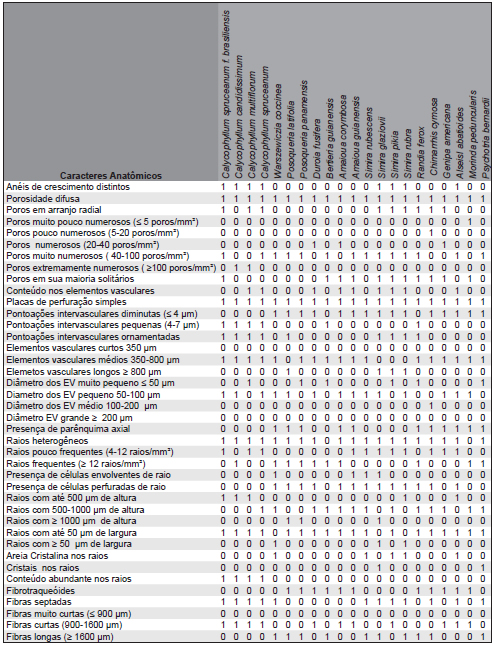
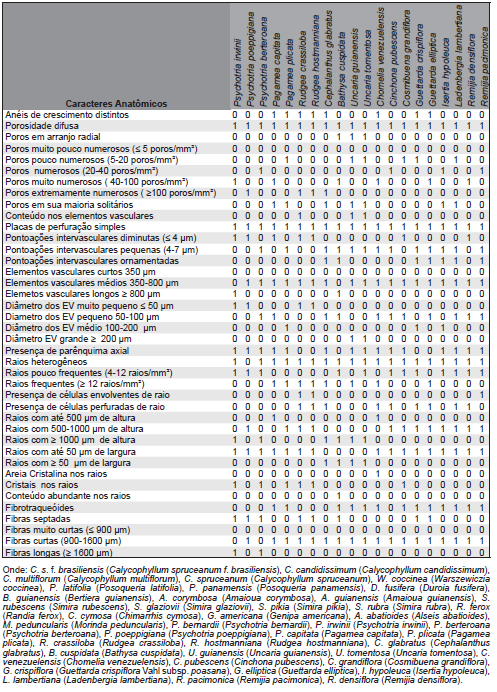
As características anatômicas qualitativas e quantitativas do lenho das espécies investigadas são reportadas na Tabela 3.
Tabela 3. Caracteres anatômicos presente (1) ou ausente (0) nas 41 espécies investigadas.
Table 3. Present anatomical characters (1). Absent character (0) in 41 species investigated.
Análise comparativa
No dendrograma (Fig. 1), o nível de distância com que as espécies se afastam ou se aproximam de 1 pode ser lido diretamente na horizontal, em dissimilaridade. Quanto maior o índice, menor a proximidade entre as espécies. O valor do coeficiente de correlação cofenético, de 0,73, de acordo com Valentin (2000), indica que o dendrograma é relevante na análise.
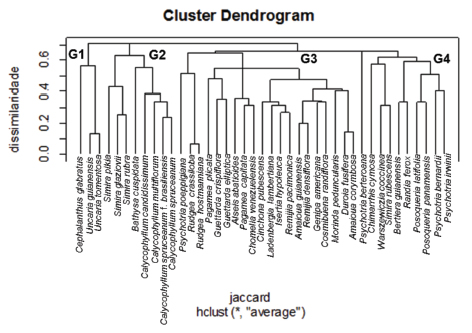
Fig. 1. Dendrograma de agrupamento, com base na distância de Jaccard e método UPGMA.Onde: G1, G2, G3 e G4 representam os grupos formados.
Fig. 1. Clustering dendrogram based on Jaccard distance and UPGMA. At where: G1, G2, G3 e G4 represent the formed groups.
Análise de componentes principais (PCA)
A análise de componentes principais da matriz de dados dos caracteres anatômicos quantitativos apontou o primeiro componente como responsável por 61,27%, o segundo como 85,39% e o terceiro como 93,04% de toda a variância dos dados. Com três eixos foi possível explicar a relação entre os caracteres anatômicos quantitativos e as espécies de Rubiaceae investigadas (Fig. 2).
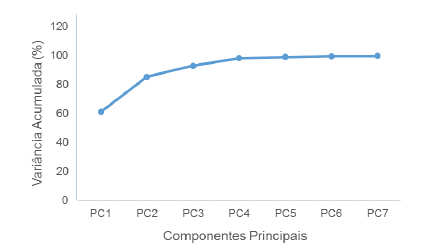
Fig. 2. Análise de componentes principais com base na matriz de caracteres anatômicos quantitativos em função das espécies investigadas.
