










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versión On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.53 no.1 Córdoba mar. 2018
Fisiología - Physiology
Efecto de la densidad de siembra sobre la actividad fotosintética en Chenopodium quinoa var. CICA (“quínoa”) en el Noroeste Argentino.
Sowing density effect on photosynthetic activity of Chenopodium quinoa var. CICA (“quínoa”) in the Argentinean Northwest.
JUAN A. GONZÁLEZ1*, LUIS E. ERAZZÚ2, SEBASTIÁN E. BUEDO1 y FERNANDO E. PRADO3
1 Instituto de Ecología, Fundación Miguel Lillo, Miguel Lillo 251, Tucumán (Argentina).
2 Instituto Nacional de Tecnología Agropecuaria (INTA), Famaillá, Tucumán, Argentina, Facultad de Agronomía y Zootecnia, Universidad Nacional de Tucumán.
3 Instituto de Bioprospección y Fisiología Vegetal (INBIOFIV), CONICET-UNT. Cátedra de Fisiología Vegetal, Facultad de Cs. Naturales e IML, Miguel Lillo 205, Tucumán, Argentina.
* jagonzalez@lillo.org.ar
Resumen: El cultivo de la quinoa para la obtención de granos para la alimentación humana ha comenzado a extenderse en la región de los Valles Calchaquíes (Tucumán, Argentina) por arriba de los 2.000 m snm. Tratándose de un cultivo nuevo para dicha región son necesarios los estudios ecofsiológicos y agronómicos para lograr un manejo adecuado de esta nueva alternativa productiva. En este estudio se analiza el efecto de la densidad de siembra sobre la respuesta fotosintética frente a diferentes densidades de fujo fotosintético de fotones (DFFF) estableciéndose los valores de asimilación máxima (Amax), punto de saturación lumínica (PSL), punto de compensación lumínica (PCL), eficiencia fotosintética (ØCO2) y respiración nocturna (Rn). Por otro lado, se proporcionan datos relacionados con la asimilación fotosintética neta (An), conductancia estomática (gs), transpiración foliar (Tr), concentración interna de CO2 (Ci), capacidad de carboxilación (An/Ci) y eficiencia intrínseca en el uso del agua (EUAi). Asimismo se brindan datos sobre el contenido de pigmentos fotosintéticos (cloroflas y carotenoides) y otros compuestos protectores (que absorben a los 305 nm). Este estudio demuestra que los parámetros del intercambio de gases están directamente correlacionados con la morfología, anatomía foliar y rendimiento en granos y que la densidad de siembra tiene un efecto directo sobre ellos.
Palabras clave: Chenopodium, densidad de siembra, fotosíntesis, pigmentos.
Summary: Sowing density effect on photosynthetic activity of Chenopodium quinoa var. CICA ("quinoa") in the Argentinean Northwest. Quinoa crop to obtain grain for human consumption has begun to spread in the region of Calchaquíes Valleys (Tucumán, Argentina) over 2,000 m asl. Because quinoa is a new alternative crop, must be carried out ecophysiological and agronomic studies to get its adequate management. In this study the effect of sowing density on the photosynthetic response against different photon fux densities (DFFF) was analyzed in order to calculate the maximum assimilation (Amax), light saturation point (PSL), light compensation point (PCL), photosynthetic efficiency (ØCO2) and dark respiration (Rn) values. Data in relation to net photosynthetis assimilacion (An), stomatal conductance (gs), leaf transpiration (Tr), internal CO2 concentration (Ci), carboxilation capacity (An/Ci) and intrinsec water use efficiency (EUAi) are provided. In addition, photosynthetic pigment (chlorophylls and carotenoids) and protective one (absorbance at 305 nm) were measured. This study demonstrates that gas exchange parameters are directly correlated with morphology, foliar anatomy and grains yield and that the sowing density has a direct effect on them.
Key words: Chenopodium, sowing density, photosynthesis, pigments.
