











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. INTRODUCCIÓN
El desarrollo de plantaciones a escala comercial con híbridos interespecíficos de Eucalyptus se realizan hace décadas en el mundo con el fin de aprovechar la complementariedad, la adaptabilidad y la heterosis o vigor híbrido entre parentales selectos en regiones donde las especies puras tienen limitantes bióticas (plagas y enfermedades), abióticas (riesgo de heladas o sequías, restricciones edáficas), para mejorar determinadas propiedades de la madera o incrementar la capacidad rizogénica (Assis y Mafia, 2007; Harwood, 2011). Dichas plantaciones con híbridos han tenido gran notoriedad en regiones tropicales y subtropicales como Brasil, Sudáfrica y el Congo. Menor desarrollo tuvieron en regiones templadas donde las especies mejor adaptadas (principalmente E. globulus, E. nitens) presentaron dificultades para la implementación de un programa de propagación clonal comercial debido al bajo porcentaje de enraizamiento de las especies parentales e híbridos interespecíficos (Potts y Dungey, 2004). En este sentido Assis y Resende (2011) destacan que la mayoría de las casi 5 millones de hectáreas forestadas con eucaliptos en el Brasil fueron realizadas con clones híbridos (en su mayoría E. urophylla x E. grandis). Potts y Dungey (2004) mencionan que existen importantes plantaciones con híbridos de eucalipto en China, Indonesia y Sudáfrica. También hay pequeñas áreas en otros países de Asia (Filipinas, Vietnam, Tailandia y Malasia) y Sudamérica (Chile, Uruguay, Argentina y Paraguay).
En Argentina los primeros antecedentes experimentales de híbridos interespecíficos fueron reportados por Alliani (1990). En la Mesopotamia Argentina los primeros cruzamientos controlados para la selección de clones híbridos de E. grandis con E. camaldulensis, E. tereticornis, E. dunnii y E. globulus fueron generados por Harrand y Schenone (2002). Más recientemente Marcó y Harrand (2012) informaron que el Instituto Nacional de Tecnología Agropecuaria (INTA) posee alrededor de 130 clones híbridos en diferentes etapas de evaluación. Seis de los mismos (3 de E. grandis x E. camaldulensis y 3 de E. grandis x E. tereticornis) ya fueron inscriptos en el Registro Nacional de Cultivares del INASE y desde 2014 están siendo transferidos a los viveros de la región a través de Convenios de Transferencia. Así mismo, en el sector privado, el CIEF y la Empresa Pomera Maderas disponen de clones híbridos para cubrir los requerimientos de sus programas de plantaciones como así también de terceros (E. grandis x E. camaldulensis, E. grandis x E. tereticornis y E. grandis x E. urophylla).
En general, los resultados disponibles en la región mesopotámica muestran que los clones híbridos de E. grandis x E. tereticornis y de E. grandis x E. camaldulensis tienen crecimientos similares o superiores a E. grandis, con valores de forma y sanidad inferiores (Raute et al., 2005; Marcó y Harrand, 2005 y 2012). No obstante, en cuanto a propiedades de la madera la información es muy limitada ya que se refiere a un único clon de E. grandis x E. tereticornis (López et al., 2007; Sanchez Acosta, 2012) no existiendo antecedentes en la literatura local respecto al grado de determinación genético de caracteres de crecimiento o de propiedades ligadas a la calidad de la madera para usos de alto valor a efectos de predecir y contrastar genéticamente su aptitud.