Fig. 2. Principal component analysis based on the quantitative characteres matrix for species.
As espécies formaram 4 (G1, G2, G3 e G4) grupos distintos (Fig. 3) onde o peso de cada variável anatômica quantitativa, nos dois fatores de maior autovalor, ou seja, que melhor explicam a variação observada (Tabela 4).

Fig.3. Análise de função discriminante com a indicação dos grupos de espécies constituídos pelas características anatômicas quantitativas.Onde 1: P.l: Posoqueria latifólia; P.p: Posoqueria panamensis; P.b: Psychotria Bernardi; P.i: Psychotria irwinii e S.rub: Simira rubescens 2: U.g: Uncaria guaianensis e U.t: Uncaria tomentosa 3: C.g: Cephalanthus glabratus e B.c: Bathysa cuspidata 4: C.b: Calycophyllum spruceanum f. brasiliensis; C.c: Calycophyllum candidissimum; C.m: Calycophyllum multiflorum; C.s: Calycophyllum spruceanum; D.f: Duroia fusifera; B.g: Bertieria guianensis; A.c: Amaioua corymbosa; A.g: Amaioua guianensis; G.a: Genipa americana; A.a: Alseisl abatioides; M.p: Morinda peduncularis; P.p: sychotria poeppigiana; P.c: Pagamea capitata; P.p: Pagamea plicata; R.c: Rudgea crassiloba; R.h: Rudgea hostmanniana; S.p: Simira pikia; C.v: Chomelia venezuelensis; C.p: Cinchona pubescens; C.g: Cosmibuena grandiflora; G.c: Guettarda crispiflora; G.e: Guettarda elliptica; I.h: Isertia hypoleuca; L.l: Ladenbergia lambertiana; R.d: Remijia densiflora e R.p: Remijia pacimonica 5: S.g: Simira glaziovii; S.rubra: Simira rubra; P.b: Psychotria berteroana; W.c: Warszewiczia coccinea e C.c: Chimarrhis cymosa.
Fig. 3. Discriminant function analysis with the indication of groups of species constitute by the quantitative characteristics. Where: P.l: Posoqueria latifólia; P.p: Posoqueria panamensis; P.b: Psychotria Bernardi; P.i: Psychotria irwinii e S.rub: Simira rubescens 2: U.g: Uncaria guaianensis e U.t: Uncaria tomentosa 3: C.g: Cephalanthus glabratus e B.c: Bathysa cuspidata 4: C.b: Calycophyllum spruceanum f. brasiliensis; C.c: Calycophyllum candidissimum; C.m: Calycophyllum multiflorum; C.s: Calycophyllum spruceanum; D.f: Duroia fusifera; B.g: Bertieria guianensis; A.c: Amaioua corymbosa; A.g: Amaioua guianensis; G.a: Genipa americana; A.a: Alseisl abatioides; M.p: Morinda peduncularis; P.p: sychotria poeppigiana; P.c: Pagamea capitata; P.p: Pagamea plicata; R.c: Rudgea crassiloba; R.h: Rudgea hostmanniana; S.p: Simira pikia; C.v: Chomelia venezuelensis; C.p: Cinchona pubescens; C.g: Cosmibuena grandiflora; G.c: Guettarda crispiflora; G.e: Guettarda elliptica; I.h: Isertia hypoleuca; L.l: Ladenbergia lambertiana; R.d: Remijia densiflora e R.p: Remijia pacimonica 5: S.g: Simira glaziovii; S.rubra: Simira rubra; P.b: Psychotria berteroana; W.c: Warszewiczia coccinea e C.c: Chimarrhis cymosa.
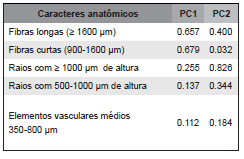
Tabela 4. Fatores de peso dos caracteres anatômicos quantitativos na separação dos grupos de espécies pela análise de componentes principais.
Table 4. Loading factors of the quantitative chacarteres in separation of the groups of specie by Principal Component Analyses.
Discussão
Os resultados das análises evidenciaram variações significativas dos caracteres quantitativos da madeira, demonstrando sua eficácia na diferenciação das espécies florestais estudadas (Fig. 3). Os fatores que mais contribuíram com variância dos dados explicada na análise de componentes principais foram relativos ao comprimento de fibras, a altura dos raios e o comprimento dos elementos vasculares (Tabela 4).
