










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista industrial y agrícola de Tucumán
versão On-line ISSN 1851-3018
Rev. ind. agric. Tucumán vol.89 no.2 Las Talitas Jul./dez. 2012
ARTÍCULOS ORIGINALES
Interacción genotipo por ambiente en cultivares de poroto para el Noroeste Argentino
Oscar N. Vizgarra*; Stephen Beebe**; Francisco J. Morales**; Carlos H. Bellone*** y L. Daniel Ploper****
*Sección Granos, EEAOC. poroto@eeaoc.org.ar
**CIAT, Cali, Colombia.
*** Cátedra de Microbiología, Facultad de Agronomía y Zootecnia, UNT.
****Sección Fitopatología, EEAOC.
Resumen
El poroto (Phaseolus vulgaris L.) es una especie fuertemente influenciada por el medio ambiente. En este trabajo se realizó un análisis retrospectivo de las respuestas de cultivares de poroto blanco y negro a los diferentes ambientes de producción del Noroeste Argentino (NOA). El objetivo fue caracterizar e identificar los ambientes de producción de porotos en el NOA, con el fin de seleccionar materiales de mejoramiento con mayor potencial de adaptación en las diferentes zonas de producción. En una primera etapa, se realizó un análisis sobre los datos de rendimiento (kg/ha) de los genotipos en los ambientes en que fueron conducidos los ensayos regionales, durante el período 1982-1999. El diseño fue en bloques con tres repeticiones. Las parcelas fueron de cuatro líneas de 6 m de largo, distanciadas entre sí a 0,70 m. La siembra y cosecha se realizaron en forma manual. Se cosecharon las dos líneas centrales, que fueron referidas a kg de grano seco/ha. Las comparaciones de medias se realizaron mediante la prueba de diferencia límite significativo. En una segunda etapa, se utilizó el programa SEQRET en porotos negros y blancos por separado, para evaluar matrices de incidencia y de proximidades entre ambientes. Estos análisis permitieron agrupar ambientes según la reacción de diferentes tipos y colores de grano, demostrando que los porotos negros (mesoamericanos) y blancos (andinos) tienen una respuesta diferencial ante los ambientes del NOA. La Cocha es un ambiente único para la caracterización de moho blanco y antracnosis en grano negro, y Campichuelo lo es para moho blanco en poroto negro y blanco. Campichuelo, Rapelli, Viclos y Galpón sufren de otros complejos, tales como el mosaico dorado y la bacteriosis. En Monte Redondo, la mustia hilachosa es una limitante.
Palabras clave: SEQRET; Análisis retrospectivo; Phaseolus vulgaris.
Abstract
Bean crop genotype-environment interaction in North Western Argentina
The common bean (Phaseolus vulgaris L.) is strongly influenced by environmental factors. In this work, we retrospectively analyze the responses of black and white-seeded common bean genotypes in different crop production environments in North Western Argentina (NOA). The main objective was to characterize and identify the most suitable common bean production environments to select common bean genotypes adapted to the environmental and phytosanitary conditions of the bean production regions evaluated. Initially, yield data for black and white common bean genotypes planted in different environments in NOA during the 1982-1999 regional evaluation trials were analyzed. Variables assessed throughout the considered period were yield, genotypes and environments. At the second stage, SEQRET software was used to evaluate black and white common bean genotypes separately. These data were used to calculate incidence and proximity matrices among environments. SEQRET analysis allowed grouping environments according to reactions of different bean grain colors: black beans (Mesoamerican) and white beans (Andean) responded differently to environments in NOA. La Cocha is a unique environment for white mold and anthracnose incidence on black bean, and Campichuelo is an environment where white mold affects both black and white beans. Campichuelo, Rapelli, Viclos and Galpón locations are conducive to common bacterial blight and Bean golden mosaic virus incidence. In Monte Redondo, collar rot disease is a constraint on bean production.
Key words: SEQRET; Retrospective analysis; Phaseolus vulgaris.
Introducción
El cultivo del poroto seco (Phaseolus vulgaris L.) ha sido una actividad tradicional en el Noroeste Argentino (NOA) desde comienzos del siglo XX, cuando el poroto blanco tipo Alubia fue introducido por inmigrantes españoles a la zona sur de Salta (departamentos de Rosario de la Frontera y Metán).
A partir de la década de 1970, el poroto se convirtió en un cultivo extensivo de gran importancia económica regional, cuyo destino final fue la exportación. Esto sucedió cuando se produjo un incremento considerable en la demanda de poroto en los países europeos, que repercutió en el aumento de la producción de esta leguminosa en la Argentina. Así, la producción de poroto Alubia, que en 1968 solo alcanzaba 15.000 t, pasó en 1971 a 45.000 t y en 1976 a 115.000 t, cifras que demostraron el vigoroso crecimiento de este cultivo en el NOA. En ese entonces, la Argentina ya era catalogada como el segundo país exportador mundial de porotos y único en el tipo Alubia (Voysest Voysest, 2000). Más tarde, en 2004, la Argentina se convertiría en el principal exportador mundial de este tipo de poroto y cuarto exportador mundial de porotos en general (colores negros, rojos y no tradicionales) (Vizgarra, 2004).
Al presente, el 95% de la producción porotera proviene de las provincias del NOA, en especial de Salta, el este de Tucumán, oeste de Santiago del Estero, sudeste de Jujuy y Catamarca. El considerable incremento del área sembrada con poroto en el NOA favoreció la aparición de diversos problemas de producción, particularmente de tipo fitosanitario. La primera gran epifitia ocurrió a finales de la década de 1970, cuando emergió el problema del "achaparramiento", que afectó severamente a la variedad Alubia. Esta enfermedad detiene el crecimiento y deforma totalmente la planta a una temprana edad, hasta hacerla improductiva. Este problema se hizo sentir con mayor intensidad en las áreas más calientes y secas del NOA.