introducción
En un estudio previo sobre Chenopodium quinoa Willd. ("quinoa") (var. CICA) realizado en condiciones de campo, se demostró que la densidad de siembra afectó tanto la morfología externa como la anatomía de la hoja y el rendimiento de granos (Erazzú et al., 2016). En efecto, utilizando dos densidades de siembra (4,2 y 27,9 plantas/m lineal con entresurcos a 0,50 m), se encontró que tanto la altura de las plantas como el diámetro del tallo resultaron mayores en la densidad más baja. El área foliar específica (AFE), medida indirecta del espesor de la lámina foliar; así como la densidad estomática, variable importante en el intercambio de gases, también resultaron incrementadas en el tratamiento de densidad baja. En esta misma condición de densidad, el contenido foliar de nitrógeno y fósforo también mostraron un incremento en su concentración (Erazzú et al., 2016). Es conocido que el rendimiento de un cultivo depende de la densidad de plantas sembradas, debido a la competencia que se establece entre ellas por el agua y los nutrientes fundamentalmente, pero también por la captura de la radiación fotosintéticamente activa (RFA) (Milthorpe & Moorby, 1982; Lambers et al., 2008). La densidad de siembra afecta los parámetros morfológicos externos y los anatómicos, por lo que resulta esperable que los procesos fsiológicos, como la fotosíntesis, también resulten afectados. De manera que el objetivo de este estudio fue evaluar el efecto de dos densidades de siembra sobre los parámetros fotosintéticos del cultivar CICA de "quinoa", cultivado en una zona árida montañosa de Tucumán (Argentina); con la finalidad de correlacionar dichos parámetros con el rendimiento en granos. Esta evaluación resulta importante puesto que la actividad fotosintética, al ser el refejo de las condiciones ambientales en la que se desarrolla una especie, puede ser un indicador del rendimiento. Por otro lado, este tipo de estudios en una especie como "quinoa" son aún incipientes o escasos en la región montañosa del NOA, por lo que su utilidad a nivel biológico o agronómico es importante para el conocimiento ecofsiológico de las diferentes variedades (nativas e introducidas) que se están ensayando tanto en Encalilla (Amaicha del Valle, Tucumán) como en el resto del denominado Valle Calchaquí que cubre porciones del territorio de
Catamarca, Tucumán, y Salta. El conocimiento que brindan estos estudios permitirá la selección de variedades más aptas para su cultivo en función de los objetivos agronómicos que se persigan. Si bien este cultivo ha comenzado a desarrollarse en el NOA en la última década, el mismo puede ser una importante innovación debido a que representa una alternativa, o quizás una actividad complementaria, para los productores de la zona. En efecto, "quinoa" es una especie nativa de la zona y como tal presenta adaptaciones como la resistencia a la salinidad y también a sequía, que le confere ventajas sobre otras especies que no son de la región. También sería posible cultivarla en lugares donde las especies clásicas no pueden prosperar, como en tierras con alta salinidad, suelos marginales pobres en materia orgánica y otras condiciones microclimáticas. En la actualidad la mayoría de los cultivos en esta región están representados por especies no nativas (pimiento, tomates, frutas de carozo, avena, entre otras) y que poseen otros requerimientos agronómicos entre los cuales el agua, que es escasa en la zona, es el elemento que más consumen. De manera que el desarrollo del cultivo de "quinoa", destinada a la alimentación de personas por las propiedades nutricionales que posee (González et al., 1989; 2015), forrajes (González et al., 2016) o productos naturales para la industria como son las saponinas (Quiroga et al., 2014) podrían generar mercados que hasta el momento no existen como la provisión de alimentos libres de gluten para las personas que padecen celiaquía. Como todo nuevo cultivo, es necesario el conocimiento biológico y ecofisiológico, además de los agronómicos, para afianzarlo en una zona donde existe la oferta ambiental para realizarlo.
Materiales y Métodos
El estudio se llevó a cabo en el Campo Experimental de Encalilla (22º 31` S y 65º 59` O, Amaicha del Valle, Tucumán, Argentina). Este lugar se halla ubicado a 1.995 m snm y según la clasificación climática de Köppen corresponde al tipo desértico (BWkaw) (Minetti, 2005). El régimen anual de precipitaciones promedio es 220 mm. Las temperaturas máximas y mínimas (promedios diarios) registradas son de 30,4 y 11,2ºC respectivamente, mientras que la HR oscila entre el 44,2% y 54,2%. La radiación fotosintéticamente activa (RFA), a medio día, registrada durante este estudio, en condiciones de días soleados, osciló entre 1.403 y 1.993 μmol m-2 s-1. Como material vegetal se utilizaron semillas de quinoa variedad CICA provenientes de ensayos de años anteriores. Se realizaron dos tratamientos