











 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducáo
Os sistemas de polinizagao das angiospermas tem sido interpretados sob diferentes pontos de vista em relagao á evolugao das interagoes entre plantas e polinizadores e o grau de especializagao- generalizagao (Faegri & van der Pjil, 1979; Waser et al., 1996; Fenster et al., 2004; Ollerton et al., 2007; Rosas-Guerreiro et al., 2014; Armbruster, 2017; Martins et al., 2022). Em virtude das diferengas morfológicas, fisiológicas e comportamentais entre visitantes florais é esperado que as pressoes seletivas que estes exercem sobre os atributos florais de certa espécie sejam distintas, prevalecendo no sistema os polinizadores mais efetivos. Essa ideia é a base do “princípio do polinizador mais efetivo” (Stebbins, 1970) e encontra correspondencia no conceito de síndromes de polinizagao (Faegri & van der Pjil, 1979), que implicitamente assume evolugao em diregao a sistemas de polinizagao especializados (Ávila Jr. & Freitas, 2011). Contudo, muitas espécies sao polinizadas por uma ampla gama de espécies de visitantes, pertencentes a diferentes grupos taxonómicos, ou seja, constituem sistemas generalistas (Waser et al., 1996). O predomínio de sistemas de polinizagao especialistas ou generalistas nas comunidades e a validade do conceito de síndromes estiveram na base dos principais debates na literatura internacional neste campo de estudo nas últimas décadas (e.g. Rosas- Guerreiro et al., 2014; Armbruster, 2017; Dellinger, 2020). Apesar das divergencias e lacunas, existe uma razoável concordancia em torno de que as interagoes estao distribuidas em um gradiente entre extremos de especializagao e generalizagao e que existem diferentes formas de se definir os sistemas de polinizagao como especializados e generalistas (Fenster et al., 2004; Ollerton et al., 2007; Armbruster, 2017).
Muitas espécies arbóreas nas florestas tropicais, com flores pequenas a inconspícuas, coloragao pálida e com néctar em pequena quantidade e facilmente acessível, tem sido associadas a um sistema generalista de polinizagao por diversos pequenos insetos (Wardhaugh, 2015), ou sistema de polinizagao por diversos insetos (DI) (sensu Moreira & Freitas, 2020). Apesar de ter sido proposto há quase quatro décadas para espécies de árvores de uma comunidade de floresta tropical na Costa Rica (Bawa et al., 1985; Bawa, 1990), foram realizados muito poucos estudos de biologia da polinizagao de plantas com atributos florais do sistema DI (Moreira & Freitas, 2020). Plantas classificadas neste sistema sao visitadas por insetos de diversos grupos (e.g., abelhas, besouros, borboletas, moscas e vespas) e tem características florais semelhantes ás plantas classificadas em um sistema de polinizagao por abelhas pequenas, principalmente abelhas sem ferrao, proposto por Bawa & Opler (1975) para árvores tropicais com sistema sexual dióico. A diferenciagao entre esses dois sistemas de polinizagao (i.e., DI e abelhas pequenas) tem como base a frequencia de visitas dos polinizadores (Bawa et al., 1985; Bawa, 1990), embora seja difícil se reconhecer os respectivos fenótipos florais, além de dificuldades para delimitagao conceitual dos dois sistemas (Moreira & Freitas, 2020). Na verdade, o conjunto de espécies apontadas como polinizadas por diversos insetos pode de fato nao constituir um único sistema de polinizagao e sim uma gama ampla de sistemas generalistas (q.v., Martins et al., 2022).
