











 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntrodugao
As toxinas estão presentes em diferentes animais, plantas e microrganismos com o objetivo de interromper ou influenciar componentes essenciais do processo fisiológico de outros organismos, o que pode trazer vantagens em sua sobrevivência e evolução. Esses venenos podem causar dor, paralisia e até mesmo a morte de suas presas ou predadores. Essas toxinas funcionam siner-gicamente ou individualmente, geralmente com grande potencia (Walker 2020). A toxicidade do veneno geralmente está ligada a poucas toxinas que compóem o veneno. Apesar do nome, muitas toxinas nao sao tóxicas e podem ter aplicapóes terapéuticas (Ferraz et al. 2019).
Os peptídeos de toxinas animais apresentam alta especificidade e poténcia para alvos moleculares específicos. Sao compostos biologicamente ativos que muito estimulam o interesse dos pes-quisadores e podem ser candidatos a fármacos extremamente valiosos. Essas substancias quan-do caracterizados, possibilitam sua compreen-sao funcional, além de promover sua possível aplicabilidade biotecnológica (Boldrini-Franca et al. 2017). Existe uma enorme biblioteca natural inexplorada de compostos bioativos contidos em secrepóes de venenos, pois uma única secrepao pode conter vários compostos diferentes, principalmente proteínas, pequenos peptídeos, mas também contém sais e compostos organicos, como aminoácidos e neurotransmissores (Madio et al. 2017).
A secrepao do veneno de muitos animais pe-ponhentos, como cobras, escorpióes, aranhas, insetos e anfíbios, pode ser aplicada na fabri-capao de agentes e/ou ferramentas para pesquisa básica e aplicada para serem usados em aplicapóes farmacológicas e biotecnológicas (da Silva et al. 2014; Peigneur e Tytgat 2018). As toxinas tém sido uma importante fonte de fer-ramentas moleculares para o desenvolvimento de medicamentos que visam uma variedade de condipóes tais como a dor crónica (Hamad et al. 2018), diabetes (Furman 2012), cáncer (Qiao et al. 2017), acidente vascular cerebral (Chassagnon et al. 2017) e doenpas autoimunes (Tarcha et al. 2017). Entretanto, o mecanismo de como essas toxinas ajudam no tratamento de várias doenpas permanece desconhecido.
O estresse oxidativo e o efeito de antioxidantes sao importantes na avaliapao do estado de saúde/ doenpa. Qualquer desequilíbrio entre a produpao de radicais livres e a defesa antioxidante gera estresse oxidativo que promove ou agrava várias condipóes fisiopatológicas, tais como envelheci-mento, diabetes, cáncer, doenpas de Alzheimer e Parkinson (Valko et al. 2007; Negre-Salvayre et al. 2010; Matsushima et al. 2014; Marrocco et al. 2017).
Tendo em vista que o estresse oxidativo está envolvido na evolupao de muitos distúrbios e que os venenos sao misturas complexas de compostos com funpóes farmacológicas específicas que podem ser um alvo potencial para o desenvolvimento de drogas, revisamos a participapao de toxinas animais em doenpas, como cáncer, Parkinson, Alzheimer, toxicidade hepática e renal e artrite, atuando como possíveis ferramentas terapéuticas contra o estresse oxidativo.
Marcadores do estresse oxidativoA definipao de estresse oxidativo inclui dois mecanismos diferentes, um relacionado ao dano molecular e outro relacionado a interruppao da sinalizapao redox. Assim, o estresse oxidativo pode ser definido como um desequilíbrio entre oxidantes e antioxidantes, que resulta em dano macromolecular e alterapao na sinalizapao redox (Sies e Jones 2017).
Biomoléculas, como proteínas, lipídios e ácidos nucléicos, podem ser irreversivelmente oxidadas por espécies reativas de oxigénio e nitrogénio (ERO/ERN) produzidas sob estresse oxidativo (Roberts et al. 2010). Existem vários sistemas de defesa intracelular para evitar o aumento descontrolado de ERO/ERN. Essas espécies reati-vas sao moléculas químicas heterogéneas que incluem radicais, como óxido nítrico (NO), anion superóxido e radicais hidroxila. As mitocondrias sao a fonte predominante de EROs em todos os tipos de células (Musatov e Robinson 2012). As espécies reativas sao geradas principalmente no nível da cadeia transportadora de elétrons mitocondrial ou também podem ser produzidas enzimaticamente por várias enzimas ligadas a inflamapao, como NADPH oxidase, óxido nítrico sintase (NOS), xantina oxidase, mieloperoxida-se (MPO), lipoxigenase (LOX), e ciclooxigenase (COX) (Forrester et al. 2018).
