










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista argentina de dermatología
versão On-line ISSN 1851-300X
Rev. argent. dermatol. vol.91 no.4 Ciudad Autónoma de Buenos Aires out./dez. 2010
ARTÍCULOS ORIGINALES
Alteraciones biofarmacológicas de la procaína empleada por vía intradérmica
Biopharmacological alterations of procaine used by via intradermal
LC López Agüero * y RJ Dittrich **
* Médica Especialista en Dermatología. Práctica Privada.
** Médico Especialista en Clínica Quirúrgica. Práctica Privada.
e-mail: lucyk@infovia.com.ar
rdittrich@intramed.net
RESUMEN
Se ha demostrado que los anestésicos locales tienen muchos efectos en diferentes células. Actúan directamente a nivel de la membrana plasmática e interactúan con lípidos y proteína de membrana produciendo desorden molecular, inhibición de la actividad de los fibroblastos, induce vacuolización de varios tipos celulares y entre otras acciones, disminuye la síntesis de colágeno.
La procaína inhibe selectivamente el ATP calcio dependiente, bloqueando el ingreso celular del calcio extracelular, altera la función de los microtúbulos y microfilamentos, modifica la forma celular y la distribución de los receptores de membrana, inhibe diversas enzimas como la fosfolipasa A2 implicada en la lipólisis, altera el ADN y el ciclo celular.
A pesar del hecho que los anestésicos locales (Ej: procaína) han sido empleados en fórmulas de mesoterapia, como un "vector indispensable" por más de 55 años, debemos ser cuidadosos en su utilización como un diluyente de fórmulas.
En base a estas modificaciones tisulares, consideramos que es importante conocer las acciones específicas de la procaína en las diferentes estructuras celulares.
Palabras clave: Procaína; Intradermoterapia; Mesoterapia.
SUMMARY
It has been shown that local anesthetics have many effects on different cells. They act directly at the plasma membrane and interact with membrane lipid and protein producing molecular disorder, inhibition of fibroblast activity, induces vacuolization of many cell types and, among other actions, decreases the synthesis of collagen.
Procaine selectively inhibits ATP-calcium dependent, blocking the entry of extracellular calcium to the cell, alters the function of microtubules and microfilaments, changes cell shape and the distribution of membrane receptors, inhibits various enzymes like phospolipase A2 involved in lipolysis, alter the DNA and cell cycle.
Despite the fact that local anesthetics (e.g. procaine) have been used in mesotherapy formulas as an "indispensable vector" for over 55 years, we must be careful in its use as a diluter of formulas.
Based on these tissue changes, we consider it important to know the specific actions of procaine in different cell structures.
KEY WORDS: Procaine; Intradermotherapy; Mesotherapy.
INTRODUCCIÓN
En la actualidad la vía intradérmica se emplea como vector en el tratamiento de diversas patologías, tanto dolorosas como estéticas.
La técnica denominada mesoterapia o intradermoterapia ha empleado la procaína desde hace más de 55 años, como una sustancia elemental e "infaltable" en las fórmulas mesoterápicas.
A principios del siglo 20 la inyección de procaína se utilizó en el tratamiento de ciertas formas de artritis, arteritis, prurito y trombos de los miembros inferiores. Es en los años 50 cuando el Doctor Michael Pistor comienza a emplear la procaína, por vía mesodérmica, para tratamientos locorregionales en diversas patologías, comprobando efectos beneficiosos en trastornos dolorosos (Ej: reumatismo, esguinces, deportología). Esta forma de administración de medicamentos por vía mesodérmica, dio origen a todo un movimiento o escuela denominada "mesoterapia". Con el transcurso de los años, esta vía ha sido empleada en tratamientos estéticos con fármacos extraídos del empleo de cosmecéuticos, muchas veces asociados a la procaína.
Esta larga tradición del empleo de procaína en mesoterapia o intradermoterapia, nos indujo a realizar una revisión de los efectos bio-farmacológicos a nivel tisular del anestésico.
En diversas publicaciones se señalan diferentes acciones de la procaína ejercidas a nivel celular, con especial interés en lo que se refiere a modificaciones de los componentes de la membrana (lípidos y proteínas), tanto celular como de organelas, de los receptores y por último, de la función celular.
El propósito de esta revisión es recordar y actualizar las funciones, bases farmacológicas y el uso clínico que tiene la procaína. También determinar si su empleo en mesoterapia debe ser considerado como un "diluyente" o como una "droga con acción propia" y de empleo restringido a determinadas patologías.
