










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkLatin American journal of sedimentology and basin analysis
versão On-line ISSN 1851-4979
Lat. Am. j. sedimentol. basin anal. vol.21 no.2 La Plata dez. 2014
VOLUMEN ESPECIAL
Historia ambiental de los lagos someros de la llanura Pampeana (Argentina) desde el Holoceno medio: inferencias paleoclimáticas
Silvina Stutz 1, Marcela Sandra Tonello 1, María de los Ángeles González Sagrario 2, Diego Navarro 3 y Sonia L. Fontana 4
1 Laboratorio de Paleoecología y Palinología y Ecología y Paleoecología de Ambientes Acuáticos Continentales, IIMyC, CONICET-UNMdP. Funes 3250 (7600) Mar del Plata, Argentina. smstutz@mdp.edu.ar; mtonello@mdp.edu.ar
2 Ecología y Paleoecología de Ambientes Acuáticos Continentales, IIMyC, CONICET-UNMdP. Juan B. Justo 2550 (7600) Mar del Plata, Argentina. gonsagra@mdp.edu.ar
3 IIMyC, CONICET-UNMdP. Peña 4046 1º nivel (7600) Mar del Plata, Argentina. dnavarro@conicet.gov.argamil.com
4 Department of Palynology and Climate Dynamics, Albrecht-von-Haller-Institute for Plant Sciences, Universidad de Göttingen. UntereKarspüle 2, 37073 Göttingen, Alemania. School of Geography, Archaeology and Palaeoecology, Queen's University Belfast, UK. sonia.fontana@biologie.uni-goettingen.dem
Recibido 15 de noviembre de 2014
Aceptado 1 de julio de 2015
RESUMEN
En este trabajo se presenta un estudio paleolimnológico basado en el análisis de múltiples indicadores en múltiples sitios con el objetivo de reconstruir la historia evolutiva de cinco lagos someros de la llanura Pampeana (Argentina) desde el Holoceno medio, y así realizar inferencias paleoclimáticas para esta región. Los cinco lagos estudiados respondieron sincrónicamente y mostraron el mismo patrón de evolución paleoambiental durante el Holoceno medio y tardío. Entre los ca. 7.000 y 700-500 años cal AP, se evidencia una primera fase clara, dominada por carofitas del género Chara, que cambia a los ca. 700-500 años cal AP hacia una fase turbia dominada por fitoplancton y con una importante presencia de carofitas sumergidas del grupo de las angiospermas, que indican fases claras que alternan con el estado turbio general. En la vegetación circundante, se evidencia un cambio desde una comunidad halófita que indica condiciones salobres e inestables hacia la comunidad de macrófitas que caracterizan en la actualidad a estos lagos, por lo cual sugiere ambientes con mayor estabilidad. Durante el Holoceno medio y por largos períodos (alrededor de 6.500 años) la estabilidad del paisaje se mantuvo debido a las condiciones climáticas más secas que las actuales, en las que predominaron fluctuaciones de sequías e inundaciones y/o condiciones de alta evaporación. Estas fluctuaciones son las que mantuvieron a las comunidades tanto acuáticas como terrestres. A partir de ca. 2.000 años cal AP el incremento de la diversidad de macrófitas sumergidas (carofitas y angiospermas) indica mayor aporte de nutrientes y materia orgánica, causado por un aumento en la intensidad y/o duración de las precipitaciones. A partir de este momento comienza un cambio gradual que se efectiviza a los ca. 700-500 años cal AP, con el establecimiento de las comunidades de macrófitas emergentes, flotantes y sumergidas, características de los sistemas actuales.
Palabras clave: Cambio de estados; Estado claro; Estado turbio; Macrófitas sumergidas; Cambio climático; Llanura Pampeana.
EXTENDED ABSTRACT
Environmental history of pampa plain shallow lakes (Argentina) since middle Holocene: Paleoclimatic inferences
High-resolution paleolimnological studies based on multi-proxy analysis constitute an important tool to reconstruct the evolution of aquatic systems as well as evaluating their responses to natural and/or anthropogenic forcing factors (Lotter, 2003; Birks and Birks, 2006). While individual palaeoecological studies reveal local developments, general patterns often only emerge when information from several sites is combined together. Numerous shallow lakes occur throughout the Pampa plain of Argentina, between 33° to 39°S and 57° to 66W. These lakes present two alternative states of equilibrium (Scheffer and Jeppesen, 2007; Scheffer and van Ness, 2007). Some of them are turbid lakes due to the high amount of algae, while others are clear macrophytedominated lakes. A third type of lake that can be recognized within the region is inorganic-turbid lakes, in which turbidity is caused by high amount of suspended inorganic material (Quirós et al., 2002; Allende et al., 2009). These lakes are characterized by low productivity, with scarce phytoplankton and macrophytes. Most of the lakes developed on deflation basins that originated during the late Pleistocene by the prevailing westerly winds. Often, the lakes are associated to lunettes, fixed palaeodunes that developed at the shore, on the windward side of the basins (Tricart, 1973; Zárate and Tripaldi, 2012). During the Holocene, the basins became areas of groundwater discharge and surface water accumulation gradually filling with sediments. Today, the lakes are nutrient-rich, eutrophic to hypereutrophic, and polymictic, too shallow to develop thermal stratification. Water depth and salinity are highly variable. During summer and/or during episodic droughts the lakes suffer significant reduction in water volume, and thus strong fluctuations in their water levels (Quirós and Drago, 1999; Sosnovsky and Quirós, 2006). These aquatic systems are able to support an abundance of macrophytes and phytoplankton, which have proven to leave an exceptional fossil record of environmental changes (Stutz et al. 2002, 2006, 2010, 2012; Fontana, 2005). For more than a decade the authors have been investigating these aquatic systems in order to reconstruct their evolutionary history and the regional environments with the main goal of inferring the past climatic conditions. In order to achieve this, multi-proxy analysis of diverse biological indicators were carried out in several lakes of the south-east region of the Pampa plain. Pollen, non-pollen palynomorphs and plant macrofossil remains and associated fauna were analysed in sediment sequences from lake Hinojales- San Leoncio (37º23'S; 57º23'W) and lake Tobares (37°30'S; 57°28'W). These new results were then combined with former investigations from lakes Lonkoy (37º12'S; 57º25'W), Nahuel Rucá (37º37'S; 57º26'W) and Hinojales (37º34'S; 57º27'W) (Fig. 1). The studied lakes range in suface from 200 to 300 ha and a water depth of about 1 m. The regional vegetation is characterized by temperate subhumid grasslands, knowns as pampas. The modern landscape is strongly influenced by human activities since the establishment of European settlements in the XVI century. The land has been intensively used, in particular for grassing, as well as for agriculture. Native trees are absent in the vegetation, except for Celtis ehrenbergiana, a deciduous tree known as tala, which occurs on Pleistocene lunette dunes and Holocene ridges of shell debris. The aquatic plant communities of the studied sites are characterized by several species of emergent, free-floating leaved and submerged macrophytes. The lakes are surrounded by a ring of Schoenoplectus californicus, among which other emergent macrophytes occur: e.g. Zizaniopsis bonariensis, Typha latifolia, Hydrocotyle bonariensis, H. ranunculoides, Alternanthera philloxeroides, Solanum glaucophyllum, Bacopa monnieri, Polygonum punctatum, Ranunculus apiifolius, Triglochin striata, and Phyla canescens. Near the shore, in sheltered areas, free floating plants like Ricciocarpus natans, Azolla filiculoides, Limnobium laevigatum, Lemna valdiviana, Wolffia brasiliensis, Wolffiella lingulata and W. oblonga form a dense carpet. Diverse submerged macrophytes occur when the water tends to be clear, so the light available is enough for their development: e.g. Myriophyllum elatinoides, Ceratophyllum demersum, Potamogeton pectinatus, and Zannichellia palustris together with diverse Charophytes. The studied sediment records were recovered with different samplers: Dacknovsky (Hinojales), vibracorer (Nahuel Rucá, Hinojales-SL and Tobares) and Livingston-type corer (Lonkoy). The chronology of the records is based on AMS radiocarbon age determinations on terrestrial plant remains, where possible (Table 1). Gastropod shells of Heleobia parchappii have also been used for dating, since previous studies have yielded reliable results (Fontana, 2005, 2007). Age-depth models (Fig. 2) are constructed using CLAM 2.2 (Blaauw, 2010) with the Southern Hemisphere calibration curve, SHCal13 (Hogg et al., 2013). The studied records, spanning the time since the middle Holocene, document similar evolutionary pattern. Changes in the different proxies are consistent, showing similar trends in the evolution of the aquatic ecosystems (Figs. 3-7). At the beginning of the records a clear phase characterized the water bodies, dominated by the green algae Chara, the dinoflagellate Peridinium and aquatic plants such as Myriophyllum, Ruppia and/or Potamogeton. At ca. 2,000 cal yr BP the diversity of submerged macrophyte communities increased, indicating clear water lakes with increasing nutrient content. Towards ca. 700-500 cal yr BP the lakes switched to a turbid phase dominated by phytoplankton together with water fleas and flatworms. Among the phytoplankton the main constitutes are: Pediastrum, Scenedesmus and Tetraedron species of green algae Desmidiaceae and the cyanobacterium Gloeotrichia. On land, halophyte plant communities, dominated by Chenopodiaceae, surrounded the water bodies during the first clear-water phase, suggesting instable periods with brackish conditions. Towards the present, the surroundings of the lakes are characterized by dominance of Cyperaceae together with Bacopa, Ranunculus, Polygonum, Typha and Apiaceae. This vegetation is characteristic of more stable environmental conditions. During the middle Holocene and part of the late Holocene, brackish-shallow lakes with clear water phases characterised the landscape of the south-east pampean plain. After ca. 2,000 cal. yr BP, water run-off and/or wind action increased, incorporating organic matter from the surroundings into the basins and initiating a gradual change towards the next phase. This dynamics could be associated to periods of pronounced drought followed by floods. A marked seasonality in the annual precipitation regimen would explain this dynamics indicating a regionally instable environment with dryer climatic conditions than present (Zárate et al., 1998; Zárate, 2005; Vilanova et al., 2010; Laprida et al., 2014). The change to turbid conditions at around 700-500 cal yr BP was probably caused by a significant input of nutrients into the basin, impeding the development of submerged aquatic plants, characteristic of the clear phase, which in turn favoured the expansion of phytoplankton. However, the presence of submerged macrophytes that tolerate some degree of turbidity like Miryophyllum, Ceratophyllum and Potamogeton would suggest that during some periods the level of turbidity did not reach critical values. The presence of submerged flowering plants may also suggest alternate periods of clear phases within a general turbid state. The synchronous change to a turbid face in all studied sites suggests a climatic regional trigger like an increase of precipitation, with a more stable seasonal regime. Similar values of precipitation compared to today did not occur until historical times (Irurzun et al., 2014; Laprida et al., 2014). This study constitutes the first of its type basing the reconstruction of paleoenvironmental and paleoclimatic conditions on the dynamics and functioning of the shallow lakes, studied in the context of multiproxy and multi-site analysis.
Keywords: Regimen shifts; Clear state; Turbid state; Submerged macrophytes; Climate change; Pampa plain.
INTRODUCCION
Los escenarios climáticos futuros predicen que las precipitaciones y las temperaturas extremas serán más frecuentes en muchas regiones del mundo, con profundas consecuencias para la estructura y el funcionamiento de la biosfera (Bellard et al., 2012). Un aspecto clave en este contexto es la degradación de los ambientes de agua dulce, sistemas particularmente vulnerables al cambio climático debido a la naturaleza de su aislamiento y fragmentación física dentro del paisaje, y a la presión de las actividades humanas (Woodward et al., 2010). Teniendo en cuenta la importancia de la calidad y cantidad del agua dulce y la diversidad biológica que sostienen los ecosistemas acuáticos, resulta esencial poder predecir cómo responderán estos sistemas a futuros escenarios climáticos. Para ello se requiere comprender como respondieron a diferentes condiciones ambientales del pasado.
Los estudios paleolimnológicos de alta resolución basados en múltiples indicadores biológicos (e.g., diatomeas, polen, palinomorfos no polínicos o PNPs, macrorestos vegetales, y fauna asociada de. moluscos, ostrácodos, quironómidos), geofísicos (e.g. susceptibilidad magnética), geoquímicos (e.g., TOC, TIC, potencial redox) y sedimentarios (e.g. tamaño de grano), constituyen una de las herramientas más importantes para reconstruir la evolución de los sistemas lacustres y para evaluar las respuestas de estos ecosistemas a forzantes naturales y antropogénicos. La principal ventaja que tienen estos estudios es que presentan líneas de evidencia potencialmente independientes para la reconstrucción paleoambiental. Cada indicador fósil tiene sus fortalezas y debilidades, que dependen de características tales como la presencia y abundancia, la resolución espacial y temporal, la capacidad de reacción y de recuperación y la sensibilidad a diferentes forzantes. En un enfoque basado en múltiples indicadores se busca aprovechar las fortalezas e identificar las debilidades de cada uno, con el objetivo de reconstruir condiciones paleoambientales consistentes (Lotter, 2003; Birks y Birks, 2006). Cuando las reconstrucciones paleoambientales a partir de múltiples indicadores se realizan en una red de sistemas semejantes, análisis de múltiples sitios o multi-sitios, las inferencias obtenidas se pueden extrapolar y generar reconstrucciones de carácter regional consistentes en el espacio y en el tiempo, mediante el uso de un marco cronológico común. Debido a su baja profundidad, los lagos someros son ecosistemas de agua dulce altamente vulnerables a las fluctuaciones climáticas (variaciones en las precipitaciones y temperaturas), ya que variaciones en la columna de agua favorecen o restringen la penetración lumínica, la acción del viento y la consecuente resuspensión del sedimento, lo que a su vez afecta a las comunidades biológicas que en ellos habitan (Jeppesen et al., 1998; Quirós et al., 2002; Scheffer y van Nes, 2007; Torremorell et al., 2007; Lagomarsino et al., 2011). Estos ambientes dinámicos presentan diferentes estados de equilibrio. Un estado claro, donde la penetración lumínica es alta y permite el desarrollo de macrófitas sumergidas, que dominan en el cuerpo de agua, estabilizan la columna de agua evitando la resuspensión del sedimento y disminuyen la carga de nutrientes. El otro estado alternativo, turbio, está caracterizado por abundante fitoplancton, baja penetración lumínica y alta carga de nutrientes (Scheffer y Jeppesen, 2007; Scheffer y van Ness, 2007). En la llanura Pampeana Quirós et al. (2002) describen un tercer estado con alta turbidez inorgánica y escaso desarrollo de macrófitas sumergidas y de fitoplancton. En algunos casos los estados se alternan de manera periódica, y en otros se estabilizan por largos períodos de tiempo. Desde el punto de vista limnológico, la llanura Pampeana presenta mucho interés debido a la cantidad de lagos someros que se encuentran dispersos en toda su extensión. Estos lagos someros varían tanto en el tamaño como en sus características limnológicas (e.g., Torremorell et al., 2007; Allende et al., 2009), y si bien su cantidad es difícil de estimar, según Geraldi et al. (2011) el número se eleva aproximadamente a 13.000, tomando en cuenta sólo aquellos que poseen superficies mayor a 10 ha. A pesar de este elevado número, recién en las últimas décadas se han realizado estudios que muestran el potencial que estos sistemas tienen como archivos paleoambientales (Stutz et al., 2002, 2010, 2012; Borel et al., 2003; Fontana, 2005; Laprida y Valero Garcés, 2009; Laprida et al., 2009, 2014; Piovano et al., 2009; Piovano, 2011; Córdoba, 2012; De Francesco et al., 2013; Hassan, 2013; Irurzun et al., 2014; Tonello et al., 2014).