para lograr dos densidades de plantas por metro lineal. Los detalles de siembra para lograr las densidades 4,2 (“Densidad baja”) y 27,9 (“Densidad alta”) plantas/m lineal, así como las características fisicoquímicas y composición mineral del suelo fueron descritas en un trabajo anterior (Erazzú et al., 2016) donde se informó sobre el efecto de estas densidades sobre la morfología, anatomía de la hoja y rendimiento de granos. La siembra se realizó en bloques al azar con 4 repeticiones el 15 de noviembre de 2013. Cada parcela tuvo un largo de 5 metros con 11 surcos y la distancia entre los mismos fue de 0,50 m. Para estimar los parámetros evaluados, se procedió a su determinación en el sentido de las diagonales de cada parcela, evitando las filas externas para minimizar los efectos de borde. Todas las mediciones se realizaron al momento de la floración al estadio de flor abierta. Para las mediciones de los parámetros de intercambio gaseoso se seleccionaron hojas jóvenes completamente expandidas ubicadas en la parte superior del follaje. La asimilación fotosintética neta (An), conductancia estomática (gs), transpiración foliar (Tr) y concentración interna de CO2 (Ci), se midieron con un analizador infrarrojo de CO2 portátil equipado con cámara fluorométrica LICOR (LI-6400 XT, LiCor Inc. Nebraska, USA). La capacidad de carboxilación fue calculada como el cociente An/Ci mientras que la eficiencia intrínseca en el uso del agua (EUAi) como An/gs. Para el cálculo de EUAi se utilizó el cociente entre An y gs en vez de An y Tr para eliminar el efecto de la temperatura y el gradiente de humedad sobre la transpiración según la propuesta de Rawson et al. (1977). Las mediciones de An se realizaron bajo condiciones de luz y CO2 saturantes (1.500 μmol m-2 s-1, 400 μmol CO2) de acuerdo a Geissler et al. (2015) y temperatura foliar constante (25±0,5°C), entre las 9:00 y 11:00 h. Para estudiar la respuesta de la fotosíntesis neta (An) a intensidades distintas de densidad de flujo fotosintético de fotones (DFFF) se utilizó un programa, incorporado al analizador de gases mencionado, que genera automáticamente niveles de radiación entre 0 y 2.500 μmol m-2 s-1. A partir de las curvas An/DFFF se calcularon el punto de saturación lumínica (PSL) (definida como la DFFF a la cual An es igual al 90% de la Amax), el punto de compensación lumínica (PCL) que corresponde al valor de DFFF en el cual la An es igual a cero y la eficiencia cuántica de la fotosíntesis (ØCO2) (que es el número de moles de fotones necesarios para fijar un mol de CO2) de acuerdo al método desarrollado por Schulte et al. (2003).
Pigmentos fotosintéticos
Las hojas utilizadas para las mediciones del intercambio gaseoso se utilizaron para la extracción y determinación de los pigmentos fotosintéticos (clorofila y carotenoides) y protectores (compuestos que absorben a 305 nm). Los pigmentos fotosintéticos se extrajeron a partir de 3 discos (2,89 mm de diámetro) obtenidos de la parte media de la semilámina foliar con un sacabocados metálico. Los discos se transfirieron a tubos Eppendorf conteniendo 2 ml de dimetil sulfóxido (DMSO) y se mantuvieron a 45ºC en oscuridad (Chappelle et al., 1992). El contenido de clorofila a, clorofila b, clorofila total y carotenoides se determinó a partir de las lecturas de absorbancia a 649, 665 y 480 nm contra un blanco de reactivos en un espectrofotómetro UV-visible (Hitachi U-2800, Japan) usando las ecuaciones de Wellburn (1994):
Clorofila a (μg ml-1) = (12,19 x A665) – (3,45 x A649)
Clorofila b (μg ml-1) = (21,99 x A649) – (5,32 x A665)
Clorofila total (μg ml-1) = Cl a + Cl b
Carotenoides (μg ml-1) = (1000 x A480 – 2,14 x Cl a – 70,16 x Cl b)/ 220
El contenido de pigmentos fotosintéticos se expresó en mg g-1 PS (peso seco). Para la determinación de los pigmentos absorbentes de UV-B se utilizaron discos foliares de 3 mm de diámetro obtenidos de las mismas hojas utilizadas para cuantificar los pigmentos fotosintéticos. Para la extracción de los pigmentos absorbentes los discos (2 por muestra) se colocaron en tubos conteniendo 2 ml de metanol: ácido clorhídrico: agua (79:1:20); luego de esto, los tubos se incubaron en estufa a 45ºC durante 12 h (Mireki y Teramura, 1984). Completada la incubación se removieron los discos y se leyó la absorbancia a 305 nm contra un blanco de reactivos en un espectrofotómetro UV-visible (Hitachi U-2800, Japan). Los resultados se expresaron como Abs305 g-1 PS.
Análisis estadístico
Los resultados obtenidos fueron sometidos primero a un test de Shapiro para estudiar su distribución y luego una prueba F (Fisher) para conocer la homegeneidad o heterogeneidad de las varianzas. Posteriormente se aplicó un análisis de la varianza (ANOVA) utilizando el paquete estadístico Infostat (versión 1.1) (Universidad Nacional de Córdoba) para establecer la existencia o no de diferencias significativas entre los tratamientos (p < 0,05 y p <0,01).