Abelhas sem ferrao (Apidae: Meliponini) constituem uma parcela importante dos visitantes florais em florestas tropicais, porém sua importancia na polinizagao tem sido questionada (Janzen, 1975; Inouye et al., 1980; Renner & Feil, 1993; Murphy & Breed, 2008). Embora espécies de Meliponini sao capazes de percorrer distancias em torno de 2 km em busca de alimento (Roubik & Aluja, 1983), essa capacidade nao reflete necessariamente as distancias usuais de forrageio, influenciada por outras variáveis, tais como localizagao e abundancia de recursos alimentares, formas de orientagao e marcagao de trilhas de forrageamento e disponibilidade de locais de nidificagao (Araujo et al., 2004). Abelhas sem ferrao tendem a permanecer um longo tempo forrageando em apenas um indivíduo e por isso tem sido consideradas ineficientes quanto á movimentagao intraespecífica de pólen, o que caracterizarla as interagoes como antagonistas (Janzen, 1975). Além disso, tem sido registradas obtendo recursos florais de modo inapropriado (pilhadores e ladroes de néctar e pólen, sensu Freitas, 2018), em flores com tamanho e morfologia adequadas para polinizagao por outros grupos (Murphy & Breed, 2008; Alves-dos-Santos et al., 2016). Em contraste, outros trabalhos tem apontado que estas abelhas desempenhariam papel importante na polinizagao do dossel de florestas tropicais, mesmo que promovendo mais autogamia e geitonogamia de que polinizado cruzada (Bawa, 1977, 1980, 1990; Momose et al., 1998; Ramalho, 2004; Pires & Freitas, 2008; Monteiro & Ramalho, 2010).
Sapindaceae possui cerca de 150 géneros e 2000 espécies distribuidas em regioes tropicais e subtropicais, em sua maioria com flores díclinas (Somner et al., 2015). Cupania é um dos maiores géneros na família, com espécies que possuem inflorescéncias paniculadas terminais, com flores actinomorfas de colorado branca a amarelada e tamanho reduzido (Somner et al., 2009). As flores femininas e masculinas apresentam, respectivamente, estaminódios e pistilódios bem desenvolvidos, dificultando a distingao dos morfos feminino e masculino (Stevens 2001). Espécies do genero tém sido indicadas como tendo sistemas de polinizado generalistas, devido ao fenótipo floral e ao registro de diversos insetos como visitantes florais, embora tenha sido registrada polinizado principalmente por abelhas nas duas espécies neotropicais estudadas em detalhe (Bawa 1977; Ferreira 2009). Neste trabalho estudamos a biologia floral, sistema reprodutivo, sucesso reprodutivo e a composigao dos visitantes florais de Cupania oblongifolia Mart. Os dados foram obtidos com o intuito de caracterizar o sistema de polinizagao da espécie e explorar aspectos da morfologia floral e da biologia reprodutiva que se relacionam aos visitantes florais.
Material e Métodos
Area de estudo e espécie estudada
O trabalho foi realizado no Parque Nacional de Itatiaia (PNI) (em torno de 22° 27’44’’S; 44° 36’15’’W). O PNI está localizado a oeste do estado do Rio de Janeiro e ao sul de Minas Gerais, no Macigo de Itatiaia, um dos principais remanescentes de Mata Atlántica da Serra da Mantiqueira. Com uma área de aproximadamente 30.000 ha, a vegetagao do PNI na faixa de 500-1500 m amsl (acima do nivel médio do mar) de altitude é descrita como Floresta Ombrófila Densa Montana. O clima do Itatiaia é dividido em seis diferentes níveis altitudinais, sendo as cotas de 500 a 1000 m amsl caracterizadas, segundo a classificagao de Koeppen, como Cwa e Cfb com periodos frios apresentando baixa pluviosidade e periodos quentes com alta pluviosidade (Segadas-Vianna & Dau, 1965).
Cupania oblongifolia é uma espécie arbórea heliófita de até 20 m de altura (Somner et al., 2009), que ocorre predominantemente em formagoes florestais em estágios iniciais de sucessao secundária (Lorenzi, 1998). É endémica do territorio brasileiro, sendo encontrada nas regioes Norte, Nordeste, Sul e Sudeste, nos dominios da Floresta Amazónica, Cerrado e Mata Atlántica (Somner et al., 2015). Foi registrada na faixa altitudinal de 500 a 800 m amsl na floresta montana no PNI e em áreas limitrofes adjacentes. Em estudo sobre a composigao floristica e a estrutura de vegetagao na floresta montana do PNI com árvores de DAP > 10 cm, esta espécie apresentou 7 individuos ha-1 (Guedes-Bruni, 1998), embora sua abundancia nas florestas do Itatiaia seja maior em áreas expostas a efeitos de borda (A.T. Fonseca, obs. pess.). O fruto é do tipo cápsula, trigono-piriforme apresentando trés sementes com arilo alaranjado (Somner et al., 2009). Material testemunho foi depositado no herbário do JBRJ (A.T. Fonseca, 05 - RB707038 e A.T. Fonseca, 06 - RB707040).