En particular en el sudeste de la llanura Pampeana existe una gran cantidad de lagos someros con características semejantes en cuanto a origen, rasgos geomorfológicos y vegetación. Este particular aspecto permite realizar un estudio de tipo multisitios, que al combinarse con análisis de múltiples indicadores, posibilita obtener información detallada como base de las interpretaciones de las condiciones paleoambientales y paleoclimáticas a escala regional. El objetivo de este trabajo es reconstruir la historia ambiental de los lagos someros del sudeste de la llanura Pampeana desde el Holoceno medio, en el contexto de un análisis de múltiples indicadores en múltiples sitios, y realizar inferencias paleoclimáticas de carácter regional. Para lograr este objetivo se analizó el contenido polínico, de palinomorfos no polínicos y de macrorestos vegetales, así como de fauna asociada, en testigos sedimentarios obtenidos de cinco lagos someros.
ÁREA DE ESTUDIO
La llanura Pampeana está ubicada en la región central de Argentina (33°-39°S; 57°-66°O), en un área transicional entre las regiones subtropical y patagónica. Los lagos someros representan los ecosistemas acuáticos dominantes en la región y su mayor densidad se concentra en las zonas más planas, deprimidas y húmedas. Estos sistemas han sido caracterizados como lagos poco profundos, polimícticos (sin estratificación térmica permanente), con tiempos de permanencia del agua, profundidades, áreas y salinidad altamente variables, naturalmente eutróficos y hasta hipereutróficos (Quirós y Drago, 1999; Sosnovsky y Quirós, 2006). Son susceptibles de ser colonizados por macrófitas emergentes y sumergidas, lo que puede convertir al espejo de agua en una extensa zona litoral. El sustrato de estas cuencas lacustres son sedimentos del Pleistoceno tardío y del Holoceno (Piovano et al., 2009; Stutz et al., 2010, 2012, 2015). Los lagos Lonkoy (37º12'S; 57º25'O), Hinojales- San Leoncio (37º23'S; 57º23'O) (de ahora en adelante Hinojales-SL), Tobares (37°30'S; 57°28'O), Hinojales (37º34'S; 57º27'O) y Nahuel Rucá (37º37'S; 57º26'O), son lagos someros pequeños a medianos (200-300 ha de superficie, profundidad 0,8-1 m) ubicados en el sudeste de la llanura Pampeana (Fig. 1). En la llanura Pampeana el nombre de los lagos someros se repite en muchos casos (Cartas topográficas del IGM); en la región sudeste existen por los menos dos lagos llamados Hinojales por lo que a uno de ellos se le agregó el nombre de la estancia donde está ubicado (Hinojales-SL). Estos lagos, como la mayoría de los lagos del área, están desarrollados en cubetas de deflación de tamaño variable, desde decenas de metros hasta 3 km en diámetro. Su forma es relativamente redondeada en vista en planta y presentan paleodunas (lunettes) que los bordean por sus márgenes noreste-norte-sudeste, cuya altura oscila entre 1 y 15 m (indican una dirección predominante de los vientos desde el oestesudoeste). Ambas geoformas fueron probablemente formadas por la acción eólica durante el Pleistoceno tardío (Tricart, 1973; Zárate y Tripaldi, 2012); con el establecimiento de condiciones de mayor humedad en el Holoceno temprano (e.g. Zárate et al., 2000), estas áreas se convirtieron en descarga de agua subterránea y de acumulación de agua superficial, llenándose gradualmente con sedimentos. Si bien constituyen lagos permanentes, están sometidos a ciclos de sequías-inundaciones periódicos, lo que disminuye significativamente su volumen de agua, llegando algunos a desecarse totalmente.

Figura 1. Mapa de ubicación.
Figure 1. Location map.
La vegetación regional es un pastizal caracterizado por la familia Poaceae que es la más importante tanto en número de especies como abundancia, acompañada por Asteraceae, Cyperaceae, Solanaceae, Brassicaceae, Apiaceae y Chenopodiaceae (León, 1991). La vegetación fue modificada en las características florísticas y estructurales desde el arribo de los europeos en el siglo XVI. Particularmente, el sudeste tuvo un limitado desarrollo agrícola pero una importante actividad ganadera, que se intensificó durante los últimos 100 años y que continúa hasta la actualidad. Sin embargo disturbios endógenos al ecosistema, como las inundaciones, revierten los efectos del pastoreo favoreciendo el crecimiento de gramíneas perennes nativas y reduciendo la presencia de dicotiledóneas exóticas (Chaneton et al., 2002). Probablemente esta sea la causa por la cual es posible encontrar actualmente áreas de pastizales similares a los originales. Este sector de la llanura Pampeana carece de árboles nativos, excepto por la presencia de la especie azonal Celtis ehrenbergiana que crece sobre las paleodunas pleistocenas y sobre los cordones conchiles del Holoceno hasta los 38°S, aunque en los últimos años se ha extendido a lo largo de los alambrados como consecuencia de la dispersión zoófila de sus semillas. Las comunidades de macrófitas (emergentes, flotantes y sumergidas) de los lagos objeto de estudio, están caracterizadas por un anillo periférico de Schoenoplectus californicus (junco), donde crecen otras macrófitas emergentes como Zizaniopsis bonariensis, Typha latifolia, Hydrocotyle bonariensis, H. ranunculoides, Alternanthera philoxeroides, Solanum glaucophyllum, Bacopa monnieri, Polygonum punctatum, Ranunculus apiifolius, Triglochin striata y Phyla canescens. En las zonas reparadas se forma una densa carpeta de Ricciocarpus natans (hepática), Azolla filiculoides (helecho), Lemna valdiviana, Limnobium laevigatum, Wolffia brasiliensis, Wolffiella lingulata y W. oblonga. Myriophyllum elatinoides, Ceratophyllum demersum, Potamogeton pectinatus, Zannichellia palustris y algas carofitas son algunas de las macrófitas sumergidas que suelen aparecer en estos lagos, cuando la penetración de luz es suficiente para su desarrollo. El clima del área es templado con una temperatura media anual de 13,8ºC, y medias de enero de 20ºC y de julio de 7,3ºC (Estación meteorológica Mar del Plata Aero, 37º33'S, 57º21'O; Servicio Meteorológico Nacional). El valor anual de precipitación es de 950 mm. El régimen de precipitación experimenta un ciclo estacional pronunciado, con máximos de lluvia en diciembre y marzo relacionados con una intensa actividad convectiva de masas continentales y oceánicas inestables, y el desarrollo de un sistema tipo monzónico (Vera et al., 2006) que influye hasta el sur de los 35°S. Durante la estación seca (invierno) prevalece la actividad convectiva relacionada con sistemas frontales, siendo el anticiclón del Atlántico Sur la fuente principal de humedad (Labraga et al., 2002). Con frecuencia, el patrón de circulación atmosférica es modificado por incursiones de aire polar que originan precipitaciones frontales. Durante el verano, los vientos son predominantes desde el norte, noreste y este; y desde el noroeste o sudoeste en invierno.