Um marcador amplamente utilizado para determinar o aumento do estresse oxidativo é a peroxidapao lipídica (Negre-Salvayre et al. 2010). Este marcador pode estar presente em diversos distúrbios como doenpas cardiovasculares, cáncer, doenpas neurodegenerativas e envelhe-cimento (Niki et al. 2005; Forman et al. 2008). Reapóes enzimáticas ou reapóes em cadeia au-tocatalizadas desencadeadas por EROs podem promover a oxidapao de lipídios. Malondialdeído (MDA) é um dos vários produtos finais de baixo peso molecular formado através da decompo-sipao de certos produtos de peroxidapao lipídica primária e secundária (Signorini et al. 2013). MDA constitui as substancias reativas ao ácido tiobarbitúrico (TBARS), que podem reagir com dois equivalentes de ácido tiobarbitúrico dando origem a um complexo de aduto rosa, facilmente medido por um ensaio colorimétrico ou fluori-métrico (Sousa et al. 2017). A paraoxonase-1 (PON-1) tem papel fundamental na defesa da peroxidagao lipídica. PON 1 é uma enzima sérica ligada as lipoproteínas de alta densidade (HDL). A atividade dessa enzima mostrou ser modulada em condigSes de estresse oxidativo, e foi reco-nhecida também como um agente modulador do papel antioxidante e antiinflamatório do HDL (Ferretti et al. 2010).
Enzimas geradoras de EROs e sistemas de defesa antioxidante, que mudam em resposta ao aumento do estresse oxidativo, podem ser usados para avaliar o estado redox do organismo ou o estado saúde/doenga de células específicas. Os sistemas antioxidantes sao compostos por enzimas, como superóxido dismutase (SOD), catalase (CAT) e glutationa peroxidase (GPX), além de moléculas nao enzimáticas, como glutationa, vitaminas A, C e E e diversos antioxidantes presentes em nossa alimentagao (Valko et al. 2007; Sies e Jones 2017). Os antioxidantes atuam nos organismos vivos por meio de diferentes mecanismos. Dentre eles, podem ser citados: a complexagao de íons metálicos, a captura de radicais livres, a decomposigao de peróxidos, a inibigao de enzimas responsáveis pela geragao de espécies reativas de oxigenio e nitrogenio e a modulagao de vias de sinaliza-gao celular (Vasconcelos et al. 2006) (Figura 1).
A dismutagao do radical superóxido em oxigenio e H2O2 é catalisada por uma família de enzimas chamada SOD. Essas enzimas ajudam a controlar a concentragao intracelular de EROs e a manter o fluxo de H2O2 adequado para a regulagao redox da sinalizagao intracelular, em conjunto com CAT e GPx (Zelko et al. 2002). CAT é uma proteína ho-motetramérica que contém quatro itens de ferro e está amplamente localizada nos peroxissomos. É uma das enzimas que catalisam a conversao de H2O2 em água e oxigenio (Vives-Bauza et al. 2007). A glutationa (GSH) também converte H2O2 em água e oxigenio por meio da oxidagao rever-sível em glutationa oxidada (GSSG), pela agao da glutationa peroxidase (GPX). GSSG pode ser reciclado enzimaticamente em GSH pela atividade da glutationa redutase (GR) e pelo poder redutor de NADPH (Bachhawat e Yadav 2018) (Figura 2). A capacidade antioxidante total é definida como a concentragao de oxidantes neutralizados por um litro de fluidos corporais (Apak et al. 2010). Vários ensaios para determinar a capacidade antioxidante total medem sua eficiencia em eliminar ou reduzir radicais livres. Uma das técnicas usadas para detectar a presenga de compostos antioxidantes, é um método baseado na estabilizagao do radical 2,2-difenil-1-picrilhidrazil (DPPH), onde o composto testado fornece um elétron ao radical acarrentando a mudanga de cor na amostra (Mensor et al. 2001). Outro método, que mede o poder redutor dos antioxidantes é por meio da reagao redox com o ferro, conhecido como potencial antioxidante redutor férrico (FRAP) (Benzie e Strain 1996).