DESARROLLO
Los anestésicos locales son fármacos que, aplicados en concentración suficiente en su lugar de acción, bloquean la conducción de impulsos en tejidos eléctricamente excitables 1. Además de bloquear los impulsos, los anestésicos locales pueden inhibir varios receptores, aumentar la liberación de neurotransmisores (Ej: glutamato) y deprimir la actividad de algunas vías de señalización intracelular 2, entre otras acciones.
Los anestésicos locales actualmente disponibles para el empleo en mesoterapia son dos: los amino-éster (procaína) y los amino-amidas (lidocaína) 2.
Para poder comprender las acciones no tan conocidas de la procaína, se deben mencionar brevemente algunas características farmacológicas básicas 2.
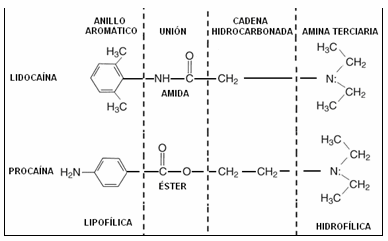
Estructura química: estos Anestésicos Locales (AL) presentan una estructura química superponible, que pueden dividirse en cuatro subunidades (Gráfico I) 1,2,3:
Subunidad 1: anillo aromático. Formado por un anillo benzénico sustituido. Es el principal responsable de la liposolubilidad de la droga. A mayor número de núcleos, mayor liposolubilidad.
Subunidad 2: unión éster o amida. Tipo de unión entre el anillo aromático y la cadena hidrocarbonada. Determina el tipo de degradación de la molécula: los amino-ésteres son metabolizados por las pseudocolinesterasas plasmáticas y los amino-amidas a nivel hepático.
Subunidad 3: cadena hidrocarbonada. Generalmente es un alcohol con dos átomos de carbono. Se relaciona con la liposolubilidad, la duración de acción y la toxicidad de la droga.
Subunidad 4: grupo amina. Determina la hidrosolubilidad de la molécula y su unión a proteínas plasmáticas. Está formado por una amina terciaria (soluble en lípidos) o cuaternaria (soluble en agua).
Gráfico I: estructura química de anestésicos locales. Anillo Aromático = Liposolubilidad. Unión Éster = Degradación Molecular. Cadena Hidrocarbonada = liposolubilidad, acción y toxicidad. Grupo Amina = Hidrosolubilidad, Unión proteínas plasmáticas 1,2,3.
Propiedades físicas: los AL son bases débiles poco solubles en agua y muy solubles en solventes orgánicos, relativamente hidrofóbicos, por lo que deben combinarse con un ácido fuerte (clorhidrato), para obtener una sal estable y soluble en agua a un pH entre 4 a 7. Los AL tipo éster son rápidamente hidrolizados en medios alcalinos, por lo que su preparación tiene un pH menor 2.
La hidrosolubilidad está directamente relacionada con el grado de ionización e inversamente relacionada con la liposolubilidad.
La forma libre ionizada, no es capaz de atravesar las membranas. Es "no liposoluble" o "hidrofílica", está eléctricamente cargada.
La forma no ionizada es "liposoluble", no está eléctricamente cargada y sí atraviesa las membranas.
Cuando la droga es inyectada, la forma liposoluble ("no ionizada") se difunde a través de los lípidos de la membrana celular hacia el interior de la célula, donde al encontrar un pH más bajo se ioniza. Esta molécula ionizada es la forma activa del fármaco al interactuar desde el interior de la célula, ya que no hace efectos cuando se aplica en el exterior de la membrana, pero sí cuando lo hace en el interior.
Al aumentar el pH de la solución aumenta la proporción de fármaco en forma básica, no iónica, mejorando la tasa de difusión a través de la membrana, pero debe tenerse en cuenta que si el pH asciende más allá de 7, se favorece el riesgo de precipitación 2.
Farmacocinética: la absorción de la droga está supeditada a varios factores:
a) Lugar de administración: la absorción depende del grado de vascularización tisular y de la presencia de tejidos, a los que el anestésico local pueda fijarse. La velocidad mediante la que los AL se absorben hacia el torrente sanguíneo y alcanza su máxima concentración sanguínea, varía según la vía de administración (Tabla I).
b) Concentración y dosis.
c) Presencia de vasoconstrictor.
d) Forma unida a proteínas: está unida a la "α1-glicoproteína ácida" y a la "albúmina". Al unirse a proteínas, disminuye la fracción libre de la droga.