MATERIALES Y MÉTODOS
Los registros de los lagos Hinojales, Nahuel Rucá y Lonkoy ya han sido analizados y publicados (Stutz et al., 2002, 2010, 2012) y se incluyen en este trabajo con el objetivo de reconstruir la historia ambiental en un contexto de estudio multi-sitio. Los testigos sedimentarios se obtuvieron en la parte central y más profunda de los lagos. El testigo de la laguna Hinojales (110 cm), se obtuvo con una sonda tipo Dachnovsky; los testigos de Nahuel Rucá (123 cm), Hinojales-SL (230 cm) y de Tobares (160 cm) fueron tomados con un muestreador tipo vibracorer; y el testigo Lonkoy (142 cm), con una sonda tipo Livingston. Todos los testigos fueron submuestreados en el laboratorio cada 1 cm para los análisis de polen, palinomorfos no polínicos (PNPs) y macrorestos vegetales y fauna asociada. En los testigos de Hinojales y de Tobares no se analizaron macrorestos vegetales. Se aplicaron técnicas estandarizadas para la extracción de los palinomorfos (polínicos y no polínicos): KOH, HCl, separación por densidad con líquidos pesados, ZnCl2, HF y acetólisis (Faegri e Iversen, 1992). Dos tabletas de esporas de Lycopodium clavatum se agregaron a cada muestra previamente al tratamiento físico-químico. Los recuentos se realizaron al microscopio óptico con un aumento final de 1000x. Los granos de polen y los PNPs fueron identificados, contabilizados y expresados como porcentajes de diferentes sumas totales. Los porcentajes de los tipos polínicos de la vegetación local se calcularon sobre la suma total de polen que excluye a los tipos polínicos de especies exóticas y de larga distancia y a las esporas. Los porcentajes de los tipos polínicos de especies exóticas y de larga distancia, de las esporas y de los PNPs se calcularon sobre la suma de los tipos polínicos locales más la suma de los tipos polínicos de especies exóticas y de larga distancia, de las esporas y de los PNPs, respectivamente. Los macrorestos vegetales y la fauna asociada se analizaron en muestras de 1 cm de espesor. El volumen de cada muestra se calculó por desplazamiento de agua y los valores fluctuaron entre 2 y 8 cm3. Las muestras se tamizaron con mallas de 200 y 125 µm. El contenido fue examinado con microscopio binocular con un aumento final de 50x. Los fósiles fueron aislados, identificados y contabilizados. El número de estructuras/individuos se calculó por 1 cm3 de sedimento.
Cronología
La cronología para los diferentes testigos sedimentarios se obtuvo sobre la base de fechados radiocarbónicos (Tabla 1). En cada uno de ellos se estimó la edad de las muestras analizadas a partir de la construcción de modelos edad-profundidad ajustados a los fechados radiocarbónicos obtenidos y al año de extracción del testigo como edad del tope. Se aplicó CLAM 2.2 (Blaauw, 2010), un código escrito en lenguaje R (R CoreTeam2014) que calibra las edades radiocarbónicas, en este caso con la curva de calibración del Hemisferio Sur (SHCal13, Hogg et al., 2013) y ajusta curvas con algoritmos de distinto grado de complejidad. Se aplicaron regresiones tipo "smoothing spline" para los registros de Hinojales- SL, Nahuel Ruca y Lonkoy (Fig. 2a-c), en tanto que para los registros de Hinojales y Tobares se aplicó interpolación lineal, ya que poseen pocos fechados (Fig. 2d, e). Los modelos logran distintos grados de ajuste que dependen de la distancia entre edades y de la asimetría de las distribuciones de probabilidad de las edades calibradas (Blaauw y Heegaard, 2012). Para cada testigo se calcularon 1000 curvas a partir de edades tomadas al azar de la distribución de probabilidades de cada edad calibrada (iteración tipo Monte Carlo). Luego, se determinó la curva con mayor probabilidad (mejor ajuste) y se calcularon los intervalos de confianza para las estimaciones realizadas.
Tabla 1. Dataciones radiocarbónicas (AMS) de los lagos Hinojales-SL, Lonkoy, Nahuel Rucá, Hinojales y Tobares.
Table 1. AMS radiocarbon dates from Hinojales-SL, Lonkoy, Nahuel Rucá, Hinojales and Tobares lakes.

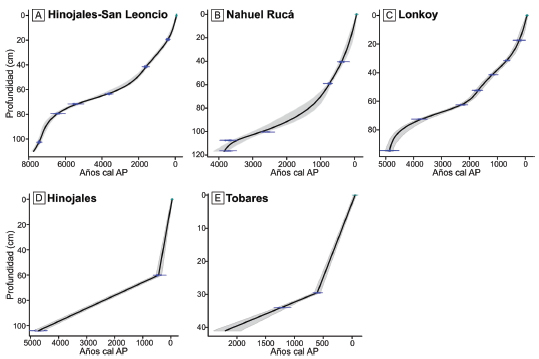
Figura 2. Modelos de edad-profundidad mediante regresiones "smoothing spline" en a) Hinojales-SL, b) Nahuel Ruca y c) Lonkoy, y mediante interpolación lineal en d) Hinojales y e) Tobares.
Figure 2. Age-Depth models applying "smoothing spline" in a) Hinojales-SL, b) Nahuel Ruca, and c) Lonkoy, and lineal interpolation in d) Hinojales and e) Tobares.
Resultados
Los resultados de los recuentos de los diferentes indicadores se presentan en diagramas porcentuales, para los palinomorfos, y de concentración para los macrorestos vegetales y fauna asociada (Figs. 3-6).

Figura 3. a) Diagrama polínico y de palinomorfos no polínicos o PNPs (porcentajes) en la laguna de Hinojales-SL. b) Diagrama de macrorrestos vegetales y fauna asociada (individuos/cm3 de sedimento) en la misma laguna.
Figure 3. a) Pollen and NPPs diagram (percentages) of lake Hinojales-SL. b) Plant macroremains and associated fauna diagram (individuals/cm3 of sediment) from the same lake.