A variação no comprimento das fibras entre as espécies é caráter esperando em análises anatômicas, posto que, a dimensão do tecido fibroso está altamente correlacionada com a idade da árvore. A estabilização do comprimento das fibras, conforme observado por Bisset & Dadswell (1950) somente irá ocorrer quando as células do câmbio atingirem comprimento máximo, iniciando a formação da madeira caracterizada como adulta.
Quanto à altura dos raios, Metcalfe & Chalk (1972) menciona o caráter como bastante variável em Rubiaceae. A variação na altura de raios pode ser igualmente reportada para as subfamílias Cinchonoideae, Ixoroideae e Rubioideae (Record & Hess, 1949).
As espécies foram agrupadas em quatro grupos principais: (G1), 3 espécies da subfamília Cinchonoideae; (G2), por 8 espécies de Ixoroideae e Cinchonoideae; (G3), o maior grupo, constituído por 20 espécies, pertencentes as três subfamílias e (G4) por 10 representantes de Ixoroideae e Rubioideae (Fig. 1). Os grupos de espécies de afinidade anatômica não seguiram as recentes classificações da família, não se reuniram, na totalidade, em suas respectivas subfamílias, demonstrando certa homogeneidade das características anatômicas.
No grupo G1, verificou-se a formação do agrupamento entre espécies de Cinchonoideae, onde Uncaria guianensis e Uncaria tomentosa, estão ligadas a uma similaridade de aproximadamente 0,55 com Cephalanthus glabratus. Embora pertencentes à mesma subfamília, poucos caracteres anatômicos são comuns às três espécies, caso de poros em sua maioria solitários, presença de parênquima axial, e raios altos, muito largos.
O grupo dois (G2) compreende Simira pikia, S. glaziovii e S. rubra (Ixoroideae), pertencentes à tribo Simireae, tribo considerada homogênea no tocante a estrutura anatômica do lenho (Callado & Silva Neto, 2003), o que corrobora a presente análise, como observado na Fig. 1, as espécies são altamente similares (agrupadas, a 0,4 de dissimilaridade). As três espécies do gênero Simira estão ligadas com Bathysa cuspidata (Cinchonoideae) e com as quatros espécies de Calycophyllum pertencente a Ixoroideae (C. spruceanum f. brasiliense, C. candidissimum, C. multiflorum e C. spruceanum).
Na presente análise, dendrograma (Fig. 1) ocorreu o agrupamento das espécies de Calycophyllum, pertencentes a tribo Condamineeae, revelando a homogeneidade do gênero dentro da tribo à qual faz parte. Todavia, estudos de Koek- Noorman & Hogeweg (1974), com os gêneros Bikkia, Morierina e Portandia, mencionam a heterogeneidade de Condamineeae no que se refere à composição anatômica da madeira.
No agrupamento de Calycophyllum, o menor valor de dissimilaridade (cerca de 0,2) é evidenciado entre Calycophyllum spruceanum f. brasiliensis e C. spruceanum; resultados que confirmam a conveniência da proposição de K. Schum. e Müll. Arg. (Chiquieri et al., 2004), que consideraram o primeiro como variedade do segundo táxon. As características anatômicas que reforçam esse entendimento são: poros com diâmetro tangencial médio (50-100 μm), muito numerosos e em múltiplos radiais; raios heterogêneos, pouco frequentes, predominantemente trisseriados e tetrasseriados.
No presente estudo, confirma-se, a homogeneidade das quatro espécies de Calycophyllum (Fig. 1), devido as características anatômicas compartilhadas: anéis de crescimento distintos, marcados visualizados por fibras de menor diâmetro radial e paredes mais espessas ao final do lenho tardio; porosidade difusa; poros muito numerosos a extremamente numerosos; placas de perfuração simples; pontoações intervasculares pequenas e ornamentadas; raios heterogêneos, com até 1000 μm de altura, e 50 μm de largura, com células perfuradas, abundante areia cristalina e conteúdos; e fibras septadas médias, de paredes delgadas a espessas.
As espécies de Calycophyllum (C. spruceanum f. brasiliensis, C. candidissimum, C. multiflorum, C. spruceanum) pertencentes atualmente à subfamília Ixoroideae (Bremer & Jansen, 1991), na presente análise, juntam-se a Bathysa cuspidata, espécie da subfamília Cinchonoideae, devido ao compartilhamento de várias características anatômicas: porosidade difusa; poros muito numerosos e em arranjo radial; placas de perfuração simples; pontoações intervasculares pequenas e ornamentadas; parênquima axial ausente; raios heterogêneos pouco frequentes, com abundante conteúdo em suas células; e fibras septadas médias.