La etiología del "achaparramiento del poroto" fue finalmente revelada en 1981, al concluir que se trataba de una enfermedad causada por el virus del enanismo del poroto (Bean dwarf mosaic virus o BDMV), un geminivirus transmitido por la mosca blanca (Bemisia tabaci Genn) (Morales, 1981; Gálvez y Morales, 1995). La emergencia de este virus estuvo estrechamente relacionada a las mayores poblaciones de mosca blanca que se generaron debido al avance de la soja [Glycine max (L.) Merrill] en el NOA, y a las condiciones de sequía que se presentaron en esa época, particularmente en la provincia de Santiago del Estero. Estas epifítias significaron la desaparición del cultivo del poroto blanco Alubia de las zonas tradicionales y de expansión.
La producción de Alubia se concentró, desde 1980, en el departamento San Martín, provincia de Salta, donde las condiciones productivas resultaban favorables. Es en ese momento en que se contempló la necesidad de incrementar la producción de otros tipos de poroto, en especial los de grano negro, debido a su mayor nivel de resistencia al "achaparramiento". Los primeros materiales de grano negro resistentes se introdujeron al NOA en viveros de adaptación y rendimiento, y habían sido enviados por el Programa de Frijol del Centro Internacional de Agricultura Tropical (CIAT), Palmira, Colombia. Estos materiales provenían en su mayoría de Centro América, donde habían demostrado su resistencia a otro begomovirus transmitido por B. tabaci: el virus del mosaico dorado amarillo del poroto (Bean golden yellow mosaic virus), que ataca el cultivo en esa región.
En 1983, procedente de Brasil, hizo su aparición otro virus transmitido por la mosca blanca, B. tabaci: el virus del mosaico dorado del poroto (Bean golden mosaic virus o BGMV), el que atacó los genotipos de poroto que habían mostrado resistencia al BDMV, debido a su mayor rango de patogenicidad.
Ante la situación fitosanitaria limitante para el cultivo del poroto en la región a principios de los años ochenta, la Estación Experimental Agroindustrial Obispo Colombres (EEAOC) de Tucumán inició un plan de trabajo para enfrentar estos nuevos problemas de producción de poroto, con vinculación al Programa de Poroto del CIAT. Los principales objetivos de este plan fueron: la identificación de problemas, su importancia económica y la conformación de estrategias de investigación (mejoramiento, agronomía, sanidad, calidad, etc.).
En el presente trabajo, se analiza la relación entre el medio ambiente y la adaptación de los diferentes genotipos de poroto mejorados introducidos en el NOA, con el fin de evitar las pérdidas de producción por introducción de genotipos de poroto no aptos para las condiciones ambientales y fitosanitarias de las localidades seleccionadas. Para tal fin, se aprovechó una importante cantidad de información resultante de la evaluación de un conjunto de genotipos en distintos ambientes del NOA, a lo largo de 18 años.
Sobre la base de este análisis, se procura potenciar los ambientes favorables mediante el manejo de tecnología, para una mejor relación insumo-beneficio (rentabilidad del productor) y para orientar el mejoramiento genético del cultivo.
Antecedentes de estudios de interacción genotipo-ambiente
Los estudios de interacción genotipo x ambiente (GxA) en poroto específicamente, no son tan abundantes como los realizados en otras especies, tales como maíz, cebada, trigo, etc. (Baker, 1969, 1990; Braun et al., 1992; Brennan and Fox, 1995; CIMMYT, 1979).
En los programas de mejoramiento genético de poroto, los materiales avanzados o líneas introducidas son evaluados en diferentes ambientes, para continuar el trabajo de selección con las líneas más destacadas. Existe una fuerte interacción de las líneas con el ambiente, por lo que la interacción G x A es uno de los aspectos más importantes que se deben afrontar en el mejoramiento (Allard and Bradshaw, 1964).
Muchas veces, los componentes de la variación empleados en el análisis de la varianza no llegan a demostrar la superioridad de alguna variedad, debido a que estas generalmente difieren en sus rangos relativos al ser evaluadas en diferentes años. Por esta razón se adoptó el análisis de regresión, una metodología de probada eficiencia en el estudio del comportamiento de genotipos con relación al ambiente.
Finlay and Wilkinson (1963) fueron los primeros investigadores en demostrar que la estabilidad de un genotipo puede ser expresada como una función lineal de un índice ambiental. Para ello calcularon un coeficiente de regresión (Bi), que se relaciona con la media de cada variedad (Vi), con el rendimiento promedio de todas las variedades para cada sitio y estación. Valores de coeficientes de regresión (Bi) cercanos a 1 indican estabilidad promedio; valores inferiores a 1 corresponden a cultivares con mayor resistencia a cambios varietales y valores superiores a 1 pertenecen a variedades inestables, proclives a aprovechar las condiciones ambientales asociadas a buenos rendimientos.
Eberhart and Russell (1966) cuantificaron la adaptabilidad usando, como su medida, el coeficiente de regresión de las medias varietales, contra la media general de todos los genotipos en cada ambiente. Estos autores usaron a Bi como el primer parámetro de estabilidad varietal, pero ahondaron aún más en el análisis al incorporar un segundo parámetro (F), que mide el desvío de la regresión lineal de una variedad en el índice ambiental. Valores significativos de la prueba F inducen a concluir que existe otro factor no considerado que está afectando la respuesta del genotipo al ambiente. Un cultivar deseable debería estar representado por un rendimiento de promedio alto, un coeficiente de regresión cercano a 1 (respuesta promedio) y los desvíos respecto a la regresión deberían ser lo más pequeños posibles.