Metabolismo: las aminoamidas (lidocaína) presentan degradación enzimática en el hígado. Los aminoéster (procaína) son hidrolizados por esterasas plasmáticas (pseudocolinesterasa) originando dos metabolitos: el PABA (ácido paraaminobenzoico) y el DEAE (dietil-aminoetanol).
La excreción de procaína es por vía renal en un 98% como metabolitos inactivos y sólo un 2% en forma inalterada 2,3.
Los anestésicos locales no solo inducen cambios en la función y estructura de membranas neuronales excitables (bloqueo de conducción nerviosa), sino que también producen cambios en células no neuronales.
La procaína interactúa con la membrana pudiendo fluidificar, expandir, engrosar o derretir los fosfolípidos de membrana 4,5,6,7. Como consecuencia de este proceso, se modifican las proteínas de membrana (estructurales, de canal, de bomba, receptores y enzimas) provocando un cambio conformacional de las proteínas 8 inducido por lípidos.
Estos cambios alteran la modulación de la interacción célula-célula y modifican y/o anulan la función y forma original de la célula 9,10.
Algunas de las alteraciones producidas por la administración de procaína son 10,5: inhibición de la propagación / difusión y adherencia celular, inhibición de la exocitosis y fusión celular, cambios en la fragilidad osmótica 11, inhibición de la acetilcolinesterasa 8, alteración de la endocitosis 4, alteración del metabolismo celular de calcio 12, alteración del citoesqueleto celular 13, modificación de los receptores de membrana 13,14 y entre otras, vacuolización celular 14.
La procaína inhibe selectivamente el ATP calcio dependiente a nivel de membrana, bloqueando el ingreso celular del calcio extracelular 15. A su vez, el anestésico tiene la capacidad de competir por el calcio y desplazar el calcio unido a membrana 13, tanto en proteína como en fosfolípidos 4. Estas modificaciones del calcio producen aumento de la concentración citoplasmática de calcio, a niveles suficientes como para provocar una alteración funcional (despolimerización) y estructural (ruptura) en los microtúbulos y microfilamentos del citoesqueleto, asociados a membrana (submembrana) 13, induciendo disrupción de la organización del citoesqueleto celular 5,10. Por su parte, el anestésico activa la fosfolipasa "C", la que por medio de la modificación del calcio, modula y reduce la concentración de fosfatidilinositol bifosfato, llevando a una disminución de la adhesión del citoesqueleto adherido a membrana 16.
Los microtúbulos y microfilamentos están relacionados con el control transmembrana de la movilidad y distribución de los receptores de membrana 13.
Al estar comprometida la integridad de los microtúbulos y microfilamentos, se modifica la organización normal de membrana, aparecen "blebs" en la superficie celular, hay cambios en la forma celular, se afecta el funcionamiento y distribución normal de los receptores de membrana, se modifica la liberación o secreción (microtúbulos dependiente) de sustancias sintetizadas por células y se altera la adhesión y movilidad celular, así como la endo y exocitosis 5,10,13 (Gráficos II y III).
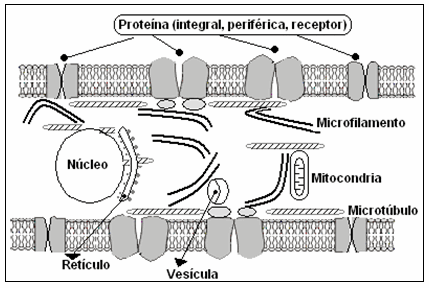
Gráfico II: estructura celular: microtúbulos y microfilamentos intracelulares y de submembrana y su relación con diversas organelas.

Gráfico III: representación esquemática de una célula con formación de blebs de superficie de membrana, alteraciones en los receptores (R) y cambio de su forma luego de ser expuesta a la procaína (MT-MF = microtúbulos-microfilamentos).
Diversos autores 10,17,18,19,20 demostraron que la procaína inhibe la secreción de mucopolisacáridos, colágeno y otras proteínas en forma independiente a la disminución del transporte de aminoácidos o síntesis de proteínas, implicando no solo una disminución del transporte de leucina y prolina, sino a un efecto mecánico sobre la membrana celular. Este efecto mecánico es atribuido, entre otros, a una alteración en la "fijación" del sistema de microtúbulos de la membrana plasmática, que al "desprenderse" de la membrana celular y retraerse, bloquean la secreción de proteínas.
La procaína inhibe la calmodulina, esencial en la regulación de ciertos procesos como 10, 20, 21:
- La bomba Calcio/Magnesio ATPasa.
- La polimerización de los microtúbulos.
- La regulación de la actino-miosina del músculo liso, músculo esquelético, de las fibras de estrés y de la terminal del huso mitótico.