Biologia floral e polinizadores
A biologia floral foi estudada em campo e no laboratorio, a partir de material coletado e armazenado em FAA 70%. A presenga de osmóforos foi verificada por meio do teste do vermelho neutro (Dafni et al., 2005). A viabilidade polinica foi estimada através do corante carmim acético (Dafni et al., 2005), em graos de pólen de anteras de flores estaminadas e pistiladas. A receptividade estigmática foi testada em campo pela reagao com peróxido de hidrogénio a 3% (adaptado de Kearns & Inouye, 1995). Os testes foram feitos em diferentes fases ao longo da vida das flores, sendo associados á turgidez e coloragao do estigma para posterior avaliagao de duragao da antese. Para determinar a longevidade e os recursos florais, flores pistiladas e estaminadas foram marcadas e ensacadas no inicio da antese e observadas até a senescéncia. Devido ao porte da espécie, andaimes de construgao foram montados para acesso ás flores.
Os polinizadores foram estudados durante o dia, através de observagao direta e registradas por meio de cámera digital (Dafni et al., 2005). O comportamento de forrageio foi registrado em flores pistiladas e estaminadas e os visitantes que entraram em contato com os órgaos reprodutivos das flores dos dois morfos foram considerados polinizadores potenciáis. A composigao dos visitantes foi determinada primeiramente por meio de observagoes aleatórias e amostragens- piloto em diferentes plantas e horarios do dia, totalizando 14 horas. Para registro da frequencia dos visitantes florais foram realizadas sessoes de coleta focal. Foram marcados oito individuos em pontos próximos a trilhas do Parque. Nas sessoes de coleta focal, todos os visitantes de cinco ramos florais por individuo foram coletados com pugá. Foram realizadas oito sessoes de coleta focal para cada morfo floral de 30 min cada, entre 10:00h e 14:00h, em dias sem chuva ao longo dos meses de abril, maio e junho. As espécies de visitantes florais coletados foram identificadas em laboratório com auxilio de literatura e consulta a especialistas. A dominancia das espécies foi calculada para ambos os morfos pela fórmula D% = (i/t) x 100, sendo i o total de individuos de uma espécie e t o total de individuos coletados. A dominancia foi categorizada em eudominante (D > 10%), dominante (5% < D < 10%), subdominante (2% < D < 5%), recessiva (1% < D < 2%) e rara (< 1%) (Friebe, 1983 apud Ferreira, 2009). Os visitantes florais coletados em ambos os morfos florais e com frequencia superior a 5% foram observados sob microscópio estereoscópico para verificar a presenga e localizagao de graos de pólen (cabega, tórax e pernas). Foi construida uma curva de acumulagao de espécies (Legendre & Legendre, 1998) a partir do número de individuos coletados por espécie, a qual nao alcangou a assintota, indicando maior riqueza de espécies de visitantes florais do que o amostrado.