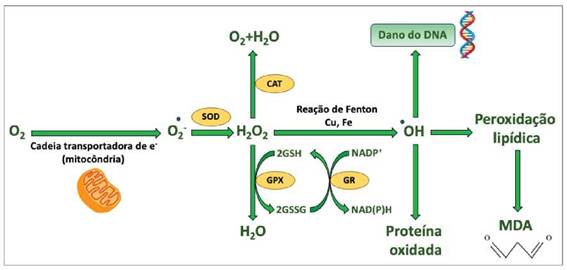
Figura 1: Várias vias de formagao de EROs e sua eliminagao por mecanismos de defesas antioxidantes. Abreviaturas: CAT: catalase, GSH: glutationa reduzida, GSSG: glutationa oxidada, GPx: glutationa peroxidase, GR: glutationa redutase, MDA: malondialdeído, SOD: superóxido dismutase.
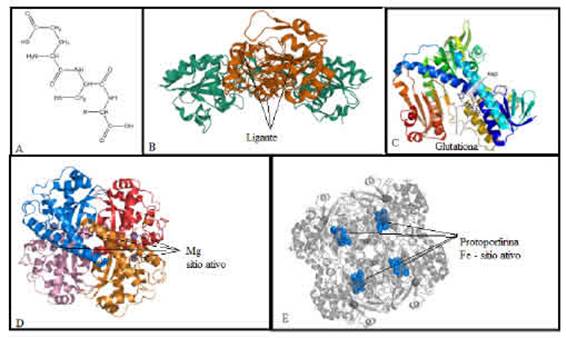
Figura 2: A: Estrutura da glutationa-disponivel em http://www.chemistry.wustl.edu/~edudev/LabTutorials/AirQuality/images/Glutathione.jpg. B: Glutationa peroxidase GPX1 adaptada do Protein Data Bank. C: Glutationa redutase 1GRE - adaptada do Protein Data Bank. D: Superóxido dismutase 2 humana adaptada do Protein Data Bank. E: Catalase adaptada do Protein Data Bank.
Toxinas animais como ferramentas terapéuticas contra o estresse oxidativoVeneno de EscorpiaoO escorpiao é um dos mais antigos artrópodes conhecidos do reino Animalia. Eles existem na Terra há mais de 400 milhoes de anos e sao am-plamente distribuidos em todo o mundo. Dentre as 2000 espécies descritas até hoje, a familia Buthidae é mais amplamente estudada e considerada a mais perigosa para o homem (Hauke e Herzig 2017; Lourengo 2018). Venenos de escorpiao tém sido usados em terapias tradicionais desde a antiguidade em diferentes paises, particularmente na Índia, China, África, Cuba e Espanha (Díaz-García etal. 2013; González e Vallejo 2013; Khusro et al. 2018).
O veneno do escorpiao é uma mistura complexa de neurotoxinas, cardiotoxinas, toxinas he-moliticas, peptideos antimicrobianos, enzimas, lipideos, nucleotideos, mucopolissacarideos e aminas biogénicas. Este veneno é um coquetel de compostos biologicamente ativos, principalmente de natureza proteica, compreendendo desde pequenos peptideos até proteinas de alta massa molecular e de múltiplos dominios, com dobramentos complexos e muitas modificagSes pós-traducionais (Peigneur e Tytgat 2018). Possui uma mistura complexa de aproximadamente 100 a 1000 compostos diferentes. Estes atuam em diversos alvos fisiológicos e bioquimicos do cor-po da presa, causando um desequilibrio em sua homeostase. O veneno de escorpiao representa uma rica fonte de moléculas bioativas e modelos estruturais para o desenvolvimento de novos medicamentos e ferramentas biotecnológicas (Ortiz et al. 2015; Ahmadi et al. 2020).