- La regulación de la propagación de los fibroblastos.
La inhibición de la calmodulina produce diversas alteraciones, entre las que se mencionan: alteración en la concentración intracelular de electrolitos, anomalías en la forma y división celular, alteraciones en las fibras de estrés y en el crecimiento de los fibroblastos.
La procaína ejerce una acción citotóxica sobre los fibroblastos, produciendo vacuolización, redondeo de las células, retracción desde la superficie en la que las células están creciendo, disminución de la motilidad, inhibición de la proliferación y cambios en la organización de la actina del citoesqueleto, induciendo la desaparición total de las fibras de estrés 20,22,23.
La procaína produce vacuolización masiva de fibroblastos cutáneos y otras células 14. La vacuolización es un proceso reversible, o no, sujeto al daño estructural celular ocasionado por el anestésico y al tiempo de exposición entre la droga y la célula; es celular dependiente 24. La vacuolización intensa y prolongada conduce a la muerte celular por lisis o por apoptosis 24.
La vacuolización inducida por procaína, es consecuencia de un aumento de la captación de líquido extracelular e inhibición de la liberación de líquido, captado por endocitosis y deterioro del tráfico de membrana entre los endosomas 14,25. También podrían alterar el ciclo de los receptores, si las vacuolas se originan desde los endosomas 14.
La vacuolización se inicia en el citoplasma perinuclear y progresan hacia la periferia. La procaína ingresa a organelas ácidas (trans-Golgi, endosomas, lisosomas y mitocondrias), se carga positivamente a bajo pH, ingresa líquido por vía osmótica en las vesículas y la droga queda atrapada en estas vacuolas 14. Morissette y col 14 demostraron, con tinciones celulares, que las vacuolas gigantes se originan del Golgi (tinción "ceramide-C5") y de los lisosomas (tinción "Lyso-Tracker").
La formación de vacuolas y acumulación de la droga dentro de las mismas, expone a las células a un aumento de concentración de droga intracelular, a la que las organelas son expuestas determinando toxicidad celular. Esto se ha demostrado a nivel axonal, donde se produce una disfunción celular, deterioro del movimiento y transporte de organelas, activación de caspasas (enzimas claves en la apoptosis) y degeneración dendrítica 26.
A nivel nuclear, la procaína actúa como un inhibidor no nucleósido de la metilación del ADN en determinadas líneas celulares, produciendo desmetilación e hipometilación global del ADN 6, al unirse al ADN rico en G-C (Guanina-Citosina) y alterar el funcionamiento de la metiltransferasa. A su vez reactiva genes silenciados por hipermetilación (Ej: RARβ2) 14,27,28,29.
También y a nivel nuclear, la procaína altera el ciclo celular. Produce una detención del mismo en fase "M" y retarda las fases "S" y "G2", ejerciendo así un efecto inhibidor del crecimiento y proliferación de diferentes células, incluidos los fibroblastos 14,27,28,29 (Gráfico IV). Ambas propiedades hacen de la procaína una sustancia potencialmente interesante, para su estudio como posible terapia antitumoral, pues ejerce un efecto inhibidor del crecimiento celular, demostrada ya en células de hepatoma y de cáncer mamario humano 27,28,30.
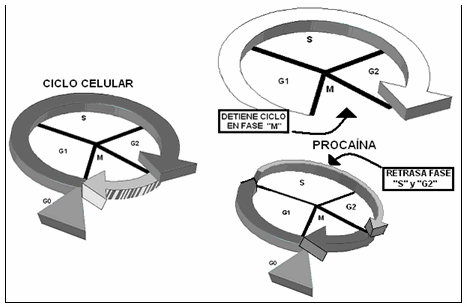
Gráfico IV: ciclo celular: acción de la procaína sobre el ciclo celular, con detención en fase "M" y retraso de las fases "S" y "G2".
A nivel muscular y continuando con la acción sobre el calcio, la procaína inhibe la liberación del mismo desde el retículo sarcoplásmico 12,31,32, ya que funciona como un antagonista de la "liberación de calcio inducida por calcio" (CICR = Calcium-Induced Calcium Release) y produce aumento de la concentración del ion dentro del retículo 12,33.
Este aumento también se ve favorecido, porque la procaína bloquea selectivamente los canales de potasio de la membrana del retículo 12.
Otra de las acciones es la de antagonizar la liberación de calcio, inducida por cafeína e inhibe la contracción muscular producida por la misma 12. La cafeína es considerada un potenciador del CICR 34,35. Antagoniza el efecto inhibitorio de la estabilización de membranaza de la procaína 36. A causa de este antagonismo, se contraindicaría el empleo conjunto de procaína y cafeína 31,34.