La estimación de los parámetros genéticos y la predicción de los valores de mejora de los materiales a seleccionar tienen una importancia central para la definición de las estrategias de mejoramiento sexual y selección clonal (Resende, 2002; White et al., 2007). En este sentido, a partir de la década del 90 con la progresiva utilización de los modelos mixtos de árboles individuales, donde se estiman los parámetros genéticos utilizando el método de Máxima Verosimilitud Restringida (REML) y se predicen los valores de mejora a través de un modelo BLUP (Mejor Predicción Lineal Insesgada), los Programas de Mejoramiento Genético Forestal dieron un gran salto cualitativo en relación a las metodologías analíticas de selección genética. Estas permiten jerarquizar a todos los genotipos evaluados estrictamente por su mérito genético libre de efectos ambientales (Borralho, 1995; Resende, 2002; White et al., 2007).
Los valores genéticos (efectos genéticos aditivos) o genotípicos (efectos aditivos y no aditivos) son variables aleatorias no observables que deben ser predichas a partir de los valores fenotípicos observados (mediciones cuantitativas o cualitativas realizadas en experimentos de campo o laboratorio). Según Resende (2004) considerar a los efectos de tratamientos (familias o clones) como aleatorios es esencial para el mejoramiento genético y es la única manera de hacer selección genética. Caso contrario la selección es fenotípica, abordaje éste ampliamente discutido por varios autores (Borralho, 1995; Duarte y Vencovsky, 2001; Piepho et al., 2008). Así mismo, al asumir que los efectos genéticos son aleatorios, las pruebas de comparaciones múltiples (Tukey, Scheffé, Duncan o similares) no son aplicables para establecer diferencias genéticas, ya que dichos métodos son derivados de la suposición de efectos fijos de tratamientos y no de variables aleatorias, con lo cual las inferencias obtenidas se refieren a medias fenotípicas y no a medias genéticas o genotípicas (Resende, 2002; White et al., 2007).
Con la aplicación de los modelos mixtos la herramienta clasificatoria para familias, clones o individuos en los ensayos genéticos es el ordenamiento jerárquico basado en la predicción puntual de los valores genéticos o genotípicos predichos generados por el BLUP (Duarte y Vencovsky, 2001). No obstante, Resende (2002) sugiere, en términos de inferencia estadística paramétrica, la conveniencia de complementar la predicción puntual con la predicción de los intervalos de confianza de los valores genéticos predichos.
El objetivo del presente trabajo fue evaluar un grupo de clones híbridos generados por la EEA Concordia dentro del Programa de Mejoramiento Genético de Eucalyptus del INTA a efectos de estimar parámetros genéticos y superioridad/inferioridad esperada respecto a algunos materiales comerciales de .. grandis utilizados en la Mesopotamia Argentina.
2. MATERIALES Y MÉTODOS
a. Material genético
El material evaluado forma parte de un ensayo originalmente constituido por 32 clones híbridos, 4 clones puros de E. grandis y 4 testigos de semilla. La generación de las progenies híbridas (F1) y la posterior selección de los individuos que dieron origen a dichos clones fue descripta por Harrand y Schenone (2002) y Harrand (2005). El grupo de clones híbridos seleccionados para este estudio fueron elegidos por características de crecimiento, rectitud del fuste y sus antecedentes de mayor tolerancia al frío. En total (Tabla 1) se muestrearon 11 clones de E. grandis x E. camaldulensis (GC), 5 clones de E. grandis x E. tereticornis (GT), 1 clon de E. grandis x E. dunnii (GD), 2 clones puros de E. grandis (EG) y un material de semilla mejorada (HSP). Los clones 5-108 (GC) y 5-130 (GT) corresponden a selecciones realizadas en 1992 en el marco del Convenio INTA-CIEF en la región pampeana (Marcó y Harrand, 2005).