Sistema Reprodutivo
Para determinar o sistema reprodutivo foram realizados testes controlados de polinizagao manual. Foram selecionados quatro individuos nos quais inflorescencias com botoes foram isoladas com sacos de “voile”. Os cruzamentos foram realizados no primeiro dia de receptividade estigmática. Todos os tratamentos foram aplicados em cada um dos quatro individuos amostrados. As flores foram submetidas aos seguintes tratamentos: 1. flores nao-manipuladas, flores pistiladas foram previamente ensacadas e deixadas intactas para aferir a formagao de frutos sem fertilizagao (i.e., teste para apomixia); 2. autopolinizagao manual (pólen de flores masculinas), flores pistiladas foram polinizadas com o pólen de flores estaminadas do mesmo individuo (i.e., geitonogamia); 3. autopolinizagao (pólen de flores femininas), flores pistiladas foram polinizadas com o pólen retirado dos estaminódios de flores pistiladas do mesmo individuo; 4. polinizagao cruzada (pólen de flores masculinas), flores pistiladas receberam pólen de flores masculinas de outro individuo a pelo menos 50 m de distancia; 5. Polinizagao cruzada (pólen de flores femininas), flores pistiladas receberam pólen retirado dos estaminódios de flores pistiladas de outro individuo, a pelo menos 50 m de distancia; e 6. Polinizagao natural, flores nao-ensacadas foram marcadas e acompanhadas para observagao da polinizagao em condigoes naturais. O sucesso de polinizagao em cada tratamento foi estimado peloa “fruit set” em cada tratamento, por meio da contagem de frutos, aproximadamente cinco semanas após a antese. Os frutos estavam bem desenvolvidos após este periodo, mas ainda imaturos, o que foi feito com intuito de evitar efeito da intensa predagao de sementes por larvas de lepidópteros na estimativa de fecundidade. A diferenga na frutificagao dos tratamentos foi analisada através do teste Qui- quadrado (x2) (Zar, 1999). A produgao de frutos após polinizagao cruzada manual e em condigoes naturais foi comparada para cálculo do indice de limitagao polinica (ILP = 1 - (Fc / Fpc), sendo Fc a quantidade de frutos produzidos sob condigoes naturais e Fpc a quantidade de frutos produzidos sob polinizagao cruzada manual (Larson & Barrett, 2000).
Resultados
Durante o periodo de estudo, C. oblongifolia floresceu de abril a julho e frutificou de agosto a outubro. A espécie apresentou dinámica de floragao alternada num mesmo individuo, com inflorescencias bissexuadas contendo flores pistiladas separadas temporalmente das flores estaminadas, com pouca ou nenhuma sobreposigao entre as fases sexuais (i.e., dicogamia sincronizada). A quantidade de botoes por inflorescencia e de inflorescencias por ramos florais foi de 357,6 ± 284,9 (n = 20) e 3,5 ± 1,4 (n = 26), respectivamente (média ± d.p., aqui em todo o texto). As flores sao pentameras e possuem tamanho pequeño, com 3-4 mm de diámetro e 4-6 mm de comprimento. As flores estaminadas apresentam um pistilo nao funcional (i.e., pistilódio) com tres óvulos vestigiais e um gineceu reduzido circundado por oito estames com anteras de coloragao variando de alvacenta a vinácea e deiscencia rimosa (Figura 1A). As flores pistiladas apresentam um pistilo bem desenvolvido com ovário contendo tres óvulos de placentagao axial (Figura 1B). O ovário é circundado por oito estaminódios, de mesma coloragao que nas flores estaminadas, os quais produziram pólen, porém as anteras permaneceram fechadas durante toda antese. Alta porcentagem de graos de pólen de ambos os morfos coraram com carmim acético, especificamente 97,8 ± 0,29% (n = 3) nas flores estaminadas e 98,4 ± 0,90% (n = 3) nas pistiladas. Em ambos os morfos, um nectário extrastaminal em forma de anel ocorre na base das flores, circundando os estames ou os estaminódios. As flores apresentaram um cheiro adocicado durante toda a antese. Os filetes e as anteras dos estames e estaminódios, o nectário e as pétalas de ambos os morfos reagiram ao teste de vermelho neutro, indicando a ocorrencia de osmóforos. A antese das flores estaminadas durou em torno de seis dias, com o erguimento gradual de um a tres estames por dia, seguido da abertura das anteras (n = 15 flores, 3 plantas). Nao foi observado um horário específico de inicio da antese. Após a senescencia das flores estaminadas, as flores pistiladas iniciavam a antese, com o erguimento completo do estigma em dois a quatro dias, permanecendo receptivas por cerca de tres dias, quando ocorre a mudanga de coloragao e perda de turgidez do estigma (n = 15, 3 plantas). A produgao de néctar em ambos os morfos nao foi medida, mas sua presenga foi constatada pela presenga de gotículas ou filme brilhante sobre o nectário e inferida pela observagao de insetos sorvendo a secregao.