Quanto à delimitação em nível de subfamília, todavia, o gênero Calycophyllum não atende aos padrões citados por Koek-Noorman (1977) e Jansen et al. (1997, 1999) para Ixoroideae: vasos solitários, ou menos comumente em pequenos múltiplos radiais, fibrotraqueóides, e parênquima axial difuso, difuso-em-agregados e em pequenas linhas marginais. Sobre este ponto, cabe salientar, que o gênero Calycophyllum, com base apenas nas características anatômicas, deveria permanecer na antiga subfamília Cinchonoideae, como o proposto inicialmente por Schumann (1891).
O mais abrangente eixo, corresponde ao grupo três (G3), apresenta cinco ramificações notavelmente distintas: A primeira, com representantes essencialmente de Rubioideae, tais como Psychotria poeppigiana e Rudgea (R. crassiloba e R. hostmanniana), agrupa suas espécies segundo as respectivas subfamílias, evidenciando o compartilhamento de grande parte dos caracteres anatômicos. As duas espécies do gênero Rudgea inclusive, são as mais similares dentro das 41 analisadas, com índice de 0,05.
Na segunda ramificação, Pagamea plicata (Rubioideae) distancia-se em cerca de 0,3 de Guettarda crispiflora e G. elliptica (Cinchonoideae). No segundo ramo, com o mesmo índice de dissimilaridade, agrupam-se Alseis abatioides (Ixoroideae) com Pagameae capitata (Rubioideae) e Chomelia venezuelensis (Cinchonoideae). A terceira ramificação evidente no grupo três, demonstra claramente a organização da subfamília Cinchonoideae, por Cinchona pubescens, Isertia hypoleuca, Ladenbergia lambertiana, Remijia pacimonica, com disimilaridade de 0,3.
Na quarta ramificação a similaridade, respectivamente, entre espécies de Ixoroideae (Amaioua guianensis e Genipa americana) com Cinchonoideae (Remijia densiflora e Cosmibuena randiflora). A quinta e última ramificação agrupa Morinda peduncularis (Rubioideae) com Duroia fusifera e Amaioua corymbosa, ambas Ixoroideae.
No agrupamento quatro (G4) Psychotria berteroana, pertencente à subfamília Rubioideae, mostra um índice de dissimilaridade de 0,6, relativamente às demais espécies do gênero (P. bernardii e P. irwinii). A separação de P. berteroana pode ser explicada por caracteres anatômicos peculiares de sua madeira, que fogem à tendência geral para a família, tais como: poros pouco numerosos; pontoações intervasculares diminutas; elementos vasculares de diâmetro reduzido; raios muito altos e pouco frequentes, com presença de ráfides em suas células.
Cabe destacar na análise, a separação do gênero Simira, organizado em dois grupos distintos: em G2, Simira glaziovii, S. pikia e S. rubra; e em G4, S. rubescens. Essa separação decorre de diferenças anatômicas como: anéis de crescimento indistintos; poros em sua maioria solitários; elementos vasculares de tamanho médio, e com conteúdo em S. rubescens; poros em arranjo radial, e com pontoações ornamentadas em Simira glaziovii, S. pikia e S. rubra.
No grupo G3 e G4 verifica-se, ainda, a proximidade de Ixoroideae e Rubioideae, o que justifica as constantes reorganizações na taxonomia das Rubiaceae, principalmente a nível de subfamília, onde inúmeras espécies inicialmente inseridas em Rubioideae (Verdcourt, 1958) passaram às Ixoroideae (Robbrecht, 1988).
Pode-se concluir que os caracteres anatômicos quantitativos que melhor discriminam as espécies referem-se ao comprimento de fibras, altura de raios e comprimento de elemento vascular.
A análise de agrupamento demostra ser útil na comparação e separação das espécies investigadas. Notavelmente, as 41 espécies de Rubiaceae dividiram-se em quatro grupos principais, e de maneira geral, não formam grupos de acordo com as suas respectivas subfamílias, evidenciando a homogeneidade dos caracteres anatômicos do lenho.
Evidencia-se a proximidade de Ixoroideae e Rubioideae, justificando, pelo menos em parte, as constantes reorganizações na taxonomia das Rubiaceae no tocante às subfamílias.
A anatomia da madeira se inclina, favoravelmente, à permanência do gênero Calycophyllum em sua antiga subfamília (Cinchonoideae).