El consumo de oxígeno, a nivel muscular y mitocondrial, también se ve afectado por la acción del anestésico 2. Este muestra un efecto paradójico dosis-dependiente 37. A menor concentración de procaína mayor consumo de oxígeno, mientras que a elevadas concentraciones, el consumo de oxígeno (mitocondrial y vesicular de membrana) disminuye 37
El efecto dosis dependiente también se ve a nivel de los glóbulos rojos, en relación a la alteración de la movilidad y de la transformación morfológica 9,38. A dosis bajas de anestésico, aumenta la movilidad celular sin alterar su forma, mientras que a dosis altas disminuye la movilidad y se altera la forma del glóbulo, que de un aspecto bicóncavo normal se convierte a un estomatocito. Esto último lleva a una agregación (aglutinación) celular, a cambios en las cargas de superficie y a un menor transporte de oxígeno 38.
La procaína modula la respuesta inflamatoria e inmune celular, porque inhibe la activación del factor de necrosis tumoral alfa (FNT-α) y la respuesta de los neutrófilos, además deteriora la respuesta de las células Natural Killer (NK Cell) 39, pudiendo también llegar a producir una modulación del sistema inmune, por la inhibición del ADN metiltransferasa en líneas de células "T" 28,40.
El anestésico suprime la activación de la NADPH-oxidasa, la formación de superóxido (O2-) 41, inhibiendo el metabolismo y la fagocitosis leucocitaria, la adherencia y locomoción de los macrófagos, la liberación de enzimas lisosomales de los neutrófilos 41,42, al igual que la citolisis y la unión de los NK-Cell 42 a células diana. Estas alteraciones a nivel inmunitario estarían provocadas por: deterioro de los microfilamentos (esenciales para el reconocimiento y unión de las NK-Cell a las células target); cambios conformacionales de las proteínas de membrana; deterioro de los microtúbulos (esencial para la liberación de enzimas granulares de los NK-Cell) o por inhibición de la fosfolipasa A2 (relacionada con la activación de los mecanismos líticos) 42.
La fosfolipasa A2 es una enzima implicada en varios procesos biológicos. Interactúa directamente con la membrana, modificando su estado físicoquimico e interviene en el control del crecimiento celular, en las señales de transducción de la cascada inflamatoria, en la lipólisis y entre otras, en los mecanismos líticos celulares 43,44,45.
La procaína inhibe la actividad de la fosfolipasa A2 en sistemas liposomales, mitocondriales y de membrana. Esta inhibición es consecuencia de la alteración en la unión por el calcio, en la propia enzima o en el complejo enzima sustrato.
La inhibición de la acción de la fosfolipasa A2 impide la liberación del ácido araquidónico, desde los fosfolípidos de la membrana celular y por lo tanto, la formación de elementos esenciales en los procesos inflamatorios como los leucotrienos, prostaglandinas, prostaciclinas y tromboxanos 46,47.
La vía lipolítica, activada por diversas sustancias, hormonas, enzimas y receptores, se ve afectada bajo la acción de los anestésicos locales.
La supresión de la lipólisis, por acción de la procaína, se debe a diversos mecanismos entre los que se mencionan: inhibición de la fosfolipasa A2, inhibición de la unión de agonistas a receptores β2 adrenérgicos (por modificación del receptor) 48, bloqueo de la enzima adenilciclasa (por bloqueo del sistema primer mensajero adenilciclasa-ATP-AMPc intracelular) 48 y entre otros, por alteración de la concentración de calcio (bloquea la lipólisis estimulada por hormonas) 49,50.
Otros de los receptores afectados por la procaína son los colinérgicos (presentes en células endoteliales vasculares, queratinocitos, células epiteliales, linfocitos y células del sistema nervioso central); los receptores TRPV1 (Transient Receptor Potential cation channel) implicados en la transducción nociceptiva periférica y los receptores NMDA (N-methyl-D-aspartic acid) relacionado con la sensibilización del dolor 4,51,52,53,54. La procaína inhibe competitivamente los receptores colinérgicos (nicotínicos y muscarínicos) 55. En los receptores TRPV1 inducen una activación y sensibilización, mientras que los receptores NMDA son inhibidos.