b. Métodos de muestreo y evaluación
Al 6º año de edad para cada uno de los 20 materiales a estudiar se seleccionaron 5 ejemplares representativos. Para los clones EG36, GD1 y GC9 solo 4 rametos fueron considerados, con lo cual un total de 97 individuos fueron muestreados. En cada uno de ellos se midió diámetro a 1,30 m de altura (DAP) con cinta diamétrica de 1 mm de precisión, altura total (ALT) con un Vertex® III de 10 cm de precisión y se calculó el volumen individual con corteza (VOLcc) utilizando la ecuación propuesta por Glade y Frield (1988). Posteriormente, dichos ejemplares fueron abatidos e inmediatamente después del corte se elaboraron en cada uno de ellos, a partir de 1,30 m de altura, 2 rollizos consecutivos (el primero de 3,2 m y el segundo de 2,5 m de longitud). Siguiendo la metodología propuesta por López et al. (2009) se estimaron las tensiones de crecimiento a través del índice de Rajado en rollizos (IRr), proporción de corteza (CORT) y se obtuvieron las probetas para medir densidad básica (DB), contracción radial (C.), contracción tangencial (C.), coeficiente de anisotropía (CA) y color del duramen a través de los parámetros colorimétricos L*, a* b* del sistema CIELab76.
c. Análisis genético
La información fue analizada utilizando la metodología de modelos mixtos (modelo lineal mixto univariado) utilizando el programa SELEGEN-REML/BLUP (Versión 2014). De acuerdo al muestreo realizado se asumió un diseño experimental totalmente al azar con parcelas de una sola planta (repeticiones) adoptando el siguiente modelo estadístico: y = Xu + Zg. ., donde y es el vector de las observaciones, u es el escalar relacionado con la media general (asumidos como fijos), .es el vector de los efectos genotípicos (asumidos como aleatorios) y g es el vector de los residuos (asumidos como aleatorios). X y Z corresponden a las respectivas matrices de incidencia. Así mismo, de manera complementaria para el VOLcc se analizaron los mismos materiales motivo de este estudio, ocupando la información del ensayo medido a la misma edad en la que se realizó el muestreo, utilizando un modelo estadístico de bloques al azar con 5 repeticiones y promedios de parcela. Siguiendo los modelos descriptos y considerando en primera instancia solo a los 17 clones híbridos y en un segundo análisis incluyendo a los dos clones puros de E. grandis (19 clones híbridos y puros) para cada propiedad evaluada se estimó la heredabilidad clonal: H2 = σ2c/σ2c + σ2e , donde H2 es la heredabilidad de parcelas individuales en sentido amplio, o sea, de los efectos genotípicos totales, σ2c es la varianza genotípica o clonal y σ2e es la varianza residual.
La significancia de la inclusión de los efectos aleatorios (clones) fue estimada a través del análisis de la desvianza (desviance) y la Prueba de Razón de Verosimilitud (LRT: likelihood ratio test) confrontando con el valor de Chi cuadrado ( χ2
) con 1 grado de libertad. Para estimar la exactitud de la predicción de los valores genotípicos (accuracy) se utilizó el estadístico  ; donde PEV es la variancia del error de predicción y σ2c es la varianza genotípica o clonal.
; donde PEV es la variancia del error de predicción y σ2c es la varianza genotípica o clonal.
Con el fin de obtener un ranking que permita visualizar la superioridad/inferioridad genética de los clones respecto al material mejorado de semilla comercial, en un tercer análisis, además de los 19 clones puros e híbridos se incluyó al testigo HSP. Sumado a ello y tal lo sugerido por Resende (2002) a efectos de predecir las diferencias genéticas entre los materiales estudiados, se calculó el límite superior e inferior de los intervalos de confianza (LSIC y LIIC) de los valores genotípicos predichos (VG) por medio de la siguiente expresión: VG ± t · SEP, donde t es el valor de la distribución de . de Student al 95% de confianza y SEP es el desvío estándar del VG.