Cupania oblongifolia depende de polinizadores pois nao é apomítica, mas é autocompatível, frutificando após cruzamento autogámicos e geitonogamicos (Tabela 1), embora isso é limitado naturalmente pela dicogamia sincronizada. A frutificagao nao diferiu entre os tratamentos, exceto pelo sucesso menor após autopolinizagao manual realizada com pólen dos estaminódios das flores femininas (Tabela 1, x2 = 23,06; p = 0,05; n = 496 e 5). Apesar desta diferenga, houve frutificagao após cruzamentos de autopolinizagao e de polinizagao cruzada utilizando pólen retirado dos estaminódios (Tabela 1).
Foram coletados 438 individuos de visitantes florais pertencentes a 87 morfoespécies de insetos de quatro ordens (Tabela 2, Figuras 2, 3). Maioria das visitas foi em flores masculinas, com 302 individuos (68,8%) de 62 espécies, enquanto que nas flores femininas foram 136 individuos (31,2%) de 45 espécies. Os morfos florais compartilharam 20 espécies (22,9%), sendo 13 de Hymenoptera, quatro de Diptera e tres de Lepidoptera. Espécies de Hymenoptera tiveram maior abundancia, com 79% do total de individuos coletados. Espécies de Diptera e Lepidoptera apresentaram abundancias semelhantes, com 11% e 10% dos individuos, respectivamente (Tabela 2). A tribo Meliponini respondeu por mais de 70% dos individuos de espécies de Hymenoptera (Figura 4), sendo Scaptotrigona bipunctata (Figura 4A, B) a única espécie eudominante nas flores femininas e masculinas. Outra espécie que desempenhou papel importante na polinizagao foi Scaptotrigona sp.1, apresentando eudomináncia em flores masculinas e domináncia em flores femininas. Apis mellifera (Figura 4C, D) apresentou subdominancia para os dois morfos florais. Já Paratrigona subnuda (Hymenoptera: Meliponini) (Figura 4E, F) e Urbanus dorantes dorantes (Lepidoptera: Hesperiidae) tiveram dominancia nas flores masculinas e femininas, respectivamente, embora tenham sido apenas recessiva e rara, respectivamente, nos morfos florais opostos. O restante dos visitantes florais foi considerado recessivo ou raro, com excegao de Ornidia obesa (Figura 3B), a qual apresentou dominancia em flores femininas.

Fig. 1: Diferentes morfos florais de Cupania oblongifolia. A. Flor feminina (pistilada) apresentando pistilo desenvolvido e oito estaminódios com filetes reduzidos e anteras parcialmente encobertas pelos verticilos florais. B. Flor masculina (estaminada) apresentando estames desenvolvidos e um pistilódio com estilete pouco desenvolvido. C. Corte longitudinal da flor feminina mostrando o ovário e os óvulos desenvolvidos, e os estaminódios ao redor do ovário circundados pelo nectário. D. Corte longitudinal da flor masculina mostrando o ovário e óvulos pouco desenvolvidos, e os estames ao redor do ovário circundado pelo nectário.
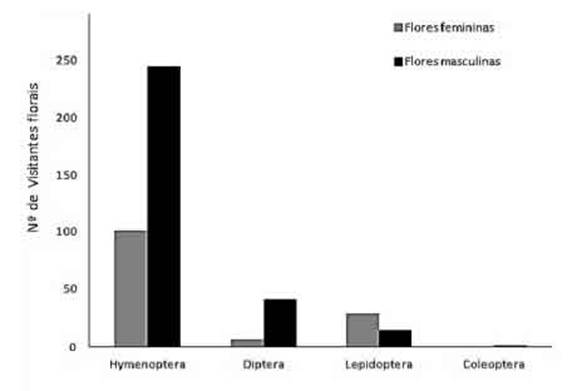
Fig. 2: Frequência acumulada de indivíduos de visitantes florais de espécies de Hymenoptera, Diptera, Lepidoptera e Coleoptera, coletados em flores masculinas e femininas de Cupania oblongifolia, na Mata Atlântica montana do Itatiaia, sudeste do Brasil.