Resultados
Respuesta de la asimilación fotosintética neta (AJ a la densidad de fujo fotosintético de fotones (DFFF). Curva (A /DFFF)
El comportamiento de la A frente a incrementos en la DFFF (curva A /DFFF) resultó diferente en los ensayos "Densidad alta" y "Densidad baja" (Fig. 1). El incremento de la A en función de la DFFF fue prácticamente el mismo en ambos tratamientos hasta aproximadamente 500 μmol m2 s1. A partir de este punto se observó un incremento significativo en la A y un desplazamiento también significativo del punto de saturación lumínica (PSL) en el tratamiento de "Baja densidad" en comparación con las de "Alta densidad". Sin embargo, el punto de compensación lumínica (PCL), la eficiencia cuántica de la fotosíntesis (ØCO2) y la respiración nocturna (Rn) no presentaron diferencias estadísticamente significativas entre los tratamientos (Tabla 1). Estos resultados ponen claramente de manifesto cuán importante resulta la elección de la densidad de siembra para obtener un óptimo rendimiento fotosintético de las plantas y por ende un mayor rendimiento en biomasa y/o granos.
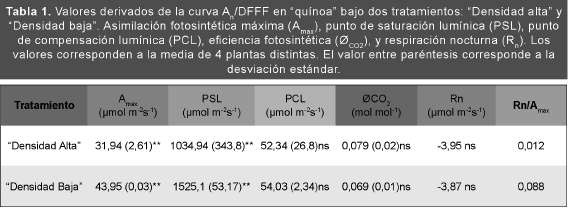
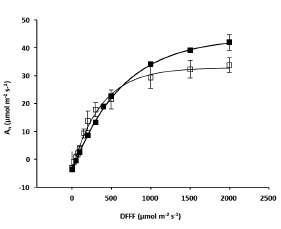
Fig. 1. Respuesta de la asimilación fotosintética neta (An) a valores crecientes de densidad de fujo fotosintética de fotones (DFFF) en "Densidad baja" (■) y "Densidad alta" (□). Cada valor corresponde a la media de 5 plantas distintas con 4 repeticiones por planta. La barra representa la desviación estándar.
Asimilación fotosintética máxima (A )max
Ambos tratamientos presentaron diferencias significativas (p ≤ 0,05) en la A de CO2. Esta max resultó mayor en las plantas que se desarrollaron a "Densidad baja" de siembra (Tabla 1). La diferencia entre ambos tratamientos fue del 27,3%. Como se discute más adelante este resultado tendría una relación directa tanto con la contenido de N foliar, concentración de pigmentos fotosintéticos y anatomía de la hoja.
Conductancia estomática (gs), concentración interna de CO2 (Ci), transpiración estomática (Tr), eficiencia intrínseca en el uso del agua (EUAi) y capacidad de carboxilación (Amax/Ci)
Los valores promedios de gs, Tr y Amax/Ci resultaron significativamente diferentes entre ambos tratamientos (Tabla 2) donde las plantas cultivadas a "Densidad baja" mostraron los valores más altos. El valor de la conductancia estomática (gs) osciló entre 0,10 y 0,20 mol m-2 s-1 para el tratamiento "Densidad alta" y entre 0,18 y 0,29 mol m-2 s-1 para el caso de "Densidad baja". La transpiración estomática (Tr) exhibió valores entre 4,36 y 7,07 mmol m-2 s-1 para "Densidad alta" y entre 6,48 y 8,76 mmol m-2 s-1 para "Densidad baja". La capacidad de carboxilación (An/Ci), un indicador de la productividad fotosintética, mostró variaciones máximas y mínimas de 124,6 y 150,9 mmol m-2 s-1 para "Densidad alta" y 188,3 y 223,6 mmol m-2 s-1 para las correspondientes al ensayo "Densidad baja".
Las variaciones no significativas, entre ambos tratamientos, se observaron en los parámetros Ci y EUAi (Tabla 2). Algunos autores han reportado como rasgo común de las plantas de quinoa, sometidas a estrés hídrico o salino, el aumento de la EUAi (Jacobsen et al., 2009; Eisa et al., 2012). Este hecho podría estar relacionado con la escasa variación de la EUAi observada entre los tratamientos "Densidad alta" y "Densidad baja" debido a la ausencia de estrés hídrico entre ambos ya que los mismos se mantuvieron bajo riego por goteo durante todo el cultivo, aunque esta hipótesis debería ser comprobada con otras experiencias a campo y laboratorio que exceden el presente estudio.