Muitos estudos sobre os peptideos do veneno do escorpiao enfocam os mecanismos de toxicidade e seus componentes farmacológicos e bioati-vos. A maioria dos peptideos sao neurotoxinas e sao responsáveis pela toxicidade do veneno. Outros peptideos, incluindo peptideos sem ponte de dissulfureto, hialoronidases, fosfolipases e enzimas inibitórias, apresentam potenciais efeitos terapéuticos anticancerigenos, antimicrobianos, analgésicos e antiepilépticos. Peptideos de baixo peso molecular derivados do veneno de escorpiao sao altamente diversos em estruturas primárias e atividades biológicas e tém potencial agao terapéutica a ser explorada (Uzair et al. 2018; Ghosh et al. 2019; Ahmadi et al. 2020).
Nadjia e Fatima (2015), estudaram o nivel de antioxidantes e marcadores de estresse oxidativo em células tumorais hepáticas induzidas por mi-cotoxina Fumonisina B1 (FB1) após tratamento com veneno de escorpiao Androctonus australis hector e sua frapao nao tóxica. Os níveis de NO e MDA aumentaram significativamente com FBI, no entanto, componentes do veneno foram ca-pazes de diminuir os níveis desses marcadores. Os níveis de GSH também foram medidos, mas o veneno nao alterou seus níveis. O mecanismo pelo qual o veneno foi capaz de alterar os marcadores de estresse oxidativo nao está claro, esse efeito necessita ser elucidado. Os autores tam-bém estudaram a atividade da catalase e encon-traram uma redupao nesta atividade promovida pelo veneno e sua frapao nao tóxica, sugerindo uma redupao na produpao de EROs, causando uma melhora no balanpo redox.
O veneno do escorpiao Buthus martensii Karsch (BmK) tem sido usado na medicina tradicional chinesa para tratar inúmeras condipoes por mi-lhares de anos. A presenpa de peptídeos bio-logicamente ativos no veneno desse escorpiao tem gerado grande interesse em possíveis alvos terapéuticos, como anticancer, antimicrobiano, analgésico, antiepilético e que atuam no sistema cardiovascular ou cerebrovascular (Li etal. 2019). Na década passada, dezenas de diferentes peptídeos BmK foram identificados, como o PeSv (Extrato de Polipeptídeo de Veneno de Escorpiao) que induz a inibipao do crescimento e apoptose de células humanas de cáncer de próstata (Zhang et al. 2009), o BmKn2 que possui atividade antimicrobiana (Zeng et al. 2012) e o peptídeo ANEP que apresenta atividade analgésica e também também mostrou atividade antiepiléptica em diferentes modelos animais (Zhang et al. 2001; Song et al. 2017).
Yin et al. (2014), investigando o efeito neuropro-tetor do veneno de escorpiao BmK na doenpa de Parkinson, conduziram um estudo com o peptí-deo resistente ao calor do veneno de escorpiao (SVHRP). Eles avaliaram se este poderia atenuar o estresse oxidativo nos neurónios do cérebro em um modelo de estudo do Parkinson em estágios iniciais. Este peptídeo reverteu significativamente os níveis aumentados de MDA e aumentou a atividade de SOD nas mitocondrias de ratos nos estágios iniciais do Parkinson. A atividade antioxidante total e a capacidade de inibir o radical livre hidroxila foram aumentadas pela SVHRP.
Zhang et al. (2016) avaliaram os efeitos protetores do peptídeo SVHRP do veneno de BmK em um modelo para o estudo da doenpa de Alzheimer. O estudo tentou elucidar alguns dos mecanismos envolvidos nestes efeitos em Caenorhabditis ele-gans. Devido a muitos estudos que associam o estresse oxidativo e a toxicidade da doenpa de Alzheimer, esses autores investigaram, in vivo, a atividade dos EROs nessa linhagem transgénica. A SVHRP atenuou os níveis de EROs nas células de uma maneira dose dependente. Este resultado sugere que a paralisia tardia observada neste modelo tratado com SVHRP pode estar relacionada, em parte, a diminuipao de EROs. Também existe a possibilidade de que a SVHRP tenha efeitos de eliminapao diretos nos EROs, uma vez que as propriedades antioxidantes da SVHRP também foram observadas em outro modelo experimental (Yin et al. 2014).