CONCLUSIÓN
La capacidad de los anestésicos locales de fluidificar, expandir, engrosar o derretir los fosfolípidos de membrana, sumado a las acciones que ejercen en las proteínas (estructurales, receptores, etc.), son un punto de gran consideración a tener en cuenta, cuando se realizan las formulaciones que se emplean durante el acto mesoterapéutico, ya que las alteraciones de la membrana conducen a una modificación de la acción original de la droga, cuyos beneficios deseamos emplear.
Como regla debiéramos decir que bajas concentraciones de procaína y aplicadas en pequeñas superficies, aumentan los efectos biológicos celulares, como es la aceleración del metabolismo, la mayor movilidad celular, el aumento del consumo de oxígeno y el aumento del funcionamiento de la mitocondria. A la inversa, dosis elevadas (tanto en concentración de droga, como en superficie de aplicación) producen intensas modificaciones con cambios en la morfología y función celular.
"Como se comprenderá, y gracias al conocimiento de la acción de este anestésico tipo éster sobre las diferentes células, la procaína no debiera ser un "vector" o un "diluyente" de las diferentes fórmulas empleadas en mesoterapia, sino que debiera ser incorporada como un fármaco con su propia acción y recurrir a ella, como monodroga, sólo en determinadas patologías".
1. Barash PG, Cullen BF, Stoelting RK, Cahalan MK y Stock MC. Handbook of Clinical Anesthesia. Capítulo 21. Local Anesthetics. Sexta Edición. 2009; 308-322. [ Links ]
2. Miller RD. Miller's Anesthesia. Séptima Edición. Churchill-Livignstone. 2010; 913-940. [ Links ]
3. Covino BJ. Physiology and Pharmacology of Local Anesthetic Agents. Anesthesia Progress 1981; 98-104. [ Links ]
4. Low PS, Lloyd DH, Stein TM y Rogers III JA. Calcium Displacement by Local Anesthetics. Dependence on pH and anesthetic charge. J Biol Chem 1979; 254 (10): 4119-4125. [ Links ]
5. Nicolson GL, Smith JR y Poste G. Effects of local anesthetics on cell morphology and membrane-associated cytoskeletal organization in BALB/3T3 cells. J Cell Biol 1976; 68: 395-402. [ Links ]
6. Jang H, Lee C, Choi M, Shin S, Chung I y Yun I. Effects of Local Anesthetics on Rotational Mobility of n-(9-Anthroyloxy) stearic Acid in Neuronal Membranes. Korean J Physiol Pharmacol 2003; 7: 119-124. [ Links ]
7. Rabinovitch M y Destefano MJ. Cell shape changes induced by cationic anesthetics. J Exp Med 1976; 143: 290-304. [ Links ]
8. Lenaz G, Curatola G, Mazzanti L y Parenti-Castelli G. Biophysical studies on agents affecting the state of membrane lipids: biochemical and pharmacological implications. Mol Cell Biochem. 1978; 22 (1): 3-32. [ Links ]
9. Palek J, Liu A, Liu D, Snyder M, Fortier NL, Njoku G, Kiernan F, Funk D y Crusberg T. Effect of Procaine HC1 on ATP: Calcium-dependent Alterations in Red Cell Shape and Deformability. Blood 1977; 50 (1): 155-164. [ Links ]
10. Eichhorn JH y Peterkofsky B. Local anesthetic-induced inhibition of collagen secretion in cultured cells under conditions where microtubules are not depolymerized by these agents. J Cell Biol 1979; 81: 26-42. [ Links ]
11. Rom S y Seeman P. All lipid-soluble anesthetics protect red cells. Nat New Biol 1971; 231: 284-285. [ Links ]
12. Stephenson DG y Wendt IR. Effects of procaine on calcium accumulation by the sarcoplasmic reticulum of mechanically disrupted rat cardiac muscle. J Physiol 1986; 373: 195-207. [ Links ]
13. Poste G, Papahadjopoulos D y Nicolson GL. Local anesthetics affect transmembrane cytoskeletal control of mobility and distribution of cell surface receptors. Cell Biology 1975; 72 (11): 4430-4434. [ Links ]
14. Morissette G, Moreau EC, Gaudreault R y Marceau F. Massive cell vacuolization induced by organic amines such as procainamide. J Pharmacol Exper Therap 2004; 310 (1): 395-406. [ Links ]