Correlaciones An/gs, Tr/gs y EUAi/gs
Las curvas que relacionan la tasa de fotosíntesis neta (An) con la conductancia estomática (gs) muestran que para valores altos de gs, la asimilación de CO2 resulta menos afectada en las plantas del tratamiento de "Baja densidad" (Fig. 2). Para un mismo valor de gs (por ej. 0,2 mol m-2 s-1) las plantas en el tratamiento "Densidad baja" muestran un valor de An mayor, lo que sugiere que además de limitaciones estomáticas habría otras de naturaleza metabólica en aquellas correspondientes al tratamiento "Densidad alta". Por otro lado, de los datos resulta que a una gs de 0,25 la respuesta fotosintética satura, es decir la conductancia estomática ya no limita la entrada de CO2 y empezarían a actuar otro/s factor/es sobre la An. La variación de Tr en función de la gs, (Fig. 3) mostró para ambos tratamientos un comportamiento no lineal que responde a una función cuadrática. Aunque la curva representativa de la función cuadrática fue más estilizada en el ensayo "Densidad alta", los valores de r de las ecuaciones correspondientes a ambas densidades de siembra fueron bastantes similares. La relación entre EUAi y gs se muestra en la Fig. 4. Para ambas densidades de siembra la curva EUAi/gs responde a una función polinómica cuadrática. Teniendo en cuenta que las plantas en ambos tratamientos no presentan problemas de estrés hídrico o limitaciones de agua durante el desarrollo del cultivo, ya que se mantuvo el riego, a partir de las curvas (EUAi/gs) obtenidas se puede deducir que para valores altos de gs el valor de EUAi es bajo, pero crece a medida que gs decrece.
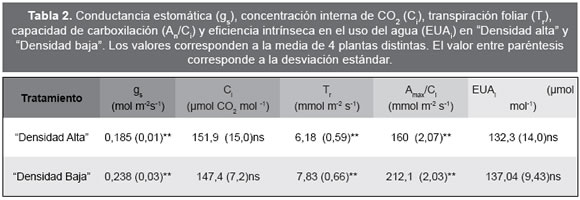
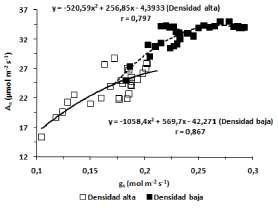
Fig. 2. Correlación entre An y gs en los tratamientos "Densidad baja" (■) y "Densidad alta" (d).

Fig. 3. Correlación entre Tr y gs en los tratamientos “Densidad baja” (■) y “Densidad alta” (d).
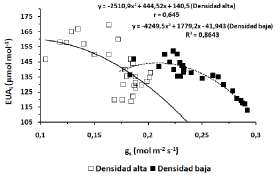
Fig. 4. Correlación entre EUAi y gs en los tratamientos Densidad baja” (■) y “Densidad alta” (d).
Contenido de pigmentos fotosintéticos y compuestos protectores de UV-B
Con excepción de la clorofla b, los pigmentos fotosintéticos (clorofila total, clorofla a, relación Cl a/Cl b y carotenoides) y protectores de UV-B fueron más elevados en las plantas "Densidad alta" (Tabla 3). Sin embargo, el análisis estadístico no mostró diferencias significativas respecto a los valores correspondientes al ensayo "Densidad baja". Considerando que las hojas utilizadas para las determinaciones de los pigmentos se colectaron de la parte superior del follaje, donde la incidencia de la radiación solar para ambos tipos de plantas es la misma, surge el interrogante si la densidad de siembra “ per se” afecta la síntesis y/o acumulación de pigmentos fotosintéticos y protectores de UV-B en quinoa.

Discusión y Conclusiones
Los datos existentes sobre el comportamiento de la "quinoa" en respuesta al incremento de la radiación (DFFF) son escasos. Más escasos aún son los estudios en esta especie relacionados con el efecto de diferentes densidades de siembra sobre los parámetros fotosintéticos. Al momento existen datos, derivados de otras experiencias, con objetivos diferentes (Eisa et al, 2012), donde se informa que trabajando con "quinoa" de la variedad peruana Hualhuas se encontraron valores de 0,156 mol mol1 y -4,57 μmol m2 s1 para ØCO2 y R , respectivamente. Por su parte, los valores de PSL y PCL informados para esta misma variedad fueron 1.149 y 28,4 μmol m2 s1, respectivamente. A excepción de la ØCO2, los valores PSL, PCL, R reportados para la variedad Hualhuas fueron similares a los que se encontraron en este estudio para el tratamiento "Densidad baja".