Os venenos de escorpiao podem ser classifi-cados em peptídeos com ligapoes dissulfeto e peptídeos sem ligapoes dissulfeto (PSPDs) (Zeng et al. 2005). Peptídeos aniónicos de escorpioes correspondem a PSPDs ricos em ácido aspártico e/ou ácido glutamico em sua sequéncia primária, estando presentes nas famílias Buthidae e nao Buthidae (He et al. 2013). Peptídeos anionicos sao abundantes no veneno do escorpiao Tityus stigmurus (espécie prevalente no nordeste do Brasil) (Melo et al. 2016).
Melo et al. (2016), avaliaram a atividade biológica de um peptídeo aniónico presente no veneno do escorpiao Tityus stigmurus, denominado TanP. Para avaliar o possível potencial antioxidante do TanP, foi realizado o teste de sequestro do radical DPPH. Os autores observaram que esse peptídeo apresentou percentual de redupao do radical de aproximadamente 32%. O ácido as-córbico, usado como controle positivo, reduziu o DPPH em aproximadamente 26%. Sugerindo, preliminarmente, que TanP possui uma potente capacidade de sequestro de radicais livres, estando esta condipao relacionada a uma provável atividade antioxidante do peptídeo.
Os venenos animais contém uma variedade de toxinas altamente seletivas e potentes. Em particular, vários desses componentes do veneno (por exemplo, peptídeos potenciadores da bra-dicinina, sarafotoxinas e peptídeos natriuréticos) tém efeitos profundos no sistema cardiovascular (Hodgson e Isbister 2009). A bradicinina e seus peptídeos relacionados sao amplamente distri-buídos em animais peponhentos, incluindo es-corpioes, serpentes e venenos de águas-vivas (Camargo et al. 2012). Foi demonstrado que o veneno do escorpiao egípcio, Buthus occitanus, contém uma frapao de peptídeo com atividade potenciadora da bradicinina (El-Saadani 2004). Bekheet et al. (2011), conduziram um estudo para investigar se o tratamento com fator de potencia-lizagao da bradicinina (BPF) isolado do veneno do escorpiao Buthus occitanus oferece efeitos benéficos na reversao do estresse oxidativo in-duzido pelo cádmio no fígado e rim de ratos. Eles descobriram que a exposigao ao estresse oxidativo causou um aumento significativo no nivel de MDA e uma diminuigao significativa ou inibigao nas atividades de GSH, SOD e CAT. Porém, quando o cádmio foi adicionado com BPF, induziu uma diminuigao significativa no nível de MDA e um aumento nas atividades de GSH, SOD e CAT. Bekheet et al. (2013), investigaram os efeitos protetores do BPF isolado do veneno de escor-piao, Buthus occitanus, no modelo de rato com hepatotoxicidade e nefroxicidade induzidas por gentamicina. Ratos induzidos por gentamicina mostraram um aumento no MDA sérico, que indica um aumento na geragao de radicais livres. Como o BPF impediu o aumento do MDA sérico, parece que esses resultados estao relacionados a propriedade antioxidante do BPF. Além disso, o BPF aumentou as atividades das enzimas antioxidantes SOD, CAT e GPx em animais com hepatotoxicidade induzida por gentamicina. Isso sugere que a redugao do estresse oxidativo neste cenário desempenha um papel no mecanismo de seu efeito hepatoprotetor.
Salman et al. (2016), investigaram o efeito do BPF, do escorpiao Buthus occitanus, na toxici-dade hepática e renal induzida pelo cloreto de mercúrio em ratos. O nivel de MDA aumentou significativamente e diminuiu as atividades de SOD, CAT e GSH-Px em tecidos de fígado e rins de ratos tratados com HgCl2. Estas alteragóes foram revertidas com a administragao do BPF, o que pode sugerir uma redugao na produgao de radicais livres.
Hasan et al. (2020), avaliaram se o BPF isolado do veneno de escorpiao Leiurus quinquestriatus poderia atuar como um modulador natural no dano cardíaco induzido por radiagao. Eles ob-servaram uma diminuigao significativa nos níveis de GSH e FRAP e um aumento significativo no nível de MDA após a exposigao a radiagao. BPF normalizou os níveis de GSH e FRAP e reduziu os níveis de MDA. Esses achados podem apontar para uma redugao na produgao de radicais livres promovida pela administragao de BPF.