De acuerdo a Hardwick and Wood (1972), el segundo parámetro de estabilidad no es funcionalmente independiente del coeficiente de regresión lineal, tanto en el sentido algebraico como biológico, pero es un resultado ineludible del ajuste de rectas a datos que solo pueden ser representados de manera apropiada en pocas dimensiones. Los procedimientos, aplicaciones de estas y otras técnicas de medición de la interacción GxA fueron revisadas por Freeman (1973).
Para muchos autores, la adaptación es la respuesta genotípica amplia a ambientes diferentes, con rendimientos estables (Allard and Bradshaw, 1964). Por esto, se emplea el término de adaptación como sinónimo de potencial de producción.
Freeman (1973) señala que la interacción genotipo x ambiente es directamente proporcional al efecto del ambiente. Baker (1969) define la interacción GxA como la diferenciación entre el valor fenotípico observado y el valor esperado de una determinada variedad en cierto ambiente. El autor logra esto con la suma de los dos componentes aditivos: genético y ambiental, indicando que el efecto del ambiente y de la interacción GxA se estima por la suma y sustracción de la expresión fenotípica.
Cuando se trabaja en forma regional en la evaluación del comportamiento de las variedades en distintas localidades a través de los años, se debe reconocer que el entorno ambiental no es homogéneo para el propósito del análisis. Por ello, es necesario agrupar las variedades por color. Para ello, se usa el método de análisis de conglomerados (cluster analysis) (Fisher and Van Ness, 1971; Mandel, 1971; Everitt, 1980; Lin, 1982).
Los primeros estudios de evaluación de respuestas de porotos en diferentes ambientes fueron los efectuados por Ghaderi et al. (1982).
En estudios de estabilidad varietal y rendimiento para porotos negros en el NOA, Vizgarra et al. (1997) emplearon las metodologías propuestas por Finlay and Wilkinson (1963) y Eberhart and Rusell (1966), que sirvieron para caracterizar el comportamiento de los principales cultivares de poroto negro. Los nuevos materiales TUC 390 y TUC 500 demostraron una diferencia productiva respecto a los cultivares tradicionales (DOR 157, BAT 304, NAG 12 y JEO) de 304 kg/ha. Esto implicaba un 23% de rendimiento adicional sobre los tradicionales, y un incremento en más de un 35% de los beneficios económicos que lograría un productor solamente con el cambio varietal.
Los citados autores, en un trabajo posterior y con el mismo conjunto de germoplasma-ambientes-rendimientos, aplicaron el método de análisis de conglomerados y lo compararon con los resultados precedentes. Este método resultó ser más preciso que la regresión lineal y los parámetros de estabilidad ambiental en la diferenciación del comportamiento varietal de los seis cultivares comerciales más difundidos de poroto negro (Vizgarra et al., 1998). Destacaron además que los cultivares de poroto negro TUC 390 y TUC 500 conformaban un conglomerado de alta capacidad de producción. Estos materiales son precoces (80 a 90 días a cosecha) y poseen una mayor capacidad, tanto adaptativa (mayor tolerancia al BGMV y a la sequía) como de producción (estructura de planta mejorada), en los diversos ambientes estudiados.
Materiales y métodos
1.a) Genotipos
Se evaluaron porotos de colores negro y blanco durante el período 1982-1999. Estos colores fueron seleccionados por conformar más del 99% del volumen de las exportaciones argentinas.
Los trabajos pioneros comenzaron con poroto negro a partir de 1981, donde se recibieron las primeras introducciones procedentes del CIAT, Colombia. En el período de trabajo correspondiente a 1982-1990, se realizó la evaluación de 105 materiales de poroto negro en diferentes ambientes del NOA. Entre 1992 y 1999, se realizó la evaluación de materiales de poroto negro con resistencia o tolerancia superior a los virus del mosaico dorado (BGMV) y mosaico enano (BDMV), y con arquitectura acorde para la trilla directa, etapa que culminó con la liberación comercial de las variedades TUC 390 y TUC 500 (Vizgarra, 1995).
Para poroto blanco, la situación consistió en llevar a cabo un trabajo de mejoramiento específico para las condiciones del NOA, situación completamente diferente a la de los porotos negros. Dado que la Argentina era el único productor mundial del tipo Alubia, el CIAT no disponía de materiales de las características requeridas en su banco de germoplasma. Es así que desde la Argentina, se le enviaron a esta institución materiales para realizar trabajos de cruzamientos, a los efectos de incorporar en ellos resistencia a las principales enfermedades, ya que el CIAT disponía de estas fuentes de resistencia, en especial para las virosis. Para porotos negros, el trabajo de mejoramiento consistió simplemente en la introducción, evaluación y selección de los genotipos adaptados a nuestras condiciones agroecológicas y con resistencia a las virosis.
En este período de trabajo, que se extendió desde 1984 a 1989, se evaluaron 140 materiales de poroto blanco en diferentes ambientes del NOA.
1.b) Experimentos a campo
Las evaluaciones formaron parte de la Red Internacional de Evaluación de Germoplasma de Fríjol del CIAT y las directrices experimentales fueron las contenidas en el Sistema Estándar para la Evaluación de Germoplasma de Fríjol (Van Schoonhoven and Pastor Corrales, 1987). El diseño experimental a campo fue en bloques completos al azar, con tres repeticiones para las variedades finales seleccionadas. Las comparaciones de medias se realizaron mediante la prueba de diferencia límite significativa (DLS) (p< 0,05). La preparación de suelos fue mecánica y tradicional (cincel, con dos rastras excéntricas). Cada parcela estuvo formada por cuatro líneas de 6 m de largo, distanciadas entre sí a 0,7 m y a 2,0 m entre bloques. La delimitación, surcado y siembra, a una densidad de 16 semillas/m lineal (densidad comercial), fueron manuales. La conducción del ensayo fue mediante prácticas comunes del gran cultivo.