Fig. 3: Exemplo de visitantes florais das diferentes ordens de insetos registradas em flores masculinas de Cupania oblongifolia na Mata Atlântica montana do Itatiaia, sudeste do Brasil. A: Hymenoptera: Melipona cf. rufiventris (Meliponini, Apidae). B: Diptera: Ornidia obesa (Syrphidae). C: Lepidoptera: Placidula euryanassa D: Coleoptera: Cantharidae sp.

Tabela 1: Produção de frutos de Cupania oblongifolia após diferentes tratamentos de polinização na Mata Atlântica montana do Itatiaia, sudeste do Brasil. Letras diferentes indicam diferenças significativas (p < 0,05) após comparação por teste Qui-quadrado (χ²).

Tabela 2: Frequências absolutas de visitantes florais de Cupania oblongifolia, coletados em flores femininas e masculinas, de abril a junho de 2011, na Mata Atlântica montana no Itatiaia, sudeste do Brasil. Abreviaturas: Ni = número de indivíduos, Dom = categoria de dominância, Eu = eudominante, D = dominante, Sd = subdominante, R = recessiva, Rr = rara; N = néctar; P = pólen; ? = incerto.
Os visitantes comegavam as visitas ás flores por volta das 7:00h, com maior frequencia e riqueza de visitantes entre 10:00h e 14:00h. A partir deste horário, as visitas comegavam a declinar, até nao serem mais observadas por volta das 17:30h. Apenas espécies de Hymenoptera foram observadas buscando tanto pólen como néctar como recurso floral. Nestes casos, a coleta de pólen ocorreu preferencialmente na parte da manha junto com a coleta do néctar, enquanto na parte da tarde, néctar foi o principal recurso explorado. A unidade de polinizagao foi a flor, com os insetos pousando demais meliponíneos analisados sob microscópio estereoscópico, todos os individuos amostrados tinham presenta de pólen de C. oblongifolia, ao menos na cabega. Somente 12% dos individuos de U. dorantes (n = 8) apresentaram pólen aderido ao corpo, sendo apenas nas pernas e em pequena quantidade em comparagao com as abelhas.

Fig. 4: Abelhas sociais coletando néctar em flores de Cupania oblongifolia na Mata Atlântica montana do Itatiaia, sudeste do Brasil. O principal polinizador da espécie, Scaptotrigona bipunctata em flor feminina (A) e masculina (B). Note pólen depositado na cabeça. A espécie exótica Apis mellifera, um polinizador de importância secundária, visitando flores femininas (C) e masculinas (D). Paratrigona subnuda, cuja importância na polinização é reduzida, pois visitam raramente as flores femininas (E), apesar de muito frequentes em flores masculinas (F). Apesar do tamanho reduzido, durante as visitas caminham sobre as flores e tocam o estigma ou as anteras com as pernas e porção ventral do corpo.
Discussáo
O sistema de polinizagao de C. oblongifolia abarca aspectos de generalizagao, mas pode ser enquadrado como um caso de especializagao em grupo funcional (sensu Ollerton et al., 2007; Armbruster 2017). Isso porque, embora uma alta riqueza de espécies de polinizadores de diferentes ordens tenha sido registrada nas flores desta espécie, abelhas Meliponini foram responsáveis pela maioria das visitas em ambos os morfos. Mais que isso, foi possivel identificar uma espécie como polinizador principal, já que Scaptotrigona bipunctata respondeu por quase metade da frequencia de visitas de polinizadores em flores femininas. Esses resultados se assemelham aos de outros sistemas de polinizagao em que ocorre especializagao funcional em situagao de generalizagao ecológica (Ollerton et al., 2007; Narbona & Dirzo, 2010; Niemirski & Zych, 2011; Bartos et al., 2015; Martins & Freitas 2018).
Elevada frequencia abelhas Meliponini foi observada em outras espécies de Cupania (Bawa 1977; Ferreira 2009). Em C. guatemalensis (Turcz.) Radlk. na América Central, os polinizadores observados foram predominantemente espécies de Trigona (Bawa 1977), enquanto que em C. vernalis Cambess. no sul do Brasil, o principal polinizador foi S. bipunctata (Ferreira 2009), tal como em C. oblongifolia no Itatiaia (ver também Ramalho, 1990 para C. cinerea Poepp. & Endl.). Variagoes espago- temporais na fauna antófila sao um componente importante dos sistemas de polinizagao, em particular nas flores com fenótipo floral generalista. Entretanto, os resultados dos estudos realizados em Cupania indicam que a polinizagao por abelhas sem ferrao é uma característica disseminada no genero.