3. RESULTADOS Y DISCUSIÓN
En la Figura 1.a se puede observar que la correlación simple entre las medias fenotípicas del VOLcc de los ejemplares recolectados para esta investigación y el promedio correspondiente al ensayo medido a la misma edad, resultó altamente significativa (p<0,01) evidenciando una buena representatividad volumétrica de los materiales muestreados (r = 0,91). En la Figura 1.b se presentan los valores genotípicos predichos (VG) del volumen individual del ensayo y los intervalos de confianza de la predicción genotípica. Allí se destaca que 9 clones híbridos tuvieron una media genotípica (VG) inferior al testigo de semilla mejorada (HSP) y 8 un VG superior entre 2,4 % (GC9) y 18,9 % (GC18) respecto del testigo HSP (Tabla 2). Sin embargo, teniendo en cuenta el solapamiento de los intervalos de confianza, dicha inferioridad/superioridad genética no pudo ser comprobada con un 95 % de confianza (Figura 1.b). Los únicos materiales genéticamente diferentes y con un volumen inferior al testigo de semilla mejorada (HSP) fueron los clones 5-108 (GC) y 5-130 (GT) dado que el LSIC de ambos clones fueron inferiores al LIIC del testigo HSP (Figura 1.b). Estos clones híbridos fueron seleccionados en la década del 90 a través del Convenio INTA-CIEF en un ensayo con semillas híbridas (F1) procedentes de Sudáfrica (Marcó y Harrand, 2005) y su inclusión en el presente análisis prácticamente fue en carácter de referencia. La no diferenciación genética en VOLcc de los demás clones híbridos respecto a los 2 clones de E. grandis al material procedente del Huerto Semillero de Progenies (HSP) es alentadora ya que, esta última fuente semillera se destaca por su alta productividad en la Mesopotamia Argentina. Sumado a ello, el resultado obtenido permite inferir que es posible esperar nuevos clones híbridos con rendimientos superiores a los aquí evaluados al utilizar parentales de E. camaldulensis y E. tereticornis seleccionados genéticamente en orígenes de mayor adaptación a la región mesopotámica.

Figura 1 a) Relación del VOLcc de los materiales muestreados y la evaluación realizada en el ensayo. b) Valores Genotípicos (VG) y límites superior e inferior de los intervalos de confianza (LSIC, LIIC) generados a partir del análisis del ensayo. La línea de puntos indica el LSIC y LIIC del testigo de semilla mejorada. (●HSP, ♢clones de E. grandis, ▂ clones híbridos).
En la Tabla 2 se detallan las medias genotípicas de las propiedades evaluadas en todos los materiales estudiados. En la misma se puede ver que al analizar solo a los 17 clones híbridos la Prueba de Razón de Verosimilitud (LRT) cotejada con el test de Chi cuadrado, detectó diferencias altamente significativas para los efectos genotípicos. Así también, con valores absolutos en general mayores de LRT, al incluir en el análisis a los 2 clones puros de E. grandis. En cuanto a la exactitud selectiva (rĝg) la misma varió entre 0,89 y 0,96 evidenciando alta precisión en las inferencias de las medias genotípicas. Este parámetro según Henderson, 1984 (citado por Resende y Duarte, 2007) es de gran importancia en el contexto de la evaluación genética ya que se refiere a la correlación entre el valor genotípico verdadero y el estimado o predicho a partir de la información proporcionada por el experimento.