15. Hahnenkamp K, Theilmeier G, Van Aken HK y Hoenemann CW. The effects of local anesthetics on perioperative coagulation, inflammation, and microcirculation. Anesth Analg 2002; 94: 1441-1447. [ Links ]
16. Raucher D y Sheetz MP. Phospholipase C activation by anesthetics decreases membrane-cytoskeleton adhesión. J Cell Sci 2001; 114 (20): 3759-3766. [ Links ]
17. Brower MC y Johnson MR. Adverse effects of local anesthetic infiltration on wound healing. Reg Anesth Pain Med 2003; 28 (3): 233-240. [ Links ]
18. Chvapil M, Hameroff SR, O'Dea K y Peacock EE Jr. Local anesthetics and wound healing. J Surg Res 1979; 27 (6): 367-371. [ Links ]
19. Morris T y Appbly R. Retardation of wound healing by procaine. Br J Surg 1980; 67 (6): 391-392. [ Links ]
20. Pierzchalska M, Michalik M, Stepien E y Korohoda W. Changes in morphology of human skin induced by local anaesthetics: role of actomyosin contraction. Eur J Pharmacol 1998; 358 (2): 235-244. [ Links ]
21. Nelson GA, Andrews ML y Karnovsky MJ. Participation of Calmodulin in Immunoglobulin Capping. J Cell Biology 1982; 95: 771-780. [ Links ]
22. Sturrock JE, Phil M y Nunn JF. Cytotoxic effects of procaine, lignocaine and bupivacaine. Br J Anaesth 1979; 51 (4): 273-281. [ Links ]
23. Gail MH y Boone CW. Procaine inhibition of fibroblast motility and proliferation. Exp Cell Res 1972; 73 (1): 252-255. [ Links ]
24. Henics T y Wheatley DN. Cytoplasmic vacuolation, adaptation and cell death: A view on new perspectives and features. Biol Cell 1999; 91: 485-498. [ Links ]
25. Michalik M, Pierzchalska M, Pabianczyk-Kulka A y Korohoda W. Procaine-induced enhancement of fluid-phase endocytosis and inhibition of exocytosis in human skin fibroblasts. Eur J Pharmacol 2003; 475 (1-3): 1-10. [ Links ]
26. Yaksh TL. Vacuoles: a hollow threat? J Can Anesth 2010; 57: 195-200. [ Links ]
27. Villar-Garea A, Fraga MF, Espada J y Esteller M. Procaine Is a DNA-demethylating Agent with Growth-inhibitory Effects in HumanCancer Cells. Cancer Research 2003; 63: 4984-4989. [ Links ]
28. Tada M, Imazeki F, Fukai K, Sakamoto A, Arai M, Mikata R, Tokuhisa T y Yokosuka O. Procaine inhibits the proliferation and DNA methylation in human hepatoma cells. Hepatol Int 2007; 1: 355-364. [ Links ]
29. Morissette G, Germain L y Marceau F. The antiwrinkle effect of topical concentrated 2-dimethylaminoethanol involves a vacuolar cytopathology. Br J Dermatol 2007; 156: 433-439. [ Links ]
30. Dueñas-González A, García-López P, Herrera LA, Medina-Franco JL, González-Fierro A y Candelaria M. The prince and the pauper. A tale of anticancer targeted agents. Molecular Cancer 2008; 7:82. http://www.molecular-cancer.com/content/7/1/82. [ Links ]
31. Uneyama H, Uneyama C y Akaike N. Intracellular mechanisms of cytoplasmic Ca2+ oscillation in rat megakaryocyte. J Biol Chem 1993; 268 (1): 168-174. [ Links ]
32. Gao Z, Xu Z, Hung MS, Lin YC, Wang T, Gong M, Zhi X, Jablons DM y You L. Procaine and procainamide inhibit the Wnt canonical pathway by promoter demethylation of WIF-1 in lung cancer cells. Oncol Rep 2009; 22 (6): 1479-1484. [ Links ]
33. Zahradnikova A y Palade P. Procaine effects on single sarcoplasmic reticulum Ca2+ release channels. Biophys J 1993; 64: 991-1003. [ Links ]
34. Klein MG, Simon BJ y Schneider MF. Effects of procaine and caffeine on calcium release from the sarcoplasmic reticulum in frog skeletal muscle. J Physiol1992; 453: 341-366. [ Links ]
35. Fujii K, Miyahara H y Suzuki H. Comparison of the effects of caffeine and procaine on noradrenergic transmission in the guinea-pig mesenteric artery. Br J Pharmacol 1985; 84: 675-684. [ Links ]
36. Hong CY, Chaput de Saintonge DM y Turner P. the inhibitory action of procaine, (+)-propranolol and (±)-propranolol on human sperm motility: antagonism by caffeine. Br J Clin Pharmacol 1981; 12: 751-752. [ Links ]