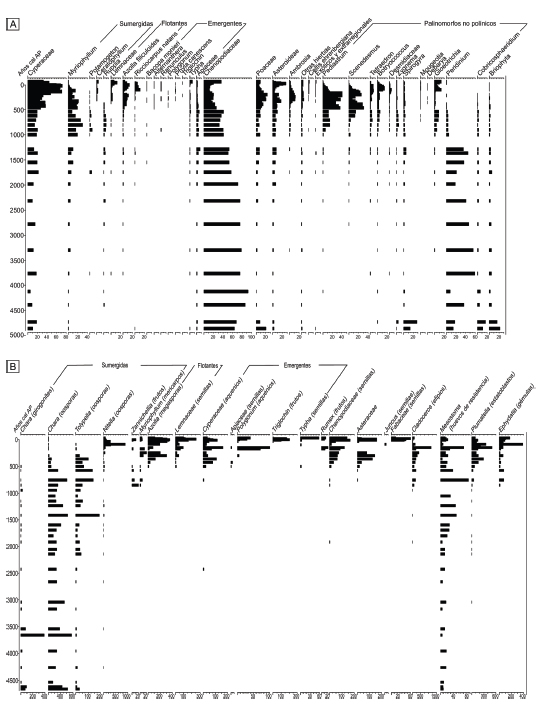
Figura 4. a) Diagrama polínico y de PNPs (porcentajes) en la laguna Lonkoy. b) Diagrama de macrorrestos vegetales y fauna asociada (individuos/cm3 de sedimento) en la misma laguna. Ambas figuras modificadas de Stutz et al. (2012).
Figure 4. a) Pollen and NPPs diagram (percentages) of lake Lonkoy. b) Plant macroremains and associated fauna diagram (individuals/cm3 of sediment) from the same lake. Both figures were modified from Stutz et al. (2012).

Figura 5. a) Diagrama polínico y de PNPs (porcentajes) de la laguna Nahuel Rucá. b) Diagrama de macrorrestos vegetales y fauna asociada (individuos/cm3 de sedimento) de la misma laguna. Ambas figuras modificadas de Stutz et al. (2010).
Figure 5. a) Pollen and NPPs diagram (percentages) of lake Nahuel Rucá. b) Plant macroremains and associated fauna diagram (individuals/cm3 of sediment) from the same lake. Both figures were modified from Stutz et al. (2010).

Figura 6. a) Diagrama polínico y de PNPs (porcentajes) de laguna Hinojales (modificado de Stutz et al. 2002). b) Diagrama polínico y de PNPs (porcentajes) de laguna Tobares.
Figure 6. a) Pollen and NPPs diagram (percentages) of lake Hinojales (modified from Stutz et al. 2002). b) Pollen and NPPs diagram (percentages) of lake Tobares.
Macrorestos vegetales y fauna asociada
En todos los lagos se observa una primera fase dominada por carofitas, principalmente del género Chara (girogonites y oósferas), que como máximo, se extiende desde ca. 7.000 hasta ca. 700-500 años. En particular desde ca. 7.000 hasta ca. 700 años cal. AP en Hinojales-SL, entre ca. 4.700 y ca. 700 años cal. AP en Lonkoy y entre ca. 3.500 y ca. 700 años cal AP en Nahuel Rucá. En Hinojales-SL y en Lonkoy las oósferas de la carofita Tolypella se incrementan con posterioridad a los ca. 2.500 años cal. AP y se mantienen similares en ambos lagos hasta ca. 500 años cal. AP (200 y 300 oósferas/cm3) y, continúan hasta ca. 300-200 años cal. AP en Lonkoy. En estos dos lagos se observa abundancia de girogonites de Chara hasta ca. 3.500 años cal AP (100 y 600 girogonites/cm3), y se evidencia un incremento de las oósferas a partir de ca. 4.500 años cal AP (1.500 y 800 oósferas/cm3). En Nahuel Rucá esta tendencia se mantiene, si bien el registro comienza a los ca. 3.500 años cal AP, con valores menores (550 oósferas/cm3 de sedimento). En los tres lagos los máximos valores de carofitas (oósferas) se registran entre ca. 2.500 y 700 años cal AP; y a partir de ca. 700-500 años cal AP se evidencia la disminución de Chara y el aumento de la diversidad en el espectro. Aparecen oósferas de la carofita Nitella, frutos de Myriophyllum y de Zannichellia palustris, semillas de Potamogeton (macrófitas sumergidas), megasporas de Azolla filiculoides y semillas de diferentes géneros de Lemnaceae (macrófitas flotantes), aquenios de Cyperaceae, semillas de Typha y de Polygonum y frutos de Triglochin (macrófitas emergentes), además de semillas de Chenopodiaceae y frutos de Asteraceae. Entre la fauna asociada, en la laguna Lonkoy entre ca. 4.700 y 200 años cal AP se registran huevos de resistencia del platelminto Mesostoma, que se incrementan a partir de ca. 2.300 años cal AP, momento en el que comienzan a registrarse en Hinojales-SL. A partir de ca. 700-500 años cal. AP, aparecen huevos de resistencia de los cladóceros Daphnia, Ceriodaphnia, Simocephalus y Bosmina (efipios); del briozoo Plumatella (estatoblastos) y del porífero Ephydatia (gémulas).
Polen y Palinomorfos no polínicos (PNPs)
En los cinco lagos se observa una primera fase dominada por Chenopodiaceae, que como máximo se extiende desde ca. 7.000 hasta ca. 700-500 años cal. AP y presenta diferentes acompañantes. En Hinojales-SL, esta fase se extiende desde ca. 7.000 hasta ca. 700 años cal AP, y acompañan a Chenopodiaceae (45-80%) Peridinium (20-80%), entre los PNPs, y Briophyta (20-60%). Cyperaceae (40%) se incrementa a los ca. 2300 años cal AP, luego disminuye y vuelve a aumentar a los ca. 1.200-1.000 años cal AP junto con Myriophyllum, Potamogeton, las flotantes Lemnaceae, Azolla filiculoides y Ricciocarpus natans y Poaceae. En el espectro de PNPs, con anterioridad a los ca. 1.300 años cal AP se registran algunas esporas de resistencia de Zygnemataceae (Zygnema, Spirogyra y Mougeotia, algas filamentosas bentónicas), y los máximos valores de Peridinium y Briophyta. Luego, comienzan a incrementarse las clorofitas del fitoplancton, Pediastrum, Scenedesmus, Tetraedron, Tetrastrum, Desmidiaceae y Botryococcus, como así también la cianobacteria Gloeotrichia. Peridinium decrece rápidamente, y más aún luego de ca. 700 años cal AP. En Lonkoy, entre ca. 5.000 y 1.300 años cal AP, domina en el espectro Chenopodiacae (60-90%), acompañada por Peridinium (20-80%). Cyperaceae comienza a incrementarse a partir de ca. 1.700 años cal AP, junto con Myriophyllum y Poaceae. Entre los PNPs, con anterioridad a los ca. 1.300 años cal AP se registran algunas esporas de resistencia de Zygnemataceae (Zygnema y Spirogyra), y los máximos valores de Peridinium y Briophyta. A los ca. 1.000 años cal AP se incrementan Pediastrum, Scenedesmus, Tetraedron, Tetrastrum, Desmidiaceae, Botryococcus y Gloeotrichia. Peridinium decrece rápidamente, y más aún luego de ca. 500 años cal AP. En Nahuel Rucá entre ca. 3.800 y 700-500 años cal AP, Chenopodiaceae presenta los máximos valores (50%) y está acompañada por Cyperaceae y Myriophyllum. Entre los PNPs se registran para este momento los mayores valores de Zygnemataceae (Zygnema y Spirogyra), Peridinium, Cobricosphaeridum y Briophyta. El espectro de PNPs refleja a partir de ca. 500 años cal AP una comunidad fitoplanctónica caracterizada por las clorofitas Pediastrum, Scenedesmus, Botryococcus y Desmidiacae. En Hinojales entre ca. 5.800 y 700-500 años cal AP el espectro polínico está caracterizado por Chenopodiaceae (70%) acompañada por Cyperaceae y Poaceae; entre los PNPs domina Cobricosphaeridium y en menores valores se presentan Peridinium, Pediastrum y esporas de resistencia de Zygnemataceae. En Tobares entre ca. 2.100 y 700 años cal AP el espectro está dominada por Chenopodiaceae (60%) acompañado por Cyperaceae. Los mayores valores de Peridinium y Briophyta se registran en la base del espectro, aunque en comparación con los valores de los otros registros, son bajos.
