











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
A lo largo de su historia, la humanidad ha transportado voluntaria o involuntariamente miles de especies más allá de sus áreas naturales de distribución (Zalba et al., 2008; Castro-Díez et al., 2005). No obstante, con el aumento demográfico este fenómeno se ha multiplicado de manera exponencial, en cuanto al tipo y cantidad de especies transportadas y el rango geográfico que pueden alcanzar (Cassey et al., 2018), constituyendo hoy en día el avance de especies exóticas y los impactos de este proceso, una preocupación creciente en el contexto de la conservación mundial (Freitas & Pivello, 2002). Las alteraciones que las especies invasoras causan en los ecosistemas pueden ser múltiples y complejas (Mack & D’Antonio, 1998; William & Baruch, 2000; Vilá & Hulme, 2017), y no solo a nivel ecológico sino también económico y social (Morandi et al., 2020; Duboscq-Carra et al., 2021).
Para revertir o minimizar esta situación, desde hace tiempo diferentes entes públicos y privados plantean políticas y estrategias de planes de monitoreo y control de las especies exóticas invasoras (Schüttler & Karez, 2009; Simberloff et al., 2013). La erradicación o control de las especies invasoras es otra de las alternativas, aunque con tratamientos que no comprometan la existencia de la vegetación nativa, o no causen efectos inesperados. Por último, será necesario poner en marcha acciones que permitan iniciar un proceso de recuperación de áreas degradadas y elegir un método para la restauración de especies nativas adecuadas a las características de cada región (Rossi et al., 2014).
En la provincia de Misiones se pueden encontrar gramíneas (Poaceae) exóticas de los géneros Cenchrus L., Cynodon Rich., Megathyrsus (Pilg.) B.K. Simon & S.W.L. Jacobs, Melinis P. Beauv., Phyllostachys Siebold & Zucc., Urochloa P. Beauv. Varios de estos géneros poseen especies que son consideradas invasoras en la Argentina y el mundo.
Una de las áreas de la provincia más amenazadas por el avance de gramíneas invasoras corresponde a sabanas arboladas sobre arenisca del departamento de San Ignacio, conocidos como los campos del Teyú Cuaré y alrededores. Este ambiente de superficie restringida (ca. 70 ha), resguarda cerca del 50% de las especies de plantas vasculares de Misiones. Sumado a esto en dicha área existe un alto nivel de endemismo con al menos ocho especies de plantas vasculares microendémicas, algunas en peligro crítico de extinción, con poblaciones restringidas en superficie o constituidas por unos pocos ejemplares (Farinaccio & Keller, 2014; Keller, 2015; Keller & Crockett, 2015; Keller & Tressens, 2016; Deble et al., 2017). Asimismo, estos pastizales se consideran los únicos fragmentos de Cerrado presentes en la Argentina (Velazco et al., 2018a) y cuentan con sectores que están siendo ocupados por poblaciones de Melinis minutiflora P. Beauv.
Esta especie originaria de África central y del sur, y fue introducida en zonas tropicales de América (Salariato, 2012). En Brasil su introducción se da alrededor del siglo XVII posiblemente de forma involuntaria por semillas traídas en las camas hechas en paja de los navíos de la época (Dutra Silva et al., 2015). Ya en el siglo XVIII se cita siendo usada en campos de pastoreo próximos al estado de Rio de Janeiro (Rossi et al., 2010). Y en el siglo XIX el pasto gordura o capim gordura como se la conoce vulgarmente, es uno de los primeros pastos exóticos citados por los viajeros europeos al describir los paisajes del interior del Cerrado en Brasil (Dutra Silva et al., 2015). En Misiones fue citada por Martinez Crovetto (1977), como nueva especie para la flora argentina. Este autor menciona que es utilizada como protectora del suelo en yerbales donde se encuentra naturalizada y adventicia en suelos modificados. Posiblemente la causa de la introducción al país se haya dado por su utilidad como forrajera.