Tabla 2 Media genotípica (VG) de los materiales evaluados. Prueba de Razón de Verosimilitud (LRT), heredabilidad clonal (H2) y exactitud selectiva (rĝg) de las propiedades analizadas
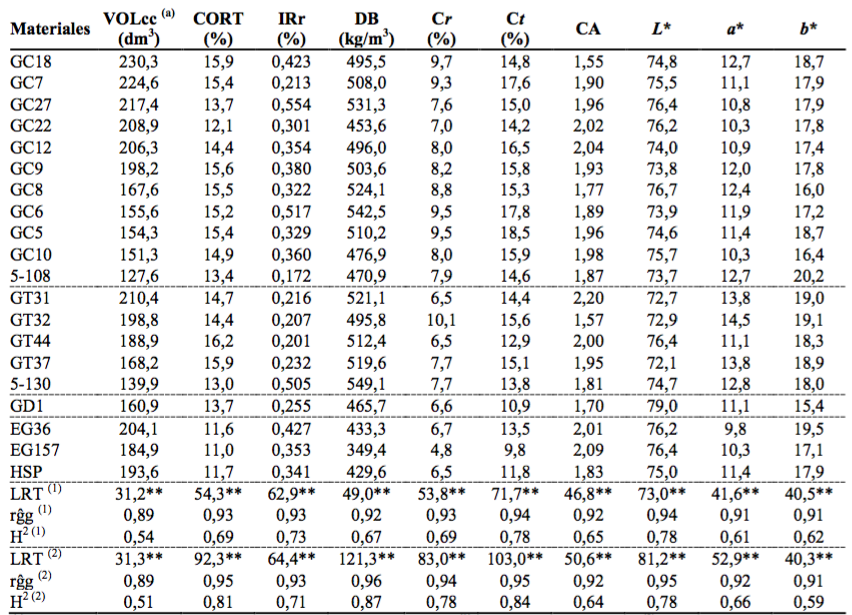
(a) corresponde a las evaluaciones realizadas en el ensayo, (1) solo clones híbridos, (2) clones puros e híbridos, ** significativo al 0,01% por el test de Chi cuadrado.
En ambos análisis las estimaciones para la H2 del VOLcc resultaron moderadas y levemente inferiores a las obtenidas para las demás características evaluadas, pero similares a las encontradas en otros estudios involucrando clones puros de Eucalyptus grandis (Lambeth et al., 1994; López, 2012) como también, clones puros e híbridos de otras especies del género (Gaspar y Borralho, 2004; Varghese et al., 2008; Wu et al., 2011).
La H2 de la proporción de corteza (CORT) al considerar solo a los clones híbridos fue de 0,69 y al incluir a los 2 clones puros de E. grandis fue de 0,81 evidenciando que dicha inclusión incrementó la varianza genotípica producto del menor porcentaje de corteza de E. grandis (Tabla 2). No obstante, dichos coeficientes fueron concordantes con la literatura disponible. En relación a ello, Harrand et al. (2009) al evaluar el porcentaje de corteza en familias de polinización libre de E. grandis reportaron heredabilidades individuales en sentido estricto (h2) entre 0,19 y 0,31 resultando éstas superiores a las h2 correspondientes a los caracteres de crecimiento (DAP, ALT y VOLcc). Por su parte Wei y Borralho (1997) en progenies de E. urophylla detectaron que la h2 del espesor de corteza y la proporción de corteza fue de 0,45 y 0,40 respectivamente. Considerando clones híbridos de E. urophylla con E. grandis, E. tereticornis y E. camaldulensis e híbridos de E. grandis con E. uropylla y E. tereticornis Wu et al. (2011) indicaron que la H2 de la media clonal fluctuó entre 0,77 y 0,88 al 4º año de edad.
Tal como se consigna en la Tabla 2 la proporción de corteza (CORT) de los clones híbridos evidenció medias genotípicas superiores al testigo HSP entre 3,4 % (GC22: 12,1) y 38,5 % (GT44: 16,2) siendo éste un aspecto negativo en cualquier proceso industrial. Sólo los clones híbridos GC22, 5-130, 5-108 y GD1 no se diferenciaron genéticamente con un 95 % de confianza (Figura 2.a). Es evidente que las especies parentales tuvieron un importante efecto en esta característica. En este sentido, Salvioni et al. (2003) determinaron que el porcentaje de corteza para E. grandis, E. dunnii, E. camaldulensis y E. tereticornis varió entre 10,9-12,3 %, 13,6-17,4 %, 18,6-19,5 % y 18,0-21,2 % respectivamente.