Para el control de malezas se utilizó un herbicida de pre-emergencia (pendimetalina, a una dosis de 3,0 l/ha). En post-emergencia del cultivo, se aplicó 0,6 l/ha de fomesafen para el control de hoja ancha, mientras que para hoja angosta se utilizó haloxifop metil (0,5 l/ha).
El manejo de insectos, principalmente mosca blanca, se realizó con aplicaciones de dimetoato (0,35 l/ha) a los 10 días de emergencia de las plántulas, momento de mayor susceptibilidad del cultivo. En caso de ambientes con alta presión de mosca o ensayos ubicados muy próximos al monte o al cultivo de soja, esta aplicación se repitió a los 10 días.
Se utilizó el fungicida preventivo Benlate (0,5 kg/ha) a los 30 días del cultivo, tanto en poroto negro como en blanco.
Se cosecharon las dos líneas centrales y se descartaron, a manera de bordura, 0,5 m en cada cabecera (arrancado, hilerado, secado al sol, embolsado); la trilla (batido y separación) y clasificación (zarandeo y selección) se llevaron a cabo en forma manual. La limpieza fue mediante corriente de aire (mecánica). El grano/semilla limpio se pesó en balanza electrónica y se transformó a kg/ha.
Para la prueba de calidad comercial, se remitieron muestras a la Cámara de Legumbres de la República Argentina (CLERA), donde se tipificaron acorde a los patrones comerciales de exportación y mercadeo establecidos.
2- Ambientes
2.a) Determinación de los principales ambientes de producción
El análisis de repetición de localidades a través de los años, realizado mediante la sub-rutina SEQELM (Sequential Elimination) del paquete Sequential Retrospective (SEQRET), identificó a las siguientes localidades (análisis retrospectivo multilocacional; DeLacy, 1997) (Figura 1):

Figura 1. Zonas de producción del Noroeste Argentino y ambientes (localidades) utilizados para este estudio.
2.1.1.-) Viclos (VI): departamento Leales, provincia de Tucumán, situado a LS 27º 10´ LW 64º53´, a 375 msnm
Aquí el poroto, especialmente de granos negros y rojos, se siembra en gran escala. La localidad presenta presión intermedia de virosis, en general, y de otras enfermedades, como la bacteriosis común (Xanthomonas axonopodis pv. phaseoli) y mustia hilachosa (Thanatephorus cucumeris).
2.1.2.-) La Cocha (LC): departamento La Cocha, provincia de Tucumán, ubicado en LS 27º47´LW 65º34´, a 444 msnm
La zona destinada al cultivo del poroto corresponde a la región Este, donde se nota una presión intermedia a las virosis. Se siembran porotos negros, rojos y blancos. Otras enfermedades que se presentan, en menor escala, son bacteriosis común, mancha angular (Phaeoisariopsis griseola) y antracnosis (Colletotrichum lindemuthianum). Estas enfermedades aparecen cuando se dan las condiciones ambientales predisponentes.
2.1.3.-) Rapelli (RA): departamento Burruyacu, provincia de Tucumán, ubicado en LS 26º45´ y LW 64º30´, a 402 msnm
La localidad y sus alrededores constituyen una típica zona porotera cálida y con alta presión de virosis, bacteriosis común y mustia hilachosa. El color predominante del poroto es el negro.
2.1.4.-) Monte Redondo (MR): ubicado en el departamento Cruz Alta, Tucumán, con una posición LS 26º57´ LW 64º54´, a 410 msnm
Es un ambiente en general caliente, que presenta incidencia de mosca blanca, vector de los begomovirus del mosaico dorado y mosaico enano. Otras enfermedades importantes son la bacteriosis común y la mustia hilachosa. En este ambiente predomina el cultivo del poroto negro, mientras que una amplia región se encuentra cultivada con soja y maíz.
2.1.5.-) Rosario de la Frontera (RF): departamento Rosario de la Frontera, provincia de Salta, localizado a 26º48´ LS y 64º58´LW, a 791 msnm
Zona tradicionalmente porotera desde sus inicios, con presencia baja a intermedia de virosis e intermedia a alta de bacteriosis y mancha angular. Se siembran porotos de grano negro, rojo y, en menor medida, blanco.
2.1.6.-) El Galpón (GA): departamento Anta, provincia de Salta, ubicado a 25º24´LS y 64º39´LW, a 625 msnm
Zona más caliente, donde la presión de virosis es más acentuada. Resulta también importante la bacteriosis común y la mustia hilachosa. Predomina la siembra de poroto negro.
2.1.7.-) Campichuelo (CA): departamento San Martín, provincia de Salta, localizado a 23º07´LS y 64º00´LW, a 308 msnm
La localidad y sus alrededores son de reciente incorporación al cultivo del poroto. Es predominantemente una zona de poroto blanco y, en menor medida, de poroto rojo y otros colores no tradicionales. Es una zona caliente, con presencia de cultivos de soja y algodón en los últimos años, lo que trajo aparejado un aumento de la presión de mosca blanca y virus. Otras enfermedades importantes son el moho blanco (Sclerotinia sclerotiorum), la bacteriosis común y la mustia hilachosa.
3- Análisis de datos
Para el análisis de los datos de rendimientos (kg de grano seco/ha) de los distintos genotipos de porotos en los diferentes ambientes del NOA a través de 18 años sucesivos, se utilizó el programa computarizado SEQRET, mediante el cual se analizaron los colores negros, rojos y blancos en forma separada. El programa está conformado por tres subrutinas:
a-) incidencia de los arreglos en modelos de análisis retrospectivos;
b-) reglas para la eliminación de celdas vacías y
c-) reubicación de las localidades eliminadas.