La diferencia observada en los valores de A max obtenido entre los tratamientos "Densidad alta" con "Densidad baja", se correlacionan con el espesor de la lamina foliar e incluso con el espesor del tejido en empalizada como ya fuera informado en un estudio complementario (Erazzú et al., 2016). Se conoce que las plantas cuyas hojas se hallan más expuestas a la radiación solar presentan una asimilación fotosintética más elevada, una mayor densidad estomática y un mayor espesor de epidermis, mesóflo esponjoso y sistemas vasculares) (Evans y Poorter, 2001). En consonancia con estos datos las hojas de las plantas del tratamiento "Densidad baja", presentaron un mayor espesor de lámina foliar, mayor desarrollo del tejido en empalizada y mayor densidad estomática (ver Erazzú et al., 2016). Resultados ecofsiológicos similares fueron reportados para otras variedades de quinoa (González et al., 2010; 2014) y para otras especies cultivadas (Evans, 1989). En relación al contenido de clorofilas y su relación con la asimilación fotosintética se ha demostrado, para otras especies, que existe una correlación positiva entre ambas variables (Morgan & Lecain, 1991). Sin embargo, en nuestro estudio esa relación no se manifesta ya que los contenidos de clorofilas totales y las isoformas a y b, no presentaron diferencias significativas entre ambos tratamientos. Esto podría explicarse por el hecho que las hojas donde se han medido las mismas, independiente de la densidad de siembra que se trate, se hallaban expuestas al mismo régimen luminoso (hojas totalmente expuestas durante el día en ambas densidades de siembra). Este último hecho es corroborado por el contenido de carotenoides que resulta ser similar en ambos casos. Se conoce que estos compuestos están relacionados con la protección del aparato fotosintético y como tal pueden incrementarse frente a posibles fotooxidaciones causada por una mayor cantidad de radiación solar (Lichtenthaler, 2007; Nisar et al., 2015). De manera que la asimilación fotosintética mayor en el tratamiento "Densidad baja" no estaría correlacionada con el contenido de cloroflas sino con otros factores como el contenido de N foliar y disposición de los tejidos en las hojas, entre otros. En efecto, se conoce que la asimilación fotosintética está fuertemente correlacionada con el contenido de nitrógeno foliar (N) (Field & Mooney, 1986; Evans, 1989) y que incluso éste puede ser usado como un predictor de la asimilación fotosintética de los cultivos cuando la intensidad de la radiación solar es elevada (Vincent, 2001). En consonancia con esta hipótesis, en un informe previo (Erazzú et al., 2016) se dio a conocer que las plantas bajo el tratamiento "Densidad baja" exhibieron un contenido de N foliar más elevado (103,1 mmol N/m2) que en aquellas que se hallaban cultivadas en "Densidad alta" (79 mmol N/m2). Relacionando estos últimos datos con los de Amax obtenidos en este estudio, surge que ambas variables sí se hallan correlacionadas positivamente. Este hecho tendría relación con lo que se conoce que un elevado porcentaje de N foliar se encuentra formando parte de las enzimas, muchas de las cuales intervienen en la fotosíntesis y más concretamente en el Ciclo de Calvin, tal el caso de la Ribulosa 1,5 Bisfosfato Carboxilasa Oxigenasa (RUBISCO) que representa el 50% de toda la proteína presente en la hoja (Lambers et al., 2008). De este modo, podemos concluir que los valores incrementados de asimilación fotosintética neta en el tratamiento "Densidad baja" se correlacionan directamente con el contenido de N foliar medido en esa condición de densidad, más las adaptaciones morfológicas y anatómicas de las hojas.
Otro punto relacionado con la disminución de la An en el tratamiento "Densidad alta" con respecto al tratamiento "Densidad baja", podría estar en conexión con la disminución de la luz al atravesar el follaje según lo postulado por Flénet et al. (1996). De acuerdo a Munns (2002), este efecto se traduciría en una reducción en el crecimiento de las plantas y un menor rendimiento en granos. Los datos de quinoa, que se informan en este estudio más los complementarios ya mencionado (Erazzú et al., 2016), la altura de las plantas, diámetro de tallo, área foliar específica (AFE) y rendimiento granario son menores en el tratamiento "Densidad alta", lo que coincidiría con la hipótesis planteada por Munns (2002). Los datos disponibles al momento indicarían que el mayor valor de Amax lograda en el tratamiento "Densidad baja" tendría su correlato con el tamaño del grano logrado. En efecto, los resultados informados por Erazzú et al. (2016) mostraron que las diferencias entre "Densidad baja" y "Densidad alta" de siembra de "quinoa" no sólo se manifestaron en un mayor rendimiento granario por planta sino también en un mayor tamaño de los granos. Así, granos con diámetro ≥1,68 mm se encontraron en mayor porcentaje en el tratamiento "Densidad baja" (23%) frente al 12% que aparece en la siembra de "Densidad alta". Sin embargo, es necesario profundizar este tema con otras mediciones a lo largo de los diferentes estratos de hojas, para arribar a una conclusión más certera.