La invasión por parte de esta especie en las áreas mencionadas con anterioridad puede determinar eventualmente la extinción de especies o su reubicación a categorías de vulnerabilidad cada vez más críticas. Sin embargo, cualquier plan de acción que se decida tomar sobre esta situación debe contar con estudios previos de la estructura y composición de la vegetación; como así también ensayar métodos de control que permitan seleccionar las opciones de manejo más adecuadas para cada situación. En consideración a todo esto, el objetivo del presente trabajo es presentar un estudio sobre el avance de M. minutiflora y evaluar su impacto sobre la vegetación nativa en pastizales de Teyú Cuaré.
Materiales y métodos
Area de estudio
El área bajo estudio se ubica en el departamento de San Ignacio, Misiones (Argentina), próxima al parque provincial del Teyú Cuaré (Fig. 1). Abarca aproximadamente 70 ha de pastizales arbolados cuya principal expresión arbórea se limita a ejemplares más o menos dispersos de Acosmium subelegans (Mohlenbr.) Yakovlev (Fabaceae). Solo una parte mínima de esta área se encuentra bajo protección desde el año 2020 como reserva natural privada.
Los campos de Teyú Cuaré se sitúan sobre un paisaje ondulado que geomorfológicamente responde a la continuación de la Sierra de Amambay de Paraguay (Soria, 1996). El suelo se corresponde con el “Udipsamente típico” (Ligier et al., 1990) que se caracteriza por ser bien drenado, fuertemente ácido, con muy baja dotación de nutrientes. Este tipo de suelo, llamado Areniscas de Misiones, sólo se encuentra en la Argentina en los alrededores de San Ignacio y se ubica sobre afloramientos de sedimentos continentales formados en el Mesozoico bajo condiciones de aridez (Teruggi, 1970).
El clima es subtropical sin estación seca. Las precipitaciones son de aproximadamente 1700 mm anuales en la zona, y el promedio anual de temperatura es 21 °C (Soria, 1996).
Se ubica fitogeográficamente en la Provincia Paranaense (Cabrera, 1976), en la unión de los distritos Fluvial, del Urunday y de los Laureles (Martínez Crovetto, 1963), aunque a nivel predial recientemente se ha demostrado su filiación con el Bioma Cerrado (Velazco et al., 2018 a, b).
Diseño experimental
Se utilizó un muestreo aleatorio que consistió en 50 parcelas de 1 m2, subdivididas en 100 cuadrículas de 10 cm por 10 cm (Fig. 2A). De estas cuadrículas, 25 fueron distribuidas en áreas afectadas por M. minutiflora (CMm) y 25 en áreas sin presencia de esta especie invasora (SMm), totalizando 50 m2 de muestreo. La distancia entre las parcelas fue con una separación mínima de 20 m. Dentro de cada parcela el relevamiento de la información consistió en identificar cada individuo, registrar su nombre científico y contabilizar el número de cuadrículas que cada individuo abarcó como medida estimativa de su cobertura. Nosotros utilizamos el término individuos para plantas que eran visiblemente identificables, para el caso de las gramíneas hace referencia a la estructura del macollo o matas. Para las gramíneas con hábitos más estolonífero u otros, que no se pudo distinguir los macollos se procedió únicamente a contabilizar las cuadriculas que abarcaba la especie y se consideraba como un único individuo.
Análisis de datos
Fitosociológicos: Los datos para el análisis florístico fueron relevados en julio de 2015. Las plantas fueron identificadas a nivel de especie y su nombre científico fue corroborado según el Catálogo de las Plantas Vasculares del Cono Sur (Zuloaga et al., 2019) y su actualización online (www.darwin.edu.ar).
Los parámetros fitosociológicos calculados para cada especie fueron Frecuencia absoluta Fa=(Pe/Ni)*100, donde Ni es el número total de muestras, Pe es el número de parcelas en la que estuvo presente una determinada especie. La frecuencia relativa Fr=(Fai/Fat)*100 donde la Fai es la frecuencia absoluta de la especie i y Fat es la frecuencia absoluta de todas las especies. La cobertura relativa Cr=(Ci/Ct)*100, donde Ci es la cobertura total de la especie i (cantidad de cuadrículas en la que se encuentra presente una especie) y Ct es la cobertura total para todas las especies. Por último, se determinó el valor de importancia para cada especie según la ecuación VI=Fr + Cr. Este valor define cuáles de las especies presentes contribuyen en el carácter y estructura al ambiente.
Evaluamos el esfuerzo de muestreo mediante dos criterios, el primero consistió en la comparación entre la curva de rarefacción individual y los estimadores de riqueza no paramétricos Chao 2 y ACE. Para este criterio, se logra una muestra satisfactoria cuando los valores de los estimadores de riqueza son aproximadamente iguales al valor extremo obtenido en la curva de acumulación de especies. Según Alvarez et al. (2004) si el número de especies encontradas es un 85% o más del total de las especies estimadas, se considera un muestreo satisfactorio. Para el segundo caso se llevó a cabo una curva de extrapolación, donde según Ellenberg & Mueller-Dombois (1974) un muestreo puede ser considerado idóneo cuando el incremento del 10% del esfuerzo de muestreo no implica un incremento de la riqueza de especie superior al 10%. Se empleó el software EstimateS versión 9.1.0 (Colwell, 2013) para generar las curvas de acumulación, extrapolación y de los estimadores no paramétricos Chao 2 y ACE. Las tres últimas curvas fueron construidas en base a 100 aleatorizaciones, utilizando una matriz de presencia/ausencia de especies de las 25 parcelas no invadidas por M. minutiflora. Esto debido a que, si tomamos en cuenta solo las áreas invadidas el esfuerzo de muestreo se debería incrementar para lograr la suficiencia, invirtiendo más recursos y tiempo de trabajo. De lo contrario si tomamos las dos áreas como base podríamos estar realizando un submuestreo de las áreas no invadidas.

Fig. 1: Ubicación general del área de estudio. A: Noreste de Argentina, con el recuadro indicando el área que se observa en B. B: Sur de Misiones, el recuadro indica el sur del departamento San Ignacio. C: Imagen satelital del área de estudio, próxima a Teyú Cuaré.

Fig. 2: Parcelas del muestreo y ambientes donde habita M. minutiflora. A: Parcelas de 1 m2 subdividida cada 10 cm. B-C: M. minutiflora en bordes de caminos. D-E: Poblaciones de M. minutiflora (manchas rojizas) en pastizales naturales. Abreviaturas= Mn: Melinis minutiflora; Mnf: Melinis minutiflora en floración.
Diversidad: Comparamos los sitios invadidos con los sitios no invadidos utilizando los perfiles de diversidad de Chao & Jost (2015). Estos perfiles de diversidad calculan diferentes valores de diversidad cambiando gradualmente el factor q, que determina la sensibilidad de la medida a las abundancias relativas de las especies. Es decir, calcula diferentes índices de diversidad para diferentes pesos de abundancia de especies, lo que permite comparar las comunidades utilizando toda la información presente en los datos (Jost, 2019). El valor q igual a 0 es la riqueza específica, por ende, cuenta las especies por igual sin tener en cuenta sus abundancias relativas. Para q igual a 1 es el exponencial del índice de Shannon-Wiener que cuenta a los individuos por igual, por lo tanto, pesa las especies en proporción a sus abundancias. Y para un valor de q igual a 2 es el valor inverso al de Simpson que descuenta todas las especies menos las dominantes y puede interpretarse como el número efectivo de especies dominantes (Hsieh et al., 2014; Chao & Jost, 2015). Con lo cual en lugar de seleccionar una o algunas medidas para describir una comunidad, es preferible transmitir la historia completa presentando un perfil continuo, es decir, un gráfico de diversidad en función de q > 0, lo que facilita la comparación visual de la composición y complejidad de múltiples comunidades, y permite juzgar la uniformidad de las distribuciones de abundancia relativa (Chao et al., 2014; Jost, 2019).
Tanto el análisis de diversidad y los gráficos se realizaron en el software R v.3.4.1 (R Core Team, 2017), se utilizó los paquetes SpadeR (Chao et al., 2016) para obtener los valores de los perfiles de diversidad, ggplot2 y ggtheme para gráficos.
Superficie: La determinación de la superficie afectada por M minutiflora se realizó en el mes de julio de los años 2015, 2016 y 2017 en dos etapas. La primera consistió en la marcación del perímetro de las áreas afectadas y de focos emergentes por medio de un GPS en el campo. La segunda etapa se llevó a cabo en gabinete y tomando como base las marcaciones en campo se realizó la cartografía pertinente y los cálculos de la superficie de invasión a través del software libre QGIS (QGIS Development Team, 2016).
Resultados
Suficiencia muestral
La comparación de las curvas para evaluar el esfuerzo muestral se puede observar en laFig. 3. Como resultado para el primer criterio utilizado según el estimador no paramétrico Chao 2 se esperó encontrar unas 95 especies, y para el estimador ACE unas 98 especies. Sin embargo, el número de especies que se encontró fue 81, lográndose una completitud de 85 y 83% correspondientemente.
Para el segundo criterio se extrapoló el número de parcelas a 30 unidades que representó un aumento del 20% del total de parcelas, esto implicó encontrar unas 85 especies. Este aumento representa aproximadamente un 5% del número total de especies, siendo como máximo el 10% de incremento para que el muestreo sea considerado como suficiente.
Fitosociológicos
Se relevaron un total de 96 especies distribuidas en 26 familias en toda el área muestreada. Analizando por separado en los lugares SMm se encontraron un total de 81 especies distribuidas en 25 familias, siendo las tres familias más importantes Asteraceae, Poaceae y Fabaceae con 14, 11 y nueve especies respectivamente (Fig. 4A). En cambio, a nivel de individuos puede observarse en la Fig. 4B que en primer lugar se ubican las Poaceae seguidas por las Asteraceae y en tercer lugar las Fabaceae.
Por otro lado, en los lugares CMm se encontraron un total de 45 especies distribuidas en 17 familias, donde las familias más representativas a nivel de especies son Asteraceae con 13, Poaceae con seis y Fabaceae con cinco. A nivel de individuo las familias con más individuos fueron asimismo Asteraceae, Poaceae y Fabaceae (Fig. 4C-D).
Para los lugares SMm (Tabla 1) puede observarse que la especie con mayor importancia fitosociológica esAxonopus suffultus (J.C. Mikan ex Trin.) Parodi (Poaceae) con el 14 %, seguida de Elionurus muticus (Spreng.) Kuntze (Poaceae) con el 11,03%, ambas gramíneas muy frecuentes en lomadas arenosas de Misiones y Corrientes. Para el caso de la muestra en áreas CMm (Tabla 2) el valor de importancia porcentual más alto corresponde a la especie invasora con más del 46%, mientras que A. suffultus pasa al segundo lugar con apenas el 5,78% y E. muticus se reduce al 0,52%, ocupando el puesto 31. En tercer lugar, de las áreas afectadas aparece Schwenkia americana D. Royen ex L. (Solanaceae), con el 4,11%, se trata de una hierba grácil, con comportamiento ruderal, capaz de adaptarse a la espesura de la cobertura graminosa densa. En estas tablas se aprecia asimismo el impacto sobre la uniformidad del valor de importancia, siendo la variación de este índice para las diferentes especies más gradual en áreas SMm, donde el 50% se distribuye en nueve especies nativas, mientras que en áreas CMm, este porcentaje se distribuye tan solo en dos especies, solo una de las cuales es nativa.

Fig. 3: Curvas de acumulación de especies (línea negra) y su intervalo de confianza (95%; área gris), extrapolación (línea roja discontinua), estimadores no paramétricos ACE (línea celeste) y Chao 2 (línea verde).

Fig. 4: A: Distribución del número de especies por familia para las áreas sin invasión. B: Distribución del número de individuos por familia para las áreas sin invasión. C: Distribución del número de especies por familia para las áreas con invasión. D: Distribución del número de individuos por familia para las áreas con invasión.

Tabla 1: Valores de parámetros fitosociológicos de las especies (ordenadas con los valores de VI en forma decreciente) para lugares no afectados por M. minutiflora. Abreviaturas= Ci: Cobertura individual; Cr: Cobertura relativa; Fa: Frecuencia absoluta; Fr: Frecuencia relativa; Pe: Presencia; VI: Valor de importancia; VI%: Valor de importancia porcentual.
Diversidad
El análisis de diversidad abordado a través de los perfiles dio como resultado que la mayor diversidad se encuentra en las áreas que no están invadidas para los tres valores del coeficiente q de mayor relevancia, siendo los valores del índice para las parcelas SMm q0 igual a 81, q1 igual a 41,1 y para q2 igual a 55,5; mientras que para las parcelas CMm los valores correspondientes son q0 igual a 45, q2 igual a 17,4 y q2 igual a 1 (Fig. 5). Por otro lado, en los gráficos de la Fig. 5 se aprecia además que las muestras SMm presentan una curva con pendiente menor, lo que refleja una mayor equidad en la distribución de los individuos por especie. Por el contrario, el perfil de las áreas CMm muestra una curva más pronunciada, pero ubicada muy por debajo de la curva SMm. Estas diferencias se deben a que, por un lado, las áreas CMm presentan mucho menos riqueza (45 especies) y con una distribución de dominancia que favorece a unas pocas especies (baja equidad).
Incremento en superficie
Para la primera medición correspondiente al año 2015 (Fig. 6A) la superficie invadida era de 9.774,5 m2, con un total de 17 parches o manchones. Para el año siguiente, 2016 (Fig. 6B) la superficie total afectada fue de 18.822,6 m2 con 20 parches, con lo cual la superficie se incrementó 9.078,1 m2 en dicho periodo, es decir, con valores cercanos al 100% de incremento. Al segundo año de medición se registraron ocho nuevos focos de proliferación, y 11 de los 17 que estaban presentes en el 2015 se expandieron de madera considerable.
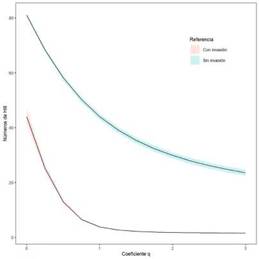
Fig. 5: Perfiles de diversidad para las áreas invadidas (línea inferior rosada) y las áreas sin invasión (línea superior celeste); las áreas sombreadas corresponden a intervalo de confianza del 95%.
Para el año 2017 la superficie afectada fue de 23.396,9 m2, uniéndose algunos de los parches aislados que se encontraban separados en el año 2016. Otro aspecto importante para este último periodo de medición es la gran cantidad de focos nuevos (Fig. 6C). También puede apreciarse que la mayoría de los focos de ocupación por parte de esta especie tienen lugar en bordes de caminos en uso o abandonados (Fig. 2B-C). En la Fig. 6D se muestra un gráfico de la tendencia creciente de la ocupación de M. minutiflora en los pastizales estudiados.
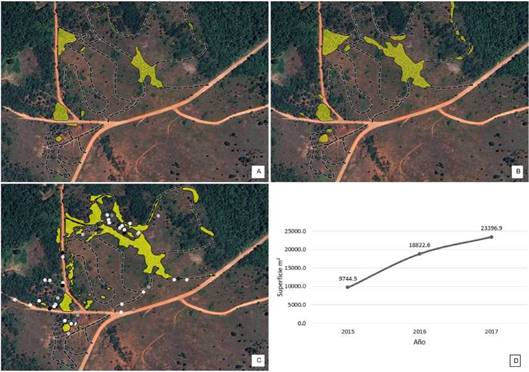
Fig. 6: Áreas invadidas por M. minutiflora (zonas amarillas) en el periodo 2015-2017. A: Año 2015. B: Año 2016. C: Año 2017. D: Línea de tendencia del avance de la superficie de invasión. Los puntos blancos corresponden a pequeños parches de 1 m2, los grises de 4 m2, 8m2 y 10m2, y los negros de 12 m2; las líneas punteadas corresponden a antiguos caminos de circuito de rally.