A partir de ca. 700-500 años cal AP en los cinco lagos comienzan a disminuir los valores de Chenopodiaceae y los espectros polínicos comienzan a reflejar la comunidad de actual de macrófitas caracterizada por las emergentes Cyperaceae, Bacopa monnieri, Alternanthera philoxeroides, Ranunculus, Polygonum, Phyla canescens, Triglochin, Apiaceae (incluye Hydrocotile bonariensis, H. ranunculoides, Eryngium) y Typha; las sumergidas Myriophyllum, Potamogeton, Ceratophyllum y Ruppia y las flotantes de la familia Lemnaceae, Azolla filiculoides y Ricciocarpus natans. Entre los PNPs se incrementan y dominan las clorofitas Pediastrum, Scenedesmus, Tetraedron, Tetrastrum, Desmidiaceae, Botryococcus, y la cianobacteria Gloeotrichia.
DISCUSIÓN
Evolución de los ecosistemas acuáticos
Todos los indicadores analizados en los cinco lagos: polen, PNPs y macrorestos vegetales y fauna asociada, son consistentes e indican un evidente cambio en estos ecosistemas a partir de ca. 700-500 años cal AP. Si bien la antigüedad de los registros es diferente, todos los lagos presentan el mismo patrón de evolución, desde una fase clara, que cambia gradualmente hacia un estado turbio general, con alternancia de fases claras (Fig. 7).

Figura 7. Modelo conceptual de la evolución de los lagos, estados tróficos y condiciones paleoclimáticas.
Figure 7. Conceptual model for lake evoution, trophic states and paleoclimatic conditions.
Fase Clara. Con anterioridad a los ca. 700-500 años cal AP, estos lagos, eran cuerpos de agua someros, de alta concentración iónica, alcalinos y con muy buena penetración lumínica. Desde el Holoceno medio y hasta este momento (ca. 700-500 años cal. AP), las comunidades de macrófitas sumergidas evolucionaron, desde comunidades dominadas por carofitas hacia comunidades con mayor diversidad, carofitas y angiospermas. La sucesión es la siguiente: la comunidad dominada por Chara, acompañada por Myriophyllum, Potamogeton y Ceratophyllum, cambia a los ca. 2.000 y hasta los ca. 500 años cal AP hacia una comunidad con mayor diversidad, caracterizada por la disminución de Chara y el aumento de las carofitas Tolypella y Nitella y de las angiospermas Myriophyllum, Zannichellia, Ruppia, Potamogeton y Ceratophyllum. Las carofitas constituyen un indicador muy sensible y una herramienta en la clasificación de los estados tróficos o ecológicos de un lago y la limitación lumínica es el principal factor determinante para su crecimiento, en particular para el género Chara (Pelatachy et al., 2014). Estas algas desaparecen con el aumento de los niveles de nutrientes (eutrofización) y con el deterioro del ambiente lumínico de la columna de agua (Blindow, 1992). Las carofitas son altamente competitivas y desplazan a otras macrófitas sumergidas del grupo de las angiospermas a bajas concentraciones de nutrientes. Cuando la situación se revierte y se eleva la carga de nutrientes en el sistema, la competencia se inclina en favor de las angiospermas, y en particular las especies del género Chara son las primeras en desaparecer (Bakker et al., 2010; Ritcher y Gross, 2013). En general las macrófitas, sean carofitas o angiospermas, son un factor clave en la estructuración de los lagos someros, ya que estabilizan la columna de agua impidiendo la remoción de sedimentos, y pueden persistir en los ambientes hasta que la carga de nutrientes alcanza niveles intermedios a altos. A medida que aumenta la turbidez se produce una pérdida en la diversidad de especies de macrófitas sumergidas, y una vez superado un nivel crítico éstas llegan a ser eliminadas de los sistemas. Entonces estos sistemas pasan a estar dominados por el fitoplancton, produciéndose un cambio de estado, hacia uno de tipo turbio. Sayer et al. (2010) y Hilt et al. (2013) indican cambios en la comunidad de macrófitas sumergidas desde una comunidad rica en especies de carofitas hacia una comunidad pobre, dominada por especies de los géneros Potamogeton y Zannichellia, en lagos someros del Hemisferio Norte. Otros indicadores de condición de aguas claras son las esporas de resistencia de las clorofitas filamentosas de la familia Zygnemataceae (Zygnema, Spirogyra, Mougeotia, Debarya), algas generalmente bentónicas y/o epífitas, que necesitan buena penetración lumínica para su desarrollo.
La presencia de quistes del dinoflagelado Peridinium y de huevos de resistencia del platelminto Mesostoma sugieren desecaciones periódicas de los cuerpos de agua, como así también la comunidad halófita desarrollada en los alrededores de estos sistemas, caracterizada por Chenopodiaceae. Asimismo Hassan (2013) señala condiciones de alta conductividad entre ca. 4.800 y 1.000 años cal AP en Lonkoy, indicadas por la abundancia de las diatomeas Cyclotella meneghiniana, Coconeis placentula, Nitzchia hustedtiana y Hyalodicus subtilis. En los lagos Hinojales-SL, Lonkoy y Nahuel Rucá, De Francesco et al. (2013) registran moluscos durante todo el lapso, con dominancia de Heleobia parchappii, acompañada por Biomphalaria peregrina y Heleobia australis, una especie estuarina y/o de marismas, que presenta en los tres lagos su máxima abundancia entre los ca. 3.700 y 2.200 años cal AP.