Apesar da elevada frequencia de visitas a flores, abelhas sem ferrao supostamente tem pouca movimentagao entre plantas coespecificas e por isso tem sido consideradas como promotoras de autogamia e geitonogamia, conferindo pouco ou nenhuma polinizagao cruzada (Janzen 1975; Ramalho, 2004; Monteiro & Ramalho 2010). Isso seria critico para a reprodugao sexual de uma espécie como C. oblongifolia, que requer transferencia de pólen entre individuos, apesar da autocompatibilidade. Porém, observamos a presenga de pólen na maioria das abelhas sem ferrao que coletamos nas flores femininas. Além disso, nao encontramos diferengas na frutificagao após cruzamentos cruzados e em condigoes naturais (i.e., ausencia de limitagao polinica), indicando que os polinizadores realizaram movimento de pólen entre coespecificos, em niveis no minimo no limiar de capacidade de produgao de frutos da espécie. A efetividade dos polinizadores pode ser otimizada pela alta densidade de individuos de C. oblongifolia na área de estudo (Fonseca & Freitas, dados nao publ.). Uma explicagao alternativa seria contribuigao do vento na polinizagao (e.g. Zamudio et al, 2021), o que nao foi avaliado.
O papel das abelhas sem ferrao na polinizagao é um tema ambivalente. Por um lado, tendem a forragear próximo ao ninho, recrutam outros individuos com grande eficiencia, umedecem cargas de pólen com néctar para adesao na corbicola e podem danificar partes florais em busca de recursos (exemplos em Roubik ,1989; Thorp, 2000; Nieh, 2004; León et al., 2015; Leonhardt, 2017). Entretanto, o comportamento de forrageio de abelhas sem ferrao é muito variável, dependendo de características da espécie e das suas interagoes com outras abelhas, bem como da comunidade de plantas e de atributos da paisagem (Biesmeijer & Slaa, 2004). A fecundidade de C. oblongifolia no Itatiaia indica que a polinizagao por abelhas sem ferrao foi efetiva. Isso pode refletir a combinagao de dois aspectos, primeiro a intensidade de floragao de C. oblongifolia, já que floragao massiva foi associada a maior chance de polinizagao por abelhas sem ferrao (Ramalho, 2004).
Além disso, as flores pistiladas de C. oblongifolia ocorrem separadas temporalmente das flores estaminadas num mesmo individuo ao longo da floragao, ou seja, ocorrencia de dicogamia sincronizada (Lloyd & Webb, 1986; ou dioicia funcional, Cruden, 1988). Floragao em sequencia sincronizada da qual ocorre pouca ou nenhuma sobreposigao entre as fases pistilada e estaminada é conhecida para várias espécies de Sapindaceae (Cruden, 1988; Lima et al., 2016), inclusive em Cupania (Bawa, 1977; Ferreira, 2009). Essa alternancia entre as fungoes sexuais da planta impede ou pelo menos minimiza as chances de autogamia e geitonogamia, enquanto aumenta as chances da polinizagao cruzada (Bawa, 1977; Cruden, 1988). Assim, a floragao massiva acarretaria em alta visitagao e a dicogamia sincronizada favoreceria o fluxo de pólen entre individuos de C. oblongifolia. Em suma, nossos resultados se somam a outros estudos que indicam que Meliponini constitui importante grupo de polinizadores de espécies arbóreas na regiao Neotropical (Roubik, 1989; Pires & Freitas, 2008; Moreira & Freitas, 2020; Bueno et al., 2021).