37. Saad KHM. A Link Between the Effect of Procaine on Oxygen Consumption and Its Effect on The Mechanical Activities of The Rat Ileum Smooth Muscle. J Sci Appl 2009; 3 (1): 1-11. [ Links ]
38. Sowemimo-Coker SO y Meiselman HJ. Effect of procaine hydrochloride on the electrophoretic mobility of human red blood cells. Cell Biophys 1989; 15: 235-248. [ Links ]
39. Kawasaki T, Kawasaki C, Ogata M y Shigematsu A. The effect of local anesthetics on monocyte mCD14 and human leukocyte antigen-DR expression. Anesth Analg 2004; 98: 1024-1029. [ Links ]
40. Scheinbart LS, Johnson MA, Gross LA, Edelstein SR y Richardson BC. Procainamide inhibits DNA methyltransferase in a human T cell line. J Rheumatol 1991; 18 (4): 530-534. [ Links ]
41. Hattori M, Dohi S, Nozaki M, Niwa M y Shimonaka H. The inhibitory effects of local anesthetics on superoxide generation of neutrophils correlate with their partition coefficients. Anesth Analg 1997; 84: 405-412. [ Links ]
42. Takagi S, Kitagawa S, Oshimi K, Takaku F y Miura Y. Effect of local anaesthetics on human natural killer cell activity. Clin Exp Immunol 1983; 53: 477-481. [ Links ]
43. Gudmand M, Rocha S, Hatzakis NS, Peneva K, Müllen K, Stamou D, Uji-I H, Hofkens J, Bjørnholm T y Heimburg T. Influence of lipid heterogeneity and phase behavior on phospholipase A2 action at the single molecule level. Biophys J 2010; 98: 1873-1882. [ Links ]
44. Ling Wee C, Balali-Mood K, Gavaghan D y Sansom MSP. The interaction of phospholipase A2 with a phospholipid bilayer: Coarse-Grained Molecular Dynamics Simulations. Biophys J 2008; 95: 1649-1657. [ Links ]
45. Lehtonen JKA y Kinnunen PKJ. Phospholipase A2 as a mechanosensor. Biophys J 1995; 68: 1888-1894. [ Links ]
46. Hendrickson HS y van Dam-Mieras MCE. Local anesthetic inhibition of pancreatic phospholipase A2 action on lecithin monolayers. J Lipid Res 1976; 17: 399-405. [ Links ]
47. Hendrickson HS. The penetration of local anesthetics into phosphatidyIchoIine monoIayers. J Lipid Res 1976; 17: 393-398. [ Links ]
48. Butterworth J, James RL y Grimes J. Structure-Affinity relationships and stereospecificity of several homologous series of local anesthetics for the ß2-Adrenergic receptor. Anesth Analg 1997; 85: 336-342. [ Links ]
49. D'Costa MA y Angel A. Inhibition of hormone-stimulated lipolysis by clofibrate. A possible mechanism for its hypolipidemic action. J Clin Invest 1975; 55: 138-148. [ Links ]
50. Siddle K y Hales CN. The action of local anaesthetics on lipolysis and on adenosine 3': 5'-Cyclic Monophosphate content in isolated rat fat-cells. Biochem J 1974; 142: 345-351. [ Links ]
51. Fujii T, Masai M, Misawa H, Okuda T, Takada-Takatori Y, Moriwaki Y, Haga T y Kawashima K. Acetylcholine synthesis and release in NIH3T3 cells coexpressing the high-affinity choline transporter and choline acetyltransferase. J Neurosci Res 2009; 87 (13): 3024-3032. [ Links ]
52. Benson BE, Carson RE, Kiesewetter DO, Herscovitch P, Eckelman WC, Post RM y Ketter TA. A Potential Cholinergic Mechanism of Procaine's Limbic Activation. Neuropsychopharmacology 2004; 29: 1239-1250. [ Links ]
53. Leffler A, Fischer MJ, Rehner D, Kienel S, Kistner K, Sauer SK, Gavva NR, Reeh PW y Nau C. The vanilloid receptor TRPV1 is activated and sensitized by local anesthetics in rodent sensory neurons. J Clin Invest 2008; 118: 763-776. [ Links ]
54. Sugimoto M, Uchida I y Mashimo T. Local anaesthetics have different mechanisms and sites of action at the recombinant N-methyl-D-aspartate (NMDA) receptors. Br J Pharmacol 2003; 138: 876-882. [ Links ]
55. Cuevas J y Adams DJ. Local anaesthetic blockade of neuronal nicotinic Ach receptor-channels in rat parasympathetic ganglion cells. Br J Pharmacol 1994; 111: 663-672. [ Links ]