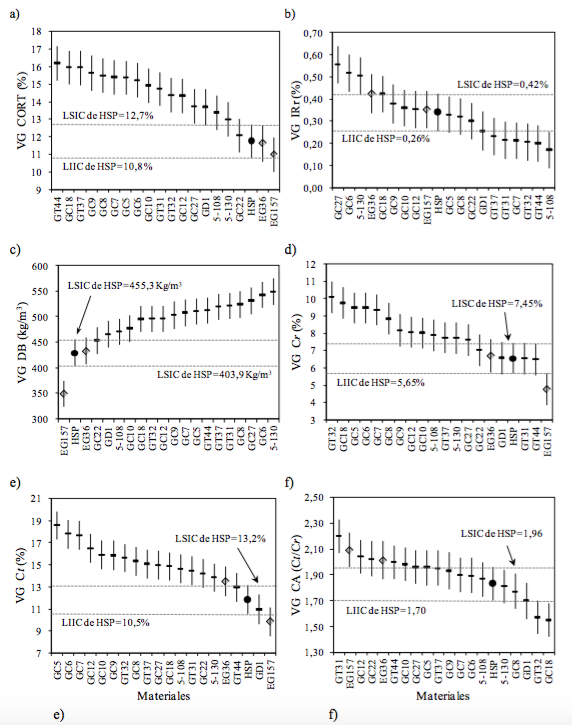
Figura 2 Valores Genotípicos (VG) y límites superior e inferior de los intervalos de confianza (LSIC, LIIC) para proporción de corteza (CORT), índice de rajado en rollizos (IRr); densidad básica (DB), contracción radial (Cr), contracción tangencial (Ct) y coeficiente de anisotropía (CA). La línea de puntos indica el LSIC y LIIC del testigo de semilla mejorada. (: HSP, : clones de E. grandis, : clones híbridos).
La H2 del IRr y de la DB varió entre 0,71-0,73 y 0,67-0,87 respectivamente, siendo éstas coherentes con las reportadas en otras investigaciones que demuestran que ambas propiedades son altamente heredables y factibles de mejorarlas aplicando estrategias convencionales de mejoramiento genético. (Pádua et al., 2004; Varghese et al., 2008; Blackburn et al., 2011; López, 2012; López et al., 2012).
Siguiendo las clases de severidad del IRr sugeridas por López et al. (2009) para E. grandis, se observó que los clones híbridos tuvieron medias genotípicas (VG) de IRr entre Muy Bajas a Medias (Tabla 2). Genéticamente se diferenciaron del testigo HSP por su bajo IRr el clon 5-108 y por su mayor IRr los clones GC6 y GC27 (Figura 2.b). Si bien las evaluaciones fueron realizadas al 6º año de edad es esperable que el comportamiento relativo de los materiales evaluados se mantenga en el tiempo. En este sentido López y López (2013) en clones de E. grandis reportaron un alto coeficiente de repetibilidad edad-edad (r = 0,82) al evaluar el IRr de los mismos clones al 6º y 11º año de edad, verificando alta consistencia en la posición relativa de los genotipos a través de las dos edades de evaluación. No obstante, si bien en clara dependencia de cada genotipo en particular, también comprobaron un incremento promedio entre ambas edades de evaluación del 64% con lo cual, clones con IRr Medio podrían pasar a la clase de IRr Alto, cuestión ésta que advierte sobre a la fuerte presión de selección que debería ejercerse a edades juveniles para minimizar su efecto al turno de cosecha y disminuir el impacto negativo en la producción de madera aserrada.