Fase turbio-clara. A partir de los ca. 700-500 años cal AP, los valores de ciertos componentes del fitoplancton como las clorofitas coloniales Pediastrum, Scenedesmus, Tetraedron, Desmidiaceae y la cianobacteria Gloeotrichia aumentan; en algunos lagos muy abruptamente. Esto indica un incremento en la turbidez del sistema y/o en la frecuencia de estados turbios. Estudios de la composición de diatomeas en los lagos Nahuel Rucá y Lonkoy también indican la ocurrencia de condiciones turbias para este momento. Hassan (2013), sugiere para Lonkoy entre 1.200 y 200 años cal AP condiciones eutróficas, turbidez orgánica y baja penetración lumínica, indicado por la abundancia de Aulacoseira granulata y Cyclotella meneghiniana; y a partir de los ca. 200 años cal AP, un incremento en el aporte de nutrientes y de turbidez inorgánica, indicado por la dominancia diatomeas perifíticas, por ejemplo Navicula peregrina, Coconeis placentula, Nitzchia amphibia. En Nahuel Rucá a partir de los ca. 740 años cal AP, el ensamble compuesto por Coconeis placentula, Aulacoseira granulata y Cyclotella meneghiniana también indica condiciones de turbidez (Stutz et al., 2010). Actualmente las variables ambientales que determinan la distribución del fitoplancton son el nivel de nutrientes, el contenido de fósforo total (TP) y de nitrógeno (N), el coeficiente de extinción lumínico (Kd), la concentración de carbono orgánico disuelto (DOC) y la conductividad (Izaguirre et al., 2012). Estudios anuales y estacionales de lagos someros del este y sudeste de la llanura Pampeana indican que los tipos dominantes en el fitoplancton corresponden a diatomeas pennadas y de gran porte, cianobacterias nostocales como Gloeotrichia, Anabaena, Rivularia y clorofitas coloniales como Pediastrum y Scenedesmus y las desmidiaceas Cosmarium, Staurastrum y Closterium (Allende et al., 2009; Sánchez et al., 2010; Izaguirre et al., 2012). Sin embargo, estos múltiples indicadores de turbidez coinciden con la presencia de Nitella, Myriophyllum y Ceratophyllum en la mayoría de los ambientes, quienes indican condiciones claras. En consecuencia, y en relación a la fase previamente registrada, se propone que la turbidez de los sistemas aumentó y que estos sistemas presentaron alternancias de fases turbias y claras que en el registro se refleja en la coexistencia de ambos tipos de indicadores. Por otro lado, el registro polínico y de macrorestos vegetales señala el establecimiento de la comunidad de macrófitas característica de estos lagos del sudeste de la llanura Pampeana, compuesta por las emergentes Cyperaceae, Bacopa monieri, Alternanthera philoxeroides, Ranunculus, Polygonum, Phyla canescens, Typha, Apiaceae y Buddleja; las sumergidas Myriophyllum, Zannichellia, Ruppia, Potamogeton y Ceratophyllum y la carofita Nitella; y las flotantes Ricciocarpus natans, Azolla filiculoides y Lemnaceae. En Nahuel Rucá, Tobares y Lonkoy, esto se pronuncia a los ca. 500 años cal AP.
En cuanto a los restos de fauna asociada, se incrementan los efipios de los cladóceros, en Lonkoy luego de los ca. 700, en tanto que en Nahuel Rucá y en Hinojales-SL este incremento se registra luego de los ca. 500 años cal AP. Corresponden a los géneros Daphnia, Ceriodaphnia, Simocephalus y Bosmina (González Sagrario, datos inéditos), que actualmente viven en estos ambientes y se alimentan del fitoplancton. El incremento de los huevos de resistencia de Mesostoma, en los lagos Lonkoy e Hinojales-SL se podrían atribuir al aumento de los cladóceros, que forman parte de su dieta y/o a las condiciones de alta turbidez que podrían limitar la eficiencia visual de sus predadores (Dumont et al., 2014 y bibliografía citada en ese trabajo).
Evolución paleoambiental e inferencias paleoclimáticas
Los cinco lagos estudiados respondieron sincrónicamente y mostraron el mismo patrón de evolución paleoambiental durante el Holoceno medio y tardío. El registro conjunto de vegetación halófita, comunidades de carófitas y estructuras de resistencia (Peridinium y Mesostoma) indica que los cuerpos de agua clara que se desarrollaron desde el Holoceno medio estuvieron sometidos a fluctuaciones hidrológicas. La alternancia de períodos de sequía - inundación y/o de períodos de alta evaporación habrían ocasionado: 1) la salinización de los cuerpos de agua y de los terrenos circundantes, sobre los que se desarrollaba la comunidad halófita, como así también 2) variaciones en la columna de agua, provocando la muerte de las carofitas, y la abundancia en el registro de sus estructuras de resistencia, que durante un nuevo ciclo de inundación habrían restablecido la comunidad. Este paisaje de cuerpos de agua someros, claros, salobres y bordeados de vegetación halófita caracterizó el sudeste de la llanura Pampeana por largos periodos de tiempo (en el caso de Hinojales-SL unos 6.500 años aproximadamente). Este ambiente dinámico e inestable se habría desarrollado bajo condiciones climáticas más secas que las actuales. Probablemente estas condiciones serían consecuencia del debilitamiento del sistema monzónico en verano como lo señalan los modelos de circulación atomosférica (e.g. Zhao y Harrison, 2012).
Hasta el momento, las condiciones climáticas inferidas para el Holoceno medio y tardío en la llanura Pampeana presentan discrepancias que han sido asociadas a un incremento en la regionalización del clima durante el Holoceno (Barrientos y Pérez, 2005; Mancini et al., 2005). Aunque también es probable que se deban a diferencias en la resolución espacial y temporal de los registros y/o a la diferente sensibilidad de los indicadores analizados. El escenario paleoclimático propuesto más aceptado para el sudeste y sur de la llanura Pampeana es el de condiciones de mayor aridez relativa a los 5000- 4000 años AP (e.g. Zárate et al., 1998; Zárate, 2005; Vilanova et al., 2010; Laprida et al., 2014). Los resultados obtenidos en este trabajo permiten inferir condiciones climáticas secas desde los ca. 7.000 años cal AP, que en ningún momento llegan a ser áridas. Sin embargo, se registran períodos de menor disponibilidad hídrica entre los ca. 4.000 y 2.000 - 1.500 años cal AP sugeridos por el mayor desarrollo de la comunidad halófita. A los ca. 2.000 años cal AP, el incremento en la diversidad de las macrófitas sumergidas, indica un incremento en la carga de nutrientes a la cuenca. Uno de los mecanismos posibles que explicaría este mayor aporte de nutrientes sería la alternancia entre períodos de sequía y de lluvia. Durante los períodos de sequía, se generaría mucho material orgánico en descomposición disponible en el suelo para ser transportado por el viento hacia la cuenca, o por escorrentía durante los períodos de lluvias. La materia orgánica podría provenir de la misma comunidad halófita que rodeaba a estos lagos, que comienza a disminuir. Es decir, que si bien las condiciones climáticas durante el Holoceno medio y tardío (hasta los ca. 700 años cal AP) fueron predominantemente secas, se registraban periodos de lluvias intensas. A partir de ca. 2.000 años cal AP comienza un cambio gradual hacia condiciones más húmedas que se efectiviza a los ca. 700-500 años cal AP. Coincidente con estos resultados, un estudio realizado en la laguna La Brava (37º52'S; 57º59'O) indica un aumento en el contenido de materia orgánica y en la producción primaria asociados a un aumento en el nivel de agua a los ca. 2.000 años cal AP, mientras que la mayor proporción de arenas en el registro sedimentario estaría indicando un aumento en el aporte fluvial a la laguna a los ca. 1.600 años cal AP (Laprida et al., 2014). A partir de ca. 700-500 años cal AP, se establecen condiciones de mayor turbidez que fluctúan con condiciones claras en un paisaje que presenta un mayor carácter de estabilidad. A partir de este momento se infieren condiciones climáticas húmedas similares a las actuales. El incremento en las precipitaciones habría sido un proceso gradual hasta alcanzar las condiciones climáticas actuales. En la laguna La Brava se detecta el inicio de condiciones más húmedas entre 2.000-1.500 años cal AP, las que se establecen a los ca. 300 años cal AP y continúan hasta la actualidad (Irurzun et al., 2014; Laprida et al., 2014).