Secregóes de anuroOs anfíbios, em sua evolugao, desenvolvem segmentos cutáneos com compostos bioativos que atuam como mecanismos de defesa contra predadores e microrganismos (Meng et al. 2016). Far-macologicamente, essas substancias, presentes em as glándulas cutáneas e parotoideas, podem ser neurotóxicas, cardiotóxicas, hemotóxicas ou miotóxicas e causar efeitos anestésicos, hipo-tensores e/ou hipertensivos (Sakate e Oliveira 2000). Componentes das glándulas dos sapos sao usados para tratar vários tipos de cáncer, como de próstata, fígado e leucemia (Yeh et al. 2003; Gomes et al. 2011) e também usados como agentes terapéuticos na medicina tradicional chinesa, bem como em outros países asiáticos (Chow et al. 2003).
As principais toxinas da secregao de sapos sao classificadas em dois grupos: derivados de es-teróides e compostos básicos. No primeiro estao os bufadienolidos e bufotoxinas; no segundo, aminas biogénicas e bufoteninas. Os derivados esteróides sao responsáveis pelos efeitos car-diotóxicos e os compostos básicos atuam no sistema nervoso autónomo simpático e no sistema nervoso central (Cao et al. 2019).
Baldo et al. (2016), avaliaram a capacidade da fragao de baixo peso molecular da secregao do sapo Rhinella schneideri denominada VR em reduzir espécies reativas de oxigénio em mitocóndrias isoladas de fígado de rato. A fragao VR apresentou efeito protetor contra a produgao de EROs. Essa característica protetora é muito relevante, uma vez que a VR demonstrou nao causar danos (ci-totoxicidade) nesta organela e, portanto, exercer apenas o efeito benéfico de protegao contra o estresse oxidativo. Para tentar elucidar por quais mecanismos ocorre essa protegao, Baldo et al. realizaram testes para avaliar os efeitos do VR sobre a oxidagao de NAD(P)H e também para avaliar seus efeitos sobre os níveis de glutationa reduzida e glutationa oxidada. Os dados obtidos no estudo indicam que o VR nao interferiu no processo de oxidagao do NAD(P)H e nenhuma protegao ocorreu contra a oxidagao da glutationa. Portanto, o VR nao foi capaz de interferir neste sistema antioxidante nas concentragóes utilizadas, o que sugere nao ser esse o processo pelo qual o VR é capaz de diminuir a formagao de EROs. Assim, o mecanismo permanece desconhecido.
Veneno de serpenteO veneno de serpente é uma secregao glandular que as mesmas usam para imobilizar e digerir suas presas. Também é usado como uma ferramenta defensiva e de sobrevivéncia (Waheed et al. 2017). Essa mistura letal é composta por aminoácidos, ácidos nucléicos, carboidratos, lipídios, proteínas e peptídeos. O avango tecnológico e o contínuo interesse em toxinas tém apoiado a descoberta de um grande número de peptídeos de veneno de serpentes. Eles podem ser usados diretamente ou como compostos principáis para a descoberta de drogas e pesquisas de desenvolvimento de fármacos, pois sao fáceis de sintetizar e menos propensas a induzir uma resposta imune (Munawar et al. 2018). Os peptídeos do veneno de serpentes sao classificados em familias distintas com base nas semelhangas estruturais e funcionais na organizado dessas moléculas.
Querobino et al. (2018 e 2019), estudaram a fragao de baixo peso molecular do veneno da serpente Bothropsjararaca. Esta fragáo contém peptídeos ricos em prolina (Bj-PROs), que sao conhecidos como BPPs. Eles tem um efeito neuroprotetor contra o estresse oxidativo induzido por H2O2. Os autores demonstraram que Bj-PRO-5a é um peptídeo neuroprotetor que diminui a peroxida-gáo lipídica, a produgáo de EROs e o aumento dos níveis de NO causados pela exposigáo ao H2O2 (Querobino et al. 2018). Querobino et al. (2019), verificaram que os peptídeos Bj-PRO-5a e Bj-PRO-7a apresentaram efeito neuroprotetor contra o estresse oxidativo induzido por H2O2, reduzindo o dano celular e a morte das células SH-SY5Y. Além disso, a capacidade antioxidante foi medida usando o método de eliminagáo de radicais livres DPPH. Entretanto, esta atividade nao foi detectada pelos peptídeos Bj-PRO-5a e Bj-PRO-7a. Este resultado sugere que esses peptídeos nao tem propriedades de eliminagáo direta de radicais livres. O mecanismo de agao sobre o estresse oxidativo ainda precisa ser esclarecido.