A partir de los datos generados para cada año-localidad-genotipo y variable de respuesta, y habiendo sido expresados los rendimientos como kg de grano seco/ha en este caso, se procedió a evaluar matrices de incidencia, en las cuales se registró el número de veces que un genotipo fue usado en cada combinación año-localidad. Se obtuvieron indicadores para cada localidad y para cada año, que es un conteo del número de veces que apareció el genotipo.
Si una combinación año-localidad-genotipos aparece muchas veces, esto constituye un indicador lógico de la credibilidad que puede tener el dato del comportamiento de la interacción genotipo-ambiente más conocido, más creíble. Si, por el contrario, aparece pocas veces, poco se podría creer sobre el desempeño del genotipo en esos ambientes.
Con la información precedente, se calculó una matriz de proximidades entre localidades con distancias euclidianas cuadráticas, a partir de los datos estandarizados por ambientes y ponderados por los años en los que la comparación era posible. Si dos localidades no habían sido usadas en el mismo año, estas no pudieron compararse, y hubo una celda faltante para ellas, en la matriz de proximidades. El método eliminó aquellas localidades que no pudieron ser comparadas o que tenían celdas faltantes. Se garantizó que se retuvieran aquellas localidades con el mayor número de comparaciones, o sea que se eliminaron filas (genotipos) y/o columnas (ambientes), según el número de datos faltantes que tuvieron en forma sucesiva, hasta que la matriz de proximidades no tuvo datos faltantes.
Con la matriz reducida, consistente en las localidades retenidas para cada año, se efectuó un análisis jerárquico de conglomerados, que permitió la elaboración de un dendograma para ilustrarlo. El método empleado fue el de suma de cuadrados incremental o método de Mard. El número de grupos pudo juzgarse por la bondad del modelo expresado por R2.
Adicionalmente, se realizó un análisis de coordenadas principales para las localidades retenidas en cada año, el cual generó la información que permitió la representación gráfica de un biplot.
En este punto, algunas de las localidades eliminadas pudieron reingresar al análisis, mediante el cálculo de la distancia cuadrática euclidiana de una de tales localidades al centroide de cada uno de los grupos definidos. La asignación se le efectuó al grupo donde se logró la menor distancia.
Se realizaron dos análisis en el caso de los porotos de grano negro y blanco. Para el primero, se consideró el periodo completo desde 1982 a 1999, y en el segundo análisis, solamente el periodo de 1992 a 1999. Esto se hizo ante la factibilidad de que el ingreso de materiales resistentes a virosis en los ensayos en la última década pudiese haber reducido la interacción de GxA, y así haber cambiado la relación entre ambientes.
Resultados y discusión
1.- Agrupación por dendogramas. Genotipos y ambientes
La subrutina SEQCLU (Sequential Clustering) brindó los valores de caracterización y agrupamiento de los distintos genotipos (negros y blancos) a través de los ambientes de evaluación para el período 1982 a 1999 inclusive, que se resumen en las Figuras 2, 3, 4 y 5.

Figura 2. Agrupación en dendogramas para distintos ambientes del Noroeste Argentino (NOA), construidos en base a los datos de genotipos de porotos negros, periodo 1982 a 1999.
CA=Campichuelo, GA=Galpón, LC=La Cocha, MR=Monte Redondo, RF=Rosario de la Frontera, RA= Rapelli y VI=Viclos.

Figura 3. Agrupación en dendogramas para genotipos de grano negro, evaluados en distintos ambientes del NOA, período 1992 a 1999.
CA=Campichuelo, LC=La Cocha, MR=Monte Redondo y VI=Viclos.
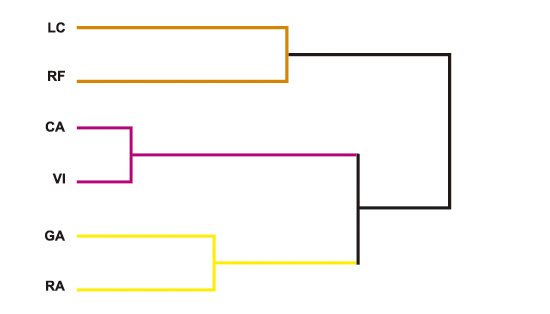
Figura 4. Agrupación en dendogramas para genotipos de poroto blanco, evaluados en distintos ambientes del NOA, período 1982 a 1991.
CA=Campichuelo, GA=Galpón, LC=La Cocha, RF=Rosario de la Frontera, RA= Rapelli y VI=Viclos.

Figura 5. Agrupación en dendogramas para genotipos de grano blanco, evaluados en distintos ambientes del NOA, período 1992 a 1999.
CA=Campichuelo, LC=La Cocha, MR=Monte Redondo y VI=Viclos.
1.1.- Genotipos de grano negro
La agrupación de ambientes en dendogramas para los porotos negros evaluados en el periodo de 1982 a 1999, se muestra en la Figura 2. Se aprecia que se formaron tres grupos de ambientes con comportamiento productivo contrastante, uniendo el ambiente CA al conjunto (GA+VI). De esta manera, para el período de análisis se obtuvieron tres grandes ambientes, a saber: A1= LC; A2= MR, RA, RF y A3= CA, GA, VI.
Para caracterizar el comportamiento ambiental del germoplasma de grano negro en la última década, se enfatizaron las evaluaciones que se ilustran en la Figura 3. Este análisis por necesidad incluyó menor número de ambientes, ya que algunos de ellos no cumplieron con el número mínimo de años necesarios. Solamente fue posible comparar los ambientes MR, VI, CA y LC. En la Figura 3, surge que las localidades LC y CA se comportaron como ambientes parecidos en términos de respuesta; MR fue el más distinto, seguido por VI.
En los ambientes considerados, la expresión de los rendimientos sufrió la presión de enfermedades (Tabla 1).
Tabla 1. Ambientes y niveles de afección de enfermedades en los germoplasmas de granos negros para el período 1982-1999.

CA=Campichuelo, GA=Galpón, LC=La Cocha, MR=Monte Redondo, RF=Rosario de la Frontera, RA= Rapelli y VI=Viclos.
*Escala del CIAT, donde 1, 2, 3: resistente; 4, 5, 6: intermedio y 7, 8, 9: susceptible.
En 1995, con la liberación de los cultivares TUC 390 y TUC 500, de alta tolerancia a las virosis (Vizgarra, 1995 y 1996), se logró un significativo avance en el desempeño sanitario para los distintos ambientes del NOA (Tabla 2).
En la Tabla 2, se observa que los materiales considerados tradicionales mantuvieron sus niveles de afección a enfermedades. En cambio, las nuevas variedades de poroto negro TUC 390 y TUC 500 mostraron un mejor comportamiento ante las virosis y un ligero mejoramiento respecto a bacteriosis común, aunque no así en relación a mancha angular, antracnosis y mustia hilachosa. Estos últimos dos materiales genéticamente no presentaban tolerancia a ellas, especialmente en ciertos ambientes como La Cocha, Rosario de la Frontera y Monte Redondo, donde las condiciones ambientales son favorables para su presencia.
Tabla 2. Ambientes y niveles de afección por enfermedades de genotipos de poroto negro tradicionales y nuevos para el período 1995-1999.

CA=Campichuelo, GA=Galpón, LC=La Cocha, MR=Monte Redondo, RF=Rosario de la Frontera, RA= Rapelli y VI=Viclos.
*Escala del CIAT, donde 1, 2, 3: resistente; 4, 5, 6: intermedio y 7, 8, 9: susceptible.
1.2.- Genotipos de grano blanco
La agrupación de ambientes en dendogramas para los genotipos de poroto blanco evaluados, se muestra en la Figura 4. Puede observarse que, para el período de análisis, se obtuvieron tres grandes ambientes, a saber: A1= LC, RF; A2= CA, VI y A3= GA, RA, los cuales no coinciden con las agrupaciones ambientales para germoplasma de poroto negro.
En los ambientes considerados, la expresión de los rendimientos sufrió la presión de enfermedades (Tabla 3).
Tabla 3. Ambientes y niveles de afección por enfermedades en poroto blanco para el período 1982-1999.

CA=Campichuelo, GA=Galpón, LC=La Cocha, MR=Monte Redondo, RF=Rosario de la Frontera, RA= Rapelli y VI=Viclos.
*Escala del CIAT, donde 1, 2, 3: resistente; 4, 5, 6: intermedio y 7, 8, 9: susceptible.
Para caracterizar también el comportamiento ambiental del germoplasma de grano blanco en la última década, se enfatizaron las evaluaciones que se ilustran en la Figura 5.
Este análisis, por necesidad, incluyó menor número de ambientes, ya que algunos de ellos no cumplieron con el número mínimo de años necesarios. En la Figura 5, se observa que el número de años de evaluación no permitió precisar el desempeño de la localidad MR, por lo que esta se comportó como una clave única. Así el A1= CA+VI, localidades que conformaron un ambiente similar, mientras que A2= LC. Por otro lado, las localidades de LC y CA resultaron contrastantes entre sí, ofreciendo información complementaria y no repetitiva entre ellas.
2.- Identificación de ambientes
El análisis de SEQRET reveló tres grupos de ambientes para poroto negro y tres grupos de ambientes para poroto blanco.
2.1. Poroto negro
En el caso de los porotos de grano negro, el A1= LC se diferencia de los restantes ambientes por ser más fresco y húmedo. Estas condiciones ambientales son favorables y predisponentes para la presencia de las enfermedades moho blanco y antracnosis, las que afectan en mayor medida al germoplasma de grano negro.
En los últimos años, merced a las condiciones ambientales favorables (las amplias fluctuaciones de temperatura, la humedad y luminosidad), la mancha angular se constituyó en la principal enfermedad limitante, con manifestaciones epifitóticas que causaron defoliaciones prematuras, altas pérdidas de rendimiento y baja calidad del grano obtenido. A las condiciones ambientales se le sumó la falta de rotación con cultivos de cobertura (maíz y sorgo), hechos estos que favorecieron la diseminación del inóculo.
El A2= MR, RA, RF se distingue por los factores de sequía estacional marcada, heladas tempranas, altas temperaturas y lluvias torrenciales. Dentro de ellos, la localidad RF (Rosario de la Frontera) es la más fresca y presenta un mayor número de días con nubosidad, merced a su característica pedemontana. Debido a ello, en los genotipos más susceptibles, fue posible encontrar daños por antracnosis. En las localidades RA (Rapelli) y MR (Monte Redondo), la enfermedad que se presenta como limitante es la mustia hilachosa.
Dentro de A3= CA, GA, VI, GA y VI presentan sequía estacional de corta duración, excesos hídricos breves y heladas tempranas. Ambientalmente, GA y VI son los más parecidos entre sí, seguidos por CA, que se les suma con características superiores debido a su mayor precipitación y sus heladas más tardías.
En el análisis de la Figura 2 se notó un patrón parecido al anterior en la relación de MR a los otros ambientes, en el sentido de que se diferenciaba ampliamente de los otros citados (VI, CA y LC). Por otro lado LC, donde la presencia de virosis es menor, se acercaba a los ambientes de CA y VI cuando los viveros contenían un mayor número de materiales resistentes, y el factor de virosis ya no era la causa de diferenciación entre ellos.