Bibliografía
1. AYRES, M. AYRES, M. Jr., AYRES, D. L., SANTOS, A. A. S. DOS. 2000. BioEstat 2.0: Aplicações estatísticas nas áreas de ciências biológicas e médicas. CNPq, Brasília. [ Links ]
2. ANDERSSON, L. & J. H. E. ROVA. 1999. The rps intron and the phylogeny of the Rubioideae (Rubiaceae). Plant Syst. Evol. 214: 161-186. [ Links ]
3. BALDIN, T. & J. N. C. MARCHIORI. 2015. Anatomia da madeira de Calycophyllum spruceanum (benth.) K. SCHUM. e Calycophyllum spruceanum f. brasiliensis K. SCHUM. Balduinia 50: 16-23. [ Links ]
4. BALDIN, T. & J. N. C. MARCHIORI. 2015a. Anatomia da madeira de Calycophyllum candidissimum (Vahl) DC. (Rubiaceae). Balduinia 48: 23-28. [ Links ]
5. BALDIN, T. & J. N. C. MARCHIORI. 2015b. Descrição anatômica de Calycophyllum multiflorum Griseb. (Rubiaceae). Balduinia 49: 16-23. [ Links ]
6. BISSET, J. J. W. & DADSWELL, H. E. 1950. The variation in cell length within one growth ring of certain angiosperms and gymnosperms. Austral. Forestry 15: 17-29. [ Links ]
7. BREMER, B., K. ANDREASEN. & D. OLSSON. 1995. Subfamilial and tribal relationships in the Rubiaceae based on rbcL sequence data. Ann. Missouri Bot. Gard. 82: 383-397. [ Links ]
8. BREMER, B. 2009. A review of molecular phylogenetic studies of Rubiaceae. Ann. Missouri Bot. Gard. 96: 4-26. [ Links ]
9. BREMER, B. & O. ERIKSSON. 2009. Time tree of Rubiaceae: phylogeny and dating the family, subfamily, and tribes. Int. J. Plant Sci. 170: 766-793. [ Links ]
10. BREMER, B. & R. K. JANSEN. 1991. Comparative restriction site mapping of chloroplast DNA implies new phylogenetic relationships within Rubiaceae. Amer. J. Bot. 78: 198-213. [ Links ]
11. CALLADO, C. H. & S. J. SILVA NETO. 2003. Anatomia do lenho de três espécies do gênero Simira Aubl. (Rubiaceae) da Floresta Atlântica no estado do Rio de Janeiro. Rodriguésia 54: 23-33. [ Links ]
12. CHIQUIERI, A., F. R. DI MAIO. & A. L. PEIXOTO. 2004. A distribuição geográfica da família Rubiaceae Juss. na Flora Brasiliensis de Martius. Rodriguésia 55: 47-57. [ Links ]
13. COELHO, V. P. M., J. P. V. LEITE., L. G. NUNES. & M. C. VENTRELLA. 2012. Anatomy, histochemistry and phytochemical profile of leaf and stem bark of Bathysa cuspidata (Rubiaceae). Austral. J. Bot. 60: 49. [ Links ]
14. CRONQUIST, A. 1988. The Evolution and Classification of Flowering Plants. 2nd. ed. The New York Botanical Garden, New York. [ Links ]
15. DELPRETE, P.G. 1996. Evaluation of the tribes Chiococceae, Condamineeae and Catesbaeeae (Rubiaceae) based on morphological characters. Opera Bot. Belg. 7: 165-192. [ Links ]
16. DELPRETE, P. G. & J. G. JARDIM. 2012. Systematics, taxonomy and floristics of Brazilian Rubiaceae: an overview about the current status and future challenges. Rodriguésia 63: 101-128. [ Links ]
17. GLEASON, H. A. & A. CRONQUIST. 1968. The Pteridophyta, Gymnospermae and Monocotyledoneae. The New Britton and Brown Illustrated Flora of the North eastern United States and Adjacent Canada 1: 1-482. The New York Botanical Garden, New York. [ Links ]
18. IAWA COMMITTEE. 1989. IAWA list of microscopic features for hardwood identificacion. IAWA Bull. 10: 218-359. [ Links ]
19. JANSEN, S., E. ROBBRECHT., H. BEECKMAN. & E. SMETS. 2002. A survey of the systematic wood of the Rubiaceae. IAWA Bull. 23: 1-67. [ Links ]