En relación al cálculo de la eficiencia intrínseca en el uso del agua (EUAi) en nuestro estudio no se han detectado diferencias significativas entre los tratamientos. En ambos casos, sí se han detectado valores altos de EUAi, coincidentes con otros valores informados para "quinoa" (Razzaghi et al., 2015; Eisa et al. 2012). Algunos autores han postulado que la EUAi podría ser utilizada como criterio para la selección de variedades destinadas a ambientes donde el agua no es abundante (Medrano et al., 2007). Sin embargo, llegar a una conclusión de este tipo, para el caso de quinoa, requiere sin duda de otras evaluaciones de campo, con diferentes variedades y con diferentes manejos agriculturales. Los datos obtenidos para EUAi en nuestro estudio se obtuvieron a partir del cociente entre An y gs, (como se menciona en Materiales y Métodos) para eliminar el efecto de la temperatura y el gradiente de humedad sobre la transpiración (Rawson et al., 1977). Este cálculo se deberá tener en cuenta en futuras comparaciones ya que existe otra forma de cálculo que involucra a la transpiración en vez de gs.
Los resultados de este estudio más otro complementario (Erazzú et al., 2016), demuestran la interrelación de parámetros morfológicos y fsiológicos con la densidad de siembra utilizada y una correlación no sólo con el rendimiento granario sino con el tamaño de los granos. Sin duda, para un cultivo nuevo en los valles intermontanos del Noroeste Argentino, como es el caso de la "quinoa", resulta necesario profundizar estos estudios referidos a la densidad de siembra en las variedades que se están ensayando (nativas e introducidas) no sólo para un mejor aprovechamiento de la misma sino también para conocer los mecanismos fsiológicos que esta especie despliega en ambientes que normalmente se consideran marginales.
Agradecimientos
A la Fundación Miguel Lillo por el apoyo en equipos de campo, personal y laboratorios. Al personal del Campo Demostrativo de Encalilla (Amaicha del Valle, Tucumán) administrado por el Instituto Nacional de Tecnología Agropecuaria (INTA-Famaillá) por su asistencia diaria para el mantenimiento de las parcelas.
BiblIografia
CHAPPELLE, E.W., M.S. KIM & J.E. MCMURTREY III. 1992. Ratio analysis of reflectance spectra (RARS): an algorithm for the remote estimation of the concentrations of chlorophyll a, chlorophyll b, and carotenoids in soybean leaves. Remote Sens. Environ. 39: 239-247. [ Links ]
EISA, S., S. HUSSIN, N. GEISSLER & H.W. KOYRO. 2012. Effect of NaCl salinity on water relations, photosynthesis and chemical composition of quinoa (Chenopodium quinoa Willd.) as a potential cash crop halophyte. Aust. J. Crop Sci. 6: 357-368. [ Links ]
ERAZZÚ, L.E., J.A. GONZÁLEZ, S.E. BUEDO & F.E. PRADO. 2016. Efectos de la densidad de siembra sobre Chenopodium quinoa Willd (quinoa). Incidencia sobre variables morfológicas y rendimiento de grano en la variedad CICA cultivada en Amaicha del Valle (Tucumán - Argentina). Lilloa 53: 12-22. [ Links ]
EVANS, J.R. 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78: 9-19. [ Links ]
EVANS, J.R. & H. POORTER. 2001. Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 24: 755-768. [ Links ]
FIELD, C. & H. MOONEY. 1986. The photosynthesis-nitrogen relationship in wild plants. In: GIVNISH, T.J. (ed.). On the Economy of Form and Function. pp. 25-55. Cambridge University Press, Cambridge, England.
FLÉNET, F., J.R. KINIRY, J.E. BOARD, M.E. WESTGATE & D.C. REICOSKY. 1996. Row spacing effects on light extinction coefficients of corn, sorghum, soybean, and sunfower. Agron. J. 88: 185-190.
GEISSLER, N., S. HUSSIN, M. EL-FAR & H.W. KOYRO. 2015. Elevated atmospheric CO2 concentration leads to different salt resistance mechanisms in a C3 (Chenopodium quinoa) and a C4 (Atriplex nummularia) halophyte. Environ. Exp. Bot. 118, 67-77.