A ausencia de sistema de incompatibilidade era esperada em C. oblongifolia. Existe uma estreita associagao entre dicogamia sincronizada e autocompatibilidade, com poucas excegóes (Cruden, 1988). A dicogamia sincronizada promove a polinizagao cruzada impedindo ou pelo menos diminuindo as chances de ocorrerem autogamia ou geitonogamia (Bawa, 1977; Cruden, 1988). As autopolinizagoes manuais com pólen retirado dos estaminódios (i.e. de anteras fechadas) das flores femininas apresentaram formagao de frutos, indicando sua viabilidade. Em Cupania guianensis Miq. (=Cupania scrobiculata Rich.) os graos de pólen retirados de estaminódios também se apresentaram viáveis quando corados, embora tratamentos de polinizagao manual nao tenham sido realizados (Bawa, 1977). A produgao de pólen nos estaminódios das flores femininas de Cupania é difícil de explicar em uma perspectiva de custo energético, podendo refletir peculiaridades da ontogenia floral, tais como efeitos de pleiotropia.
O fato dos estaminódios das flores femininas de C. oblongifolia nao sofrerem deiscencia, mesmo possuindo graos de pólen viáveis, pode refletir pressóes seletivas contra a autofertilizagao e para atratividade das flores femininas. Essas características das flores de C. oblongifolia sugerem haver mimetismo de flores masculinas por flores femininas, o que tem sido apontado na literatura como responsável pela atragao e aumento da frequencia nas flores femininas de agentes polinizadores em busca dos recursos oferecidos pelas flores masculinas (e.g., Baker 1976; Ágren et al., 1986; Castillo et al., 2012; Yadav et al., 2020). Tal mimetismo é amplamente distribuido em plantas monoicas e dioicas e é apontado como resultante de competigao intraespecífica por polinizadores (Dafni, 1984).
Conclusáo
Nossos resultados se somam a outros estudos, os quais tem mostrado que espécies com flores pequenas de simetría radial e néctar acessivel, usualmente apontadas como generalistas associadas a insetos de diversos grupos, podem na verdade apresentar sistemas de polinizagao com certo grau de especializagao. Abordagens que descrevam a efetividade dos polinizadores permitem melhor compreensao acerca destes sistemas de polinizagao e, embora nao seja o ideal, medidas de frequencia já fornecem um avango em relagao á riqueza de espécies de visitantes isoladamente (Martins et al., 2022). O agrupamento de espécies em um sistema de polinizagao por diversos insetos (sensu Bawa, 1990) nao diferencia possiveis graus de especializagao em certas espécies de polinizadores ou mesmo em um grupo funcional (q.v., Moreira & Freitas, 2020). Neste sentido, uma questao fundamental para o entendimento da ecologia e evolugao dos sistemas de polinizagao, particularmente em florestas tropicais, é se a polinizagao por diversos insetos representa um sistema convergente, ou seja, uma unidade definida nas comunidades, ou se trata de diversos conjuntos de atributos florais que determinam sistemas de polinizagao distintos, tanto em termos do grau de generalizagao, como dos principais grupos de polinizadores.
Agradecimentos
A Maria Cristina Gaglianone (UENF) pelo auxilio na identificagao das abelhas; Marcia Couri (MN-UFRJ) pela identificagao das moscas; a Pedro Acevedo-Rodríguez (Smithsonian Institute) e Genise Somner (UFRRJ) pela confirmagao da identificagao de C. oblongifolia e discussoes acerca do genero; a Anna Karla da Venda pelo auxilio nas coletas de campo; a Carolina Saad pela ajuda nas análises laboratoriais; Alexandre Christo (in memoriam) pela ajuda nas análises estatísticas; aos proprietários do sítio Rancho Fundo pela permissao de estudo e receptividade; ao ICMBio e á equipe do PNI pela autorizando de pesquisa e apoio logístico ao estudo; ao Projeto Biodiversidade do Bioma Mata Atlántica - PROBIO II/MCT/JBRJ pelo apoio financeiro parcial; á CAPES pela concessao de bolsa de mestrado a ATF; ao CNPq e FAPERJ pelas bolsas de Produtividade em Pesquisa e Cientista do Nosso Estado, respectivamente, a LF. Este trabalho faz parte da Dissertagao de Mestrado de ATF, apresentada ao Programa de Pós-Graduagao em Botánica do Jardim Botánico do Rio de Janeiro.