20. JUDD, W. S., C. S. CAMPBELL. & E. A. KELLOGG. 2009. Sistemática vegetal: Um enfoque Filogenético. 3ed. Artmed, Porto Alegre. [ Links ]
21. KOEK-NOORMAN, J. 1977. Systematische holzanatomie einiger Rubiaceen. Ber. Deut. Bot. Ges. 90: 183-190. [ Links ]
22. KOEK-NOORMAN, J. & P. HOGEWEG. 1974. The wood anatomy of Vanguerieae, Cincohoneae, Codaminae, and Rondeletieae (Rubiaceae). Acta Bot. Neerl. 23: 627- 653. [ Links ]
23. LENS, F., S. JANSEN., E. ROBBRECHT. & E. SMETS. 2000. Wood anatomy of the Vanguerieae (Ixoroidea-Rubiaceae), with special emphasis on some geofrutices. IAWA Bull. 21: 443-455. [ Links ]
24. LEÓN. H. W. 2011. Anatomía de la madera de 31 especies de Rubiaceae en Venezuela. Acta Bot. Venez. 34: 347-379. [ Links ]
25. MÓL, F. F. D. 2010. Rubiaceae em um remanescente de floresta atlântica no Rio Grande do Norte, Brasil. Dissertação. Universidade Federal do Rio Grande do Norte, Natal. [ Links ]
26. MARCHIORI, J. N. C. 1987. Anatomia da madeira de limoeiro-do-mato, Randia armata (Sw.) DC. Ciência Rural 17: 39-48. [ Links ]
27. METCALFE, C. R. & L. CHALK. 1972. Anatomy of the Dicotyledons. Clarendon Press, Oxford. [ Links ]
28. OKSANEN, J., F. G. BLANCHET., R. KINDT., P. LEGENDRE., R. B. O’HARA., G. L. SIMPSON., M. H. H. STEVENS. & H. WAGNER. 2011. Vegan: community ecology package. Version 2.0- 6 (http://vegan.r-forge.r-project.org/). Acesso em 08/09/2015.
29. POLLITO, P. A. Z. & M. TOMAZELLO. 2006. Anatomia do lenho de Uncaria guianensis e U. tomentosa (Rubiaceae) do estado do Acre, Brasil. Acta Amazôn. 36: 169-176. [ Links ]
30. R DEVELOPMENT CORE TEAM. 2011. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Disponible en: http://www.R-project.org. [ Links ]
31. RECORD, S. J. & R. W. HESS. 1949. Timbers of The New World. Yale University Press, New Haven. [ Links ]
32. ROBBRECHT, E. 1988. Tropical wood Rubiaceae – Characteristic features and progressions. Contribuitions to a new subfamilial classification. Opera Bot. Belg. 1: 1-271.
33. ROBBRECHT, E. & J. F. MANEN. 2006. The major evolutionary lineages of the coffe family (Rubiaceae, angiosperm). Combined analysis (nDNA and cpDNA) to infer the position of Coptosapelta and Lucilia, and supertree construction based on rbcL, rps16, trnL-trnF and atpB-rbcL data. A new classification in two subfamilies, Cinchonoideae and Rubioideae. Syst. Geogr. Plant 76: 85-146. [ Links ]
34. SCHUMANN, K. 1891. Rubiaceae. In: Engler, A. & Prantl, K. (eds.), Die natürlichen Pflanzenfamilien 4: 1-156. Engelmann, Leipzig. [ Links ]
35. SIEGLOCH, A. M., J. N. C. MARCHIORI. & S. R. SANTOS. 2011. Anatomia do lenho de Cephalanthus glabratus (Spreng.) K. Schum. (Rubiaceae). Balduinia 31: 20-26. [ Links ]
36. TAKHTAJAN, A. 1980. Outline of the Classification of Flowering Plants (Magnoliophyta). Bot. Rev. 46: 225-359. [ Links ]
37. THE ANGIOSPERM PHYLOGENY GROUP. 2009. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161: 105-121. [ Links ]
38. THE INTERNATIONAL PLANT NAMES INDEX - IPNI. Published on the Internet (http://www.ipni.org). Accessed 17/10/2015. [ Links ]
39. VALENTIN, J. L. 2000. Ecologia numérica: uma introdução à análise multivariada de dados ecológicos. pp. 117. Interciência, Rio de Janeiro. [ Links ]
40. VERDCOURT, B. 1958. Remarks on the Classification of the Rubiaceae. Bull. Jard. Bot. État Bruxelles 28: 209-290. [ Links ]
Recibido el 21 de abril de 2016,
aceptado el 13 de octubre de 2016.