GONZÁLEZ, J.A., A. ROLDÁN, M. GALLARDO, T. ESCUDERO & F.E. PRADO. 1989. Quantitative determinations of chemical compounds with nutritional value from Inca Crops: Chenopodium quinoa ("quinoa"). Plant Foods Hum. Nutr. 39: 331-337
GONZÁLEZ, J.A., S. EISA, S. HUSSIN & F.E. PRADO. 2015. Quinoa: an Incan Crop to Face Global Changes in Agriculture. In: MURPHY, K.S., MATANGUIHAN, J. (Eds.). Quinoa: Improvement and Sustainable Production. pp. 1-18. Wiley-Blackwell, Hoboken, NJ, USA.
GONZÁLEZ, J.A., M. BRUNO, M. VALOY & F.E. PRADO. 2010. Genotypic variation of gas exchange parameters and leaf stable carbon and nitrogen isotopes in ten quinoa cultivars grown under drought. J. Agron. Crop Sci. 197: 81-93.
GONZÁLEZ, J.A., G.I. PONESSA, S.E. BUEDO, M.I. MERCADO & F.E. PRADO. 2014. Asimilación fotosintética máxima en variedades de quinoa (Chenopodium quinoa Willd) de diferentes orígenes geográficos y su relación con la morfología foliar. Lilloa 51: 177-193.
GONZÁLEZ, J.A., G.O. MARTÍN (H), M.A. BRUNO & F.E. PRADO. 2016. La "quinoa" (Chenopodium quinoa) como alternativa forrajera en la zona de los Valles Calchaquíes (Noroeste Argentino). Lilloa 53: 74-81.
JACOBSEN, S.E., F. LIU & C.R. JENSEN. 2009. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd.). Sci. Hortic. 122: 281-287.
LAMBERS, H., F.S. CHAPIN & T.L. PONS. 2008. Plant Physiological Ecology. 2º edition. Springer-Verlag, New York, USA.
LICHTENTHALER H. K. 2007. Biosynthesis, accumulation and emission of carotenoids, α-tocopherol, plastoquinone and isoprene in leaves under high photosynthetic irradiance. Photosynth Res 92: 163-179.
MEDRANO, H., J. BOTA, J. CIFRE, J. FLEXAS, M. RIBAS-CARBÓ & J. GULÍAS. 2007. Eficiencia en el uso del agua por las plantas. Invest. Geogr. 43: 63-84.
MILTHORPE, F.L. & J. MOORBY. 1982. Introducción a la Fisiología de los Cultivos. Hemisferio Sur S.A, Argentina.
MINETTI, J.L. 2005. El clima del Noroeste Argentino. Laboratorio Climátológico Sudamericano. Editorial Magna.
MIREKI, R. & A.H. TERAMURA. 1984. Effects of Ultraviolet-B irradiance on soyben. V. the dependence of plants sensitivity on the photosynthetic photon fux density during and afiter leaf expansion. Plant Physiol. 74: 475-480.
MORGAN, J.A. & D.R. LECAIN. 1991. Leaf gas exchange and related leaf traits among 15 winter wheat genotypes. Crop Science 31: 443-448.
MUNNS, R. 2002. Comparative physiology of salt and water stress. Plant Cell. Environ. 25: 239-250.
NISAR, N., L. Li, S. LU, N.C. KHIN & B.J. POGSON. 2015. Carotenoid metabolism in plants. Molecular Plant 8: 68-82.
QUIROGA, C., R. ESCALERA, G. ARONI, A. BONIFACIO, J.A. GONZÁLEZ, M. VILLCA, R. SARAVIA & A. RUÍZ. 2014. Traditional processes and technological innovations in quinoa harvesting, processing and industrialization. In: BAZILE D. (Ed.), State of the art report on quinoa around the world in 2013 pp. 258-296. FAO (Santiago de Chile) y CIRAD, (Montpellier, Francia).
RAWSON, H.M., J.E. BEGG & R.G. WOODWARD. 1977. The effect of atmospheric humidity on photosynthesis, transpiration and water use efficiency of leaves of several plant species. Planta 134: 5-10.
RAZZAGHI, F., S.E. JACOBSEN, C.R. JENSEN & M.N. ANDERSEN. 2015. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought - mechanisms of tolerance. Funct. Plant Biol. 42: 136-148.
SCHULTE, M., C. OFFER & U. HANSEN. 2003. Induction of CO2-gas exchange and electron transport: comparison of dynamic and steady-state responses in Fagus sylvatica leaves. Trees 17: 153-163.
VINCENT, G. 2001. Leaf photosynthetic capacity and nitrogen content adjustment to canopy openness in tropical forest tree seedlings. J. Trop. Ecol. 17: 495-509.
WELLBURN, A.R. 1994. The spectral determination of clorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 144: 307-313.
Recibido el 22 de junio de 2017, aceptado el 7 de septiembre de 2017. Editor: Federico Mollard.