Discusión y Conclusiones
La superficie invadida en el año 2015 fue de 9.744,5 m2 y para el año 2017 aumentó extendiéndose a 23.396,9 m2, este aumento corresponde a la superficie de focos nuevos de infección y la expansión de los focos ya existentes, lo que en un periodo de tres años totaliza un 240%. Las actividades antropogénicas son la principal causa para que continúe su dispersión, ya que se ha determinado que, como muchas invasoras, se desarrollan mejor en lugares sujetos a disturbios, como a lo largo de caminos, senderos y sitios de deposición de basura, y de ahí avanza hacia el ecosistema circundante, como lo han observado Castillo et al. (2014). En el caso del presente estudio, estas áreas disturbadas corresponden a caminos vigentes o antiguos caminos de circuito de rally (Fig. 2C-E). Este aumento de la superficie de ocupación por sobre la vegetación nativa trae consigo un deterioro sobre el ecosistema en lo que respecta a riqueza, diversidad y distribución de la cobertura. Esto se ve reflejado en los análisis de la diversidad, donde los menores valores para el coeficiente q de los perfiles analizados se encontraron en los lugares afectados por M. minutiflora. Adicionalmente los resultados del presente estudio demuestran que el avance de esta especie exótica, impacta en la distribución de especies por familia, decayendo el número de especies de las familias que presentan mayor riqueza como las Poaceae, Fabaceae, Rubiaceae entre otras, siendo Asteraceae la menos afectada en este sentido. Ello sugiere que el avance de M. minutiflora a nivel local en las áreas que avanza sobre los campos naturales del Teyú Cuaré, se produce una alteración no solo en la diversidad y cobertura, sino también en la composición de la vegetación nativa. Por su parte, esta misma especie adventicia en los cerrados brasileños es considerada una de las principales especies invasoras (Cornwall, 2022) y el principal problema para la conservación de la vegetación en áreas naturales protegidas de este bioma (Pivello et al., 1999b; Martins et al., 2004).
Experiencias realizadas por Hoffmann & Haridasan (2008) en la región del Cerrado de Brasil compararon la germinación de sietes especies de árboles nativos en dos situaciones, un sitio dominado por M. minutiflora y otras por vegetación nativa sin invasión de especies exóticas. La especie adventicia no tuvo ningún efecto discernible sobre la emergencia inicial de plántulas, pero si en el posterior establecimiento, alcanzando valores por debajo de la mitad de la supervivencia con respecto a parcelas dominadas por especies nativas. Estos autores también determinaron que la biomasa aérea de las hierbas de parcelas invadidas era más del doble que el de las parcelas no invadida, mientras que la supervivencia de las plántulas se correlacionó negativamente con la biomasa herbácea, lo que sugiere que la competencia por la luz puede explicar la baja supervivencia de las plántulas donde M. minutiflora es dominante. Esto podría estar explicando las diferencias halladas en este estudio en cuanto al número de individuos y especies por familia comparando las áreas no afectadas y las afectadas por la invasión de esta especie.
El Alto Valor de Importancia de M. minutiflora encontrado en las parcelas afectadas son similares a los hallados por Pivello et al. (1999a) en la Reserva del Cerrado Pé-de-Gigante (Estado de Sao Paulo, Brasil) donde detectaron a las especies M. minutiflora y Urochloa decumbens (Stapf) R.D. Webster, con valores de VI muy altos, y donde ambas hierbas exóticas se asociaron negativamente con la mayoría de las gramíneas nativas, lo que sugiere que ejercen una fuerte presión competitiva en la comunidad herbácea nativa.
Melinis minutiflora aporta una gran cantidad de biomasa al sistema invadido, según Aries (2005) alcanzando hasta un 71% y Martins et al. (2011) un 62% de la biomasa total de las plantas vasculares presentes. Para el caso de los pastizales sobre areniscas en Misiones debe considerarse la gran riqueza de plantas subarbustivas xilopodíferas, que cuentan con una importante biomasa bajo el nivel del suelo, lo cual constituye un reservorio de material reproductivo que puede ser útil a la hora de plantearse iniciativas de restauración.
En virtud de que estas áreas relictuales son únicas para la Argentina y constituyen el nicho ecológico para microendemismos, se hace evidente la imperiosa necesidad de establecer programas para contrarrestar los efectos del avance y advenimiento de invasiones biológicas mediante el estudio previo de técnicas de control apropiadas a cada situación.
Contribución de los Autores
Todos los autores han realizado conjuntamente y a partes iguales la colecta de datos, su análisis e interpretación y redacción del manuscrito.
Agradecimientos
Al Consejo Nacional de Investigación Científica y Técnicas (CONICET). A la Reserva Natural Osununú de la Fundación Temaiken y Club del Río por permitirnos trabajar en parte de su reserva y brindarnos su apoyo.