En relación a la densidad básica de la madera (DB) y debido a las características intrínsecas de las especies utilizadas como padres, todos los clones híbridos exhibieron valores genotípicos (VG) superiores a los 2 clones de E. grandis y al testigo de HSP (Tabla 2) revelando mejor aptitud que E. grandis para usos de mayor resistencia mecánica o para bioenergía. Esta superioridad fue desde 5,6 % (GC22: 453,6 Kg/m3) a 27,8 % (5-130: 549,1 Kg/m3), no pudiendo diferenciarse genéticamente de HSP solamente los clones GC22, GD1, 5-108 y GC10 (Figura 2.c). Dado que la DB de la madera adulta, en promedio, es un 25-30 % mayor que la desarrollada a edades jóvenes, debe esperarse que en edades más avanzadas la DB de los materiales aquí evaluados presenten un incremento de similar magnitud (López y Staffieri, 2003; Cardozo et al., 2013; López y López, 2011). Pese a ello, el ranking genético obtenido no debería sufrir alteraciones significativas ya que la literatura muestra que la correlación genética juvenil-adulto de la DB en eucaliptos generalmente es alta (>0,80) (López, 1993; Osorio et al., 2003; Kien et al., 2008).
En cuanto a la H2 de las contracciones radiales (Cr) y tangenciales (Ct) al considerar solo a los clones híbridos fue de 0,69 y 0,78 respectivamente (Tabla 2). Al incluir a los 2 clones puros de E. grandis dichos coeficientes fueron mayores (H2 Cr = 0,78 y H2 Ct = 0,84). Estas estimaciones coinciden con Botrel et al. (2007) quienes al evaluar clones de E. tereticornis x E. pellita y E. camaldulensis x E. grandis obtuvieron una H2 de 0,79 para la Cr y de 0,80 para la Ct. Por su parte, Rodriguez et al. (2008) en clones de Eucalyptus ssp. también detectaron altas H2 para las Cr, Ct y CA aunque ésta última propiedad tuvo una H2 entre 20 y 23 % menor. En este sentido, tal como se consigna en la Tabla 2, el CA en ambos análisis también evidenció heredabilidades menores a la de las contracciones lineales primarias. No obstante, una H2 de 0,64-0,65 indica que este estimador de la estabilidad dimensional manifiesta un moderado control genético.
A excepción del clon híbrido de E. grandis x E. dunnii (GD1) y los clones GT31 y GT44, cuyos VG fueron similares al testigo HSP, los demás clones híbridos exhibieron contracciones radiales y tangenciales de mayor magnitud (Tabla 2). Cabe aquí mencionar que las medias genotípicas y los límites superior e inferior de los intervalos de confianza del testigo de E. grandis (Figura 2.d y e) son coincidentes con los promedios reportados por Calvo et al. (2007) para muestras de plantaciones ubicadas en el Noreste de Entre Ríos y Corrientes. Dichos autores informaron que la Ct osciló entre 4,45 y 7,10 % y la Ct entre 9,76 y 11,54 %.
En la Figura 2.d puede visualizase que, en cuanto a las Cr, 6 clones híbridos (GC8, GC7, GC6, GC5, GC18 y GT32) se diferenciaron genéticamente del testigo de semilla mejorada (HSP). Por su parte, 13 de los 17 clones híbridos y con un 95% de confianza (Figura 2.e) evidenciaron Ct genéticamente mayores al material de E. grandis utilizado como testigo (HSP). Contracciones radiales y tangenciales de hasta un 37 % superiores a las de E. grandis fueron reportadas por Malan (1993) al evaluar clones híbridos de E. grandis x E. camaldulensis y E. grandis x E. tereticornis al 5º año de edad en diferentes sitios de Sudáfrica.