2.2.- Poroto blanco
La separación de ambientes por respuesta para el germoplasma de poroto blanco tiene una elevada asociación con las lluvias y con la temperatura. Así, el A1= LC, RF es más lluvioso y fresco. El poroto blanco se encuentra dentro de las razas andinas de la especie, razas que típicamente son más sensibles a altas temperaturas. No resulta extraño que las temperaturas más frescas tengan un efecto mayor en la respuesta de los porotos de grano blanco que en los de grano negro, los que pertenecen a la raza mesoamericana, que tiene mayor adaptación a las altas temperaturas y mayor amplitud en su adaptación.
En cierto grado, la respuesta de los porotos blancos pudo haber estado relacionada con las enfermedades asociadas con los ambientes. En LC y RF, ambientes más frescos y lluviosos, la antracnosis presentó mayores niveles de incidencia y afección. En los otros cuatro ambientes el mosaico dorado, la bacteriosis y la mustia se presentaron con mayor intensidad, aunque otros factores no determinados causaron una separación entre los grupos A2 (CA, VI) y A3 (GA, RA).
2.3.- Agrupados
Al comparar el agrupamiento de ambientes en conjunto para los genotipos de porotos negro y blanco, surge como similitud que la localidad LC se presenta como independiente de ambientes más cálidos. A su vez, RA se comporta en forma totalmente distinta para germoplasma de grano negro (A2) y blanco (A3). En el caso de los negros, se agrupa con RF y lejos de GA, mientras que la situación contraria se expresa con los blancos. Esto resalta otra vez la gran diferencia entre los orígenes de las razas de estos dos tipos de germoplasma de poroto. Por ejemplo, los grupos blancos y negros tienen una reacción muy diferente ante las virosis. Estas constituyen las principales limitantes para la producción de poroto blanco en RA, debido al mayor nivel poblacional de B. tabaci, el vector del BGMV y BDMV.
La utilidad de aplicación de estos resultados se observa en el esquema de prueba que se empleó de 1997 a 1999, en los ensayos conducidos en MR, CA y LC, ya que ellos aparecen en diferentes agrupaciones según el estudio actual. Según el análisis de SEQRET, estos ambientes cubren el mayor rango de la variabilidad en los ambientes poroteros del NOA. De esta manera, el análisis presentado en el actual trabajo sirve para validar la efectividad del esquema de pruebas planteado y brinda una mayor confianza a los resultados obtenidos en estos años. Ello representa un uso más eficiente de recursos, ya que no se duplican sitios similares entre sí, y simultáneamente se cubre un amplio rango de ambientes de producción de porotos. Cabe señalar que de esta forma, se identificaron genotipos como TUC 510 (poroto negro), que responden en forma superior a través de este rango de ambientes (Vizgarra et al., 2002).
Cuando se comparan sanitariamente los distintos genotipos de poroto, vale decir poroto negro (raza mesoamericana) con poroto blanco (raza andina) (Tablas 1 y 3), se aprecia que, en los ambientes de mayor presión de mosca blanca (zonas calientes RA y CA), existe un mayor impacto de las virosis en los materiales de poroto blanco. Esta diferencia relativa a favor de los materiales de grano negro se hizo aún más alta a partir de 1995, con la liberación de variedades de poroto negro con alta tolerancia a las virosis: TUC 390 y TUC 500 (Tabla 2) (Vizgarra, 1995 y 1996).
La diferencia en adaptación a diferentes ambientes entre los porotos andinos y los porotos mesoamericanos, representados por sus respectivas clases comerciales, fue reconocida por Ghaderi et al. (1982), aún antes de que los estudios de diversidad genética revelaran sus orígenes contrastantes. Los citados autores utilizaron un método convencional para analizar la interacción genotipo-ambiente (GxA), como lo es el análisis de Eberhart and Russell (1966), que sirve para cuantificar el grado de interacción de genotipos individuales, utilizando parámetros estadísticos como la regresión y la desviación de la regresión.
En el estudio actual, se emplea el programa SEQRET para agrupar ambientes y revelar diferentes patrones de adaptación y respuesta de genotipos a ambientes. Mientras SEQRET no brinda una caracterización de genotipos individuales, tiene la ventaja de poderse aplicar a estudios en los que los genotipos no son constantes a través de los años. Mientras otros métodos para agrupar ambientes requieren de una matriz de datos en los que cada genotipo aparece en cada sitio y año, SEQRET sirve para extraer información de una matriz incompleta. Esta alternativa es más práctica que mantener muchos genotipos en ensayos de rendimiento a través de los años, cuando en términos prácticos la mayoría de los cultivares/genotipos ya han perdido su utilidad y otros materiales de características superiores han sido desarrollados.
Conclusiones
Del conjunto de datos presentados, analizados y discutidos anteriormente se puede concluir que:
1. El análisis de SEQRET permitió agrupar ambientes según la reacción de diferentes clases comerciales y acervos de poroto, demostrando que los porotos negros (mesoamericanos) y blancos (andinos) tienen una respuesta diferencial ante los ambientes del NOA, posiblemente debido en parte a los factores climáticos (temperatura y lluvia) y también a las enfermedades endémicas en estos ambientes.
2. El A1 LC (La Cocha) es un ambiente único para la caracterización de tolerancia a las enfermedades endémicas de clima fresco, tales como mancha angular, moho blanco y antracnosis, en germoplasma de grano negro.
3. Ambientes como CA, RA, VI, y GA sufren de otros complejos de enfermedades, que incluyen el mosaico dorado y la bacteriosis, además de moho blanco para el caso específico de Campichuelo.
4. MR es un ambiente donde la mustia hilachosa es una limitante fúngica crítica, hecho que se refleja en una correlación negativa con lluvias en la parte media del ciclo del cultivo.
5. CA es un ambiente único para la caracterización de las enfermedades endémicas de clima con temperaturas moderadas a altas y elevada humedad, tales como moho blanco, que afectan a los porotos negro y blanco.