Veneno de AbelhaAs toxinas de insetos tem uma grande variedade de produtos químicos, como pequenas moléculas, poliaminas e toxinas peptídicas (Kachel et al. 2018). O veneno de abelha é composto por vários peptídeos ativos, incluindo melitina, fosfolipase A2, apamina, adolapina e peptídeo degranulador de mastócitos (Eiseman et al. 1982). Apamina, fosfolipase A2 e melitina tem efeito neuroprotetor (Armugam et al. 2009; Alvarez-Fischer et al. 2013) e a adolapina tem um efeito antiinflama-tório (Koburova et al. 1985). O veneno de abelha tem sido usado na medicina oriental para tratar algumas doengas neurodegenerativas (Cho et al. 2012) e doengas relacionadas a imunidade, como artrite reumatóide (Kwon et al. 2002). Ele também aumenta a ativagao da via de sinalizagao apoptótica em linhas de células de cáncer de mama e pulmao (Chu et al. 2007; Ip et al. 2008). Khalil et al. (2015) desenvolveram um estudo para avaliar o efeito neuroprotetor da terapia com acupuntura com veneno de abelha (BVA) contra o estresse oxidativo induzido por rotenona, neuroin-flamagao e apoptose no modelo de camundongo com doenga de Parkinson. Os autores encontra-ram um aumento significativo nos níveis de MDA no cérebro de animais tratados com rotetona. O uso de rotetona com BVA diminuiu esses níveis aos valores encontrados para o grupo controle. A atividade de PON1 e os níveis de GSH no cérebro diminuíram significativamente no grupo que recebeu rotetona, porém o tratamento com BVA reverteu estes efeitos. Esse efeito também foi descrito no estudo de Choi et al. (2009), em que o BVA protegeu os neuronios dopaminérgi-cos contra o dano oxidativo em camundongos intoxicados por MPTP. Esses achados sugerem a terapia com BVA pode ser uma nova aborda-gem no tratamento da doenga de Parkinson ou uma terapia adjuvante com outros tratamentos. Badr et al. (2016), investigaram o impacto do veneno de abelha na cicatrizagao de feridas promovida pelo diabetes em camundongos. Este estudo mostrou que o tecido de camundongos diabéticos tratados com veneno de abelha apre-sentou níveis mais baixos de EROs, peróxido de hidrogenio e NO, o que pode sugerir uma possível atividade antioxidante desse veneno.
Sobral et al. (2016), também investigaram cinco amostras de veneno de abelha obtidas de Apis mellifera iberiensis do Nordeste de Portugal, caracterizadas quimicamente e avaliadas quanto ás suas propriedades antioxidantes, anti-inflama-tórias e citotóxicas. O veneno de abelha provou ter a maior atividade de eliminagao de radicais livres, poder redutor, inibigao da peroxidagao lipídica e também a maior capacidade de inibir a produgao de NO. Todas as amostras revelaram atividade antioxidante, mas sem relagao direta com qualquer um dos componentes químicos individuais identificados.
Mohamed et al. (2019), examinaram o efeito do veneno de abelha na ulceragao aguda induzida por ácido acetilsalicílico (AAS) em ratos. O grupo tratado por via oral com AAS apresentou eleva-goes marcantes nos níveis de MDA e diminui-gSes significativas na atividade gástrica de GSH e SOD. No entanto, o grupo tratado com AAS e veneno de abelha mostrou redugoes marcantes nos níveis de MDA e restauragao significativa da atividade de GSH e SOD. Esses achados indicam que o efeito gastroprotetor do veneno de abelha contra a ulceragao induzida pelo AAS em ratos é mediado por suas propriedades antioxidantes. Kocyigit et al. (2019), avaliaram os efeitos anti-inflamatórios, antioxidantes, antigenotóxicos e imunomoduladores do veneno de abelha melífera (HBV) na artrite induzida em ratos. Eles verificaram que o nível do estado oxidante total do plasma e o índice de estresse oxidativo foram significativamente maiores e os níveis do estado de antioxidante total do plasma foram menores em animais com artrite do que nos saudáveis. Tanto o status oxidante total quanto o índice de estresse oxidativo diminuíram significativamente após a terapia com baixa dose de HBV, enquanto os níveis plasmáticos de status antioxidante total aumentaram. No entanto, o nível do status oxidante total e o índice de estresse oxidativo comegaram a aumentar com o aumento das doses de HBV. Eles concluíram que o tratamento com HBV tem efeitos antioxidantes em ratos com artrite reumatóide. Entretanto, a dose administrada de HBV é fundamental para obter os melhores resultados quando utilizada como agente terapéutico e mais trabalhos sao neces-sários para otimizar a dose de tratamento do HBV e esclarecer seu mecanismo de agao.