Considerando los valores genotípicos (VG) del coeficiente de anisotropía (CA) la mayoría de los clones híbridos y según la clasificación de Coronel (1994) se ubicaron en la categoría de estabilidad dimensional normal (Tabla 2). La estimación puntual para el CA del testigo de semilla (HSP) fue de 1,83 siendo éste comparable al valor más bajo obtenido por Calvo et al. (2007) quienes señalan para E. grandis coeficientes de anisotropía entre 1,86 y 2,25. Algunos clones híbridos (GC8, GD1, GT32 y GC18) a pesar de los altos valores de contractibilidad evidenciaron buena estabilidad dimensional con CA entre 3,4 % (GC8: 1,77) y 18,1 % (GC18: 1,55) inferiores al testigo HSP. Otros materiales como los clones híbridos GT31, GC12 y GC22 presentaron CA que califican a su madera como de menor aptitud para varios usos (Coronel, 1994). Estas maderas “nerviosas” o de grandes movimientos y alta densidad, como algunos de los clones híbridos evaluados, según Cardozo et al. (2013) son posibles de utilizarlas en pisos de alto tránsito, mobiliarios de exterior y cubiertas, productos éstos en los que la anisotropía puede ser tratada por diseño. De todas maneras los intervalos de confianza de la predicción genotípica (Figura 2.f) solo permite asegurar que los clones híbridos GT31 y GC18 tuvieron un CA genéticamente diferente al testigo de semilla mejorada de E. grandis.
Respecto al color del duramen, la H2 de los parámetros colorimétricos L*, a* y b* y considerando solo a los clones híbridos fue de 0,78, 0,61 y 0,63 respectivamente (Tabla 2) indicando que entre el 61 y 78 % de la variación fenotípica total no pudo ser explicada por efectos meramente ambientales, siendo dichos porcentajes preponderantemente de origen genético. La inclusión en el análisis de los 2 clones puros de E. grandis prácticamente no produjo cambios en la magnitud de la H2 de los 3 parámetros colorimétricos. Si bien la literatura es escasa respecto al grado de determinación genético del color de la madera, el moderado control genético de esta propiedad es coincidente con lo reportado por Vanclay et al. (2008) para E. dunnii y Sotelo Montes (2008) para Calycophyllum spruceanum. No obstante, desde el punto de vista industrial y considerando usos sólidos de apariencia, al comparar las diferencias del color del durámen de los clones híbridos GC y GT con el testigo de E. grandis (HSP) utilizando la metodología estándar del Sistema CIELab (DE*=[(DL*)2 + (Da*)2 + Db*)2]1/2) los DE* oscilaron entre 0,58 (GC7) y 3,93 (GT32). Al confrontar el único híbrido con E. dunnii (GD1) el DE* fue de 4,73. Estos resultados posicionan a los clones híbridos GC y GT, en menor medida al híbrido GD1, en igualdad de condiciones que E. grandis para usos de apariencia ya que según Melgosa et al. (2001) un observador promedio solo podría distinguir diferencias de color “notables” cuando el DE* es superior a 5 unidades CIELab.
4. CONCLUSIONES
Para todas las características evaluadas en los clones híbridos se detectaron diferencias altamente significativas para los efectos genotípicos y las heredabilidades en sentido amplio oscilaron entre moderadas a altas (0,54-0,78). La exactitud selectiva (rĝg) varió entre 0,89 y 0,96 evidenciando alta precisión en las inferencias de las medias genotípicas.
Al comparar con el material de semilla mejorada de E. grandis, la mayoría de los clones híbridos evidenciaron alta velocidad de crecimiento, elevada proporción de corteza, alta densidad de la madera, bajo nivel de tensiones de crecimiento y una estabilidad dimensional aceptable a pesar de los altos valores de contractibilidad radial y tangencial. En cuanto al color del duramen se constató gran similitud con el testigo de E. grandis posicionando a éstos híbridos en igualdad de condiciones para usos de apariencia.
Dado que el análisis fue realizado en un único sitio las variancias genotípicas podrían estar sobrestimadas, por lo tanto sería conveniente realizar evaluaciones similares en otros sitios y a edades más adultas a efectos de contar con estimaciones más robustas e identificar clones con mayor adaptabilidad, estabilidad y productividad. Sumado a ello, la generación de nuevos clones híbridos utilizando parentales de E. camaldulensis y E. tereticornis seleccionados en orígenes de mayor adaptación a la región mesopotámica, no solo permitirá ampliar las fronteras forestales hacia zonas donde E. grandis presenta restricciones, sino también incrementar la oferta de otros productos maderables.