6. El análisis de SEQRET ha confirmado la práctica actual de utilizar los ambientes MR, CA y LC como representativos de la amplitud de ambientes en el NOA, para la prueba de nuevos materiales de poroto.
Bibliografía citada
1.Allard, R. W. and A. D. Bradshaw. 1964. Implications of genotype environmental interaction in applied plant breeding. Crop Sci. 4: 503-508. [ Links ]
2. Baker, R. J. 1969. Genotype-environment interactions in yield of wheat. Can. J. Plant Sci. 49: 743-751. [ Links ]
3. Baker, R. J. 1990. Crossover genotype-environmental interaction in spring wheat. En: Kang, M. S. (ed.), Genotype-by-environment interaction and plant breeding, Louisiana State University, Baton Rouge, Louisiana, USA, pp. 42-51. [ Links ]
4. Braun, H. J.; W. H. Pfeiffer and W. H. Pollmer. 1992. Environments for selecting widely adapted spring wheat. Crop Science 32: 1420-1427. [ Links ]
5. Brennan, J. P. and P. N. Fox. 1995. Impact of CIMMYT wheats in Australia: evidence of international research spillovers. Economics Research Report Nº1/95. NSW Agriculture, Wagga Wagga, Australia. [ Links ]
6. Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT). 1979. International testing program in wheat, triticale and barley. CIMMYT today (10). [ Links ]
7. DeLacy, I. H. 1997. Retrospective pattern analysis of plant breeding multi-environment trials. Tesis doctoral inédita. Department of Agriculture, University of Queensland, Australia. [ Links ]
8. Eberhart, S. A. and W. A. Russell. 1966. Stability parameters for comparing varieties. Crop Sci. 6: 36-40. [ Links ]
9. Everitt, B. S. 1980. Cluster analysis. 2. ed. Heinemn Educational Books Ltd., London, England. [ Links ]
10. Finlay, K. W. and G. M. Wilkinson. 1963. The analysis of adaptation in a plant breeding program. J. Agric. Res. 14: 742-754. [ Links ]
11. Fisher, L. and J. W. Van Ness. 1971. Admissible clustering procedures. Biometrika 58: 91-104. [ Links ]
12. Freeman, G. H. 1973. Statistical methods for the analysis of genotype-environmet interactions. Heredity 31: 339-354. [ Links ]
13. Gálvez, G. E. y F. J. Morales. 1995. Virus transmitidos por áfidos. En: Pastor Corrales, M. A. y H. F. Schwartz (eds.), Problemas de producción del frijol en los trópicos. 2. ed. CIAT, Cali, Colombia, pp. 385-415. [ Links ]
14. Ghaderi, A.; M. W. Adams and A. W. Saettler. 1982. Environmental response patterns in commercial classes of common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 63: 17-22. [ Links ]
15. Hardwick, R. and J. C. Wood. 1972. Regression methods for studying genotype-environment interactions. Heredity 28: 209-222. [ Links ]
16. Lin, C. S. 1982. Grouping genotypes by a cluster method directly related to genotype-environment interaction mean square. Theor. Appl. Genet. 62: 277-280. [ Links ]
17. Mandel, J. 1971. A new analysis of variance model for non-additive data. Technometrics 13: 1-18. [ Links ]
18. Morales, F. J. 1981. Informe sobre el diagnóstico preliminar de la enfermedad del poroto (Phaseolus vulgaris L.) denominada "achaparramiento": posible etiología y recomendaciones. Informe Técnico. INTA Famaillá, R. Argentina. [ Links ]
19. Van Schoonhoven, A. and M. A. Pastor Corrales. 1987. Standard system for the evaluation of bean germplasm. CIAT, Cali, Colombia. [ Links ]
20. Vizgarra, O. N. 1995. TUC 390 y TUC 500: dos nuevas variedades de poroto negro para el NOA. Rev. Ind. y Agríc. de Tucumán 72 (1-2): 81-85. [ Links ]
21. Vizgarra, O. N. 1996. Dos nuevas variedades de poroto negro para el Noroeste Argentino: TUC 390 y TUC 500. Avance Agroind. 16 (65): 3-5. [ Links ]
22. Vizgarra, O. N. 2004. Análisis retrospectivo de las respuestas de los genotipos de porotos (Phaseolus vulgaris L.) a los ambientes de producción del Noroeste Argentino. Tesis doctoral inédita. FAZ- UNT, Tucumán, R. Argentina. [ Links ]
23. Vizgarra, O. N.; L. D. Ploper; M. R. Gálvez; A. C. Ghío y P. C. Cuenca. 2002. Evaluación de nuevas líneas de poroto negro con resistencia a mancha angular. Avance Agroind. 23 (2): 33-35. [ Links ]
24. Vizgarra, O. N.; H. Terán Santofimio y J. R. Toll Vera. 1997. Estabilidad varietal y rendimiento en grano seco de los principales cultivares comerciales de poroto negro (Phaseolus vulgaris L.) en el Noroeste Argentino. Rev. Ind. y Agríc. de Tucumán 74 (1-2): 47-51. [ Links ]
25. Vizgarra, O. N.; H. Terán Santofimio y J. R. Toll Vera. 1998. Caracterización productiva mediante análisis de conglomerados de las principales variedades comerciales de poroto negro (Phaseolus vulgaris L.) en el Noroeste Argentino. Rev. Ind. Agríc. de Tucumán 75 (1-2): 57-61. [ Links ]
26. Voysest Voysest, O. 2000. Mejoramiento genético del frijol (Phaseolus vulgaris L.). Legado de variedades de América Latina 1930-1999. CIAT- PROFRIZA - COSUDE, Cali, Colombia. [ Links ]