Outras espéciesAlgumas espécies de formigas podem produzir veneno com até 75 componentes diferentes (Hof-fman 2010). O género Pachycondyla é encontrado principalmente em regioes tropicais e subtropicais e inclui a formiga Samsum (P. sennaarensis) (Klotz et al. 2005). Existem estudos sobre o potencial terapéutico do veneno P. sennaarensis, com significativo efeito antitumoral nas células do cáncer de mama (Badr et al. 2012) e propriedades an-tiinflamatórias, analgésicas e melhora da fungao dos sistemas imunológico e hepático (Kou et al. 2005). Ebaid et al. (2014), avaliaram se o veneno da formiga Samsum (SAV) poderia influenciar os efeitos da exposigao ao tetracloreto de carbono no status oxidativo e na fungao renal. Os resultados encontrados indicaram melhora da atividade de SOD e CAT e do nível de GSH devido ao SAV, embora o efeito tenha sido significativo apenas em doses mais elevadas. Mais investigagées sao necessárias, portanto, para descobrir a dose mais eficiente de SAV em relagao aos marcadores de estresse oxidativo.
A Vespa, Velutina nigrithorax, é uma espécie exo-tica da Ásia que foi acidentalmente introduzida na Europa (Monceau et al. 2014). Geralmente, o veneno da vespa compreende vários compostos bioativos potenciais: compostos de baixo peso molecular (bioaminas, como histamina, tiramina e catecolamina (Russell 1988) ou fermonios, tais como cetonas alifáticas e álcoois (Ya-Nan et al. 2017), peptídeos (ocupando até 70% do veneno seco, sendo os peptídeos mais populares o mastoparan e as cininas de vespa) e proteínas (neurotoxinas, proteases, fosfolipases) (Russell 1988). A propriedade antioxidante e os compostos responsáveis por esta propriedade do veneno de V. velutina tém sido estudados em Le et al. (2020). A fim de confirmar o envolvimento dessas moléculas no mecanismo antioxidante, os compostos identificados foram testados em ensaios DPPH e EROs em células HaCaT expostas ao UVB. Eles demonstraram que esse veneno apresentou ca-pacidade de inibigao de EROs intracelulares em baixa concentragao sem citotoxicidade. Também identificou a serotonina como o principal composto ativo responsável pela atividade de eliminagao de radicais livres neste veneno.
Conclusoes e perspectivas futurasOs venenos tém sido uma fonte importante de ferramentas moleculares para o desenvolvi-mento de drogas que podem ser aplicadas em variadas condigoes. A literatura científica revisada aqui mostra vários exemplos de toxinas de venenos animais promissores que podem se tornar candidatas potenciais para o futuro tratamento de doengas com elevado estresse oxidativo. E, se os dados observados in vitro e em modelos experimentais se traduzirem bem para o ambiente clínico, pode realmente haver uma grande promessa na exploragao os benefí-cios das toxinas dos venenos de animais. Muitos venenos sao capazes de alterar marcadores do estresse oxidativo, porém investigagoes futuras sao necessárias para descobrir a dose mais eficiente em relagao a estes marcadores, quais componentes sao responsáveis por este efeito e o mecanismo de como essas toxinas atuam na regulagao do balango redox. Assim, estudos posteriores, em diversos modelos experimentais, sao importantes para esclarecer a relagao entre estresse oxidativo e toxinas de venenos animais.
Agradecimentos.Este estudo teve apoio de bolsas de iniciagao científica VIC/UFJF.