Existen escasos estudios paleolimnológicos que permitan una discusión que conlleve a un consenso en la reconstrucción paleoambiental y paleoclimática a escala regional para la llanura Pampeana. Muy probablemente, existan situaciones locales sobreimpuestas a las condiciones regionales, que dificultan las interpretaciones. Sin embargo, los resultados del análisis de múltiples indicadores en cinco sitios del sudeste de la llanura Pampeana permite proponer al clima como el principal forzante de la historia de estos lagos. En este sentido, la precipitación surge como la variable plausible para explicar la dinámica de los cambios. Es evidente, que el Holoceno medio y casi todo el Holoceno tardío estuvieron caracterizados por condiciones climáticas más secas que las actuales, al menos, en el sudeste. Sin embargo, pareciera ser que el régimen de precipitación y la duración y/o intensidad de periodos húmedos/secos fue cambiando. Durante la última parte del Holoceno tardío, comienzan a establecerse condiciones húmedas, asociadas a un aumento gradual en las precipitaciones, que son similares a las actuales recién en tiempos históricos. La resolución temporal de los registros no permite realizar inferencias climáticas que puedan ser discutidas con los registros históricos y/o instrumentales de información climática, ni extender la discusión tomando en consideración al efecto antrópico como factor de cambio de estos ecosistemas.
CONCLUSIONES
Los lagos someros del sudeste de la llanura Pampeana están activos por lo menos desde el Holoceno medio, sin embargo su configuración actual (nivel trófico y comunidades biológicas) es muy reciente, ca. 700-500 años cal AP. A partir de la elucidación del patrón de funcionamiento de estos lagos se realizaron reconstrucciones paleoambientales y paleoclimáticas para el sudeste de la llanura Pampeana durante el Holoceno medio y tardío. Estos lagos evolucionaron desde sistemas claros, salobres en los que dominaban las carofitas, hacia sistemas turbios en los que alternaban fases turbias y claras, con predominio de fitoplancton y de macrófitas sumergidas, respectivamente. Durante el Holoceno medio y por largos períodos (alrededor de 6.500 años) el paisaje, conformado por lagos someros, claros, salobres y con vegetación halófita circundante, se mantuvo debido a constantes disturbios provocados por la alternancia de períodos de sequía - inundación y/o períodos de alta evaporación bajo condiciones climáticas más secas que las actuales. A partir de ca. 2.000 años cal AP se detecta un aumento en las precipitaciones, que se efectiviza a partir de los ca. 700-500 años cal AP y que alcanza gradualmente condiciones similares a las actuales en tiempos históricos.
Las poblaciones de los diferentes organismos responden de manera diferente a los cambios que se dan en la evolución a largo plazo de un ecosistema acuático, en este caso un lago somero. Estos cambios responden tanto a forzantes externos (e.g. climáticos, geomorfológicos) como a internos (e.g. carga de nutrientes, eutrofización, competencia por luz). Por esta razón tratar de responder y elucidar las causas desde un solo indicador es potencialmente engañoso cuando se utiliza el registro paleolimnológico para inferir los cambios pasados. Un enfoque más significativo es el uso de análisis de múltiples indicadores para reconstruir los cambios en la diversidad global de los hábitats y las relaciones tróficas en un lago. Aunque este tipo de análisis consumen mucho tiempo y más aún cuando le sumamos el análisis de múltiples sitios, los resultados ofrecen ideas mucho más reveladoras que las obtenidas a partir de un solo indicador estudiado en un solo sitio.
Agradecimientos
Los autores agradecen a los organismos que financiaron esta investigación (CONICET PIP 465 y UNMDP EXA 587/12) y a los dueños y encargados de las estancias que permitieron el ingreso a sus campos. En particular a P. Urrutia, J. Pueyrredón de Lanús, R. Handley, M. y R. Arbelaiz, y a los dueños de la Estancia Tarruhé. A dos evaluadores anónimos por las valiosas sugerencias aportadas a la primera versión de este manuscrito.
Referencias bibliográficas
1. Allende, L., G. Tell, H. Zagarese, A. Torremorell, G. Pérez, J. Bustingorry, R. Escaray e I. Izaguirre, 2009. Phytoplancton and primary production in clear-vegetated, inorganicturbid, and algal-turbid shallow lakes from the Pampa plain (Argentina). Hydrobiologia 624:45-60. [ Links ]
2. Bakker, E.S., E. Van Donk, S.A.J. Declerck, N.R. Helmsing, B. Hidding y B.A. Nolet, 2010. Effect of macrophyte community composition and nutrient enrichment on plant biomass and algal bloms. Basic and Applied Ecology 11:432-439. [ Links ]
3. Barrientos, G. y S.I. Perez, 2005. Was there a population replacement during the Late mid-Holocene in the southeastern Pampas of Argentina? Archaeological evidence and paleoecological basis. Quaternary International 132:95-105. [ Links ]
4. Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W. y F. Courchand, 2012. Impacts of Climate Change on the future of biodiversity. Ecological letters 15:365-377. [ Links ]
5. Birks, H.H. y H.J.B. Birks, 2006. Multi-proxy studies in palaeolimnology. Vegetation History and Archaeobotany 15:235-251. [ Links ]
6. Blaauw, M. y E. Heegaard, 2012. Estimation of age-depth relationship. En H.J.B. Birks, S. Juggins, A. Lotter y J.P. Smol (Eds.), Tracking environmental change using lake sediments. Vol. 5: Data handling and numerical techniques. Kluwer Academic Publishers, Dordrecht:379-413. [ Links ]
7. Blaauw, M., 2010. Methods and code for ‘classical' age-modelling of radiocarbon sequences. Quaternary Geochronology 5:512- 518.
8. Blindow, I., 1992. Long- and short-term dynamics of submerged macrophytes in two shallow eutrophic lakes. Freshwater Biology 28:15-27. [ Links ]
9. Borel, C.M., G.R. Guerstein y A.R. Prieto, 2003. Palinomorfos acuáticos (algas y acritarcos) del Holoceno de la laguna Hinojales (Buenos Aires, Argentina): interpretación paleoecológica. Ameghiniana 40:531-544. [ Links ]
10. Chaneton, E.J., S.B. Perelman, M. Omacini y R.J.C León, 2002. Grazing, Environmental Heterogeneity and Alien Plant Invasions in Temperate Pampa Grasslands. Biological Invasions 4:7-24. [ Links ]
11. Córdoba, F.E., 2012. El registro climático del Holoceno tardío en latitudes medias del SE de Sudamérica: Limnogeología de las Lagunas Encadenadas del Oeste, Argentina. Tesis de Doctorado en Ciencias Geológicas, Universidad de Córdoba, 285 pp. (inédito). [ Links ]
12. De Francesco, C.G., E. Tietze y P.A. Cristini, 2013. Mollusk succesions of Holocene shallow-lake deposits from the southeastern Pampa plain, Argentina. Palaios 28:851-862. [ Links ]
13. Dumont, H.J., A.C. Rietzler y B.P. Han, 2014. A review of typhloplanid flatworm ecology, with emphasis on pelagic species. Inland Waters 4:257-270. [ Links ]
14. Faegri, K. y J. Iversen, 1992. Texbook of Pollen Analysis, IV Edition. En K. Faegri, P. E. Kalana y K. Krzywinski (Eds.). John Willey and Sons, New York:328-332. [ Links ]
15. Fontana, S.L., 2005. Holocene vegetation history and palaeoenvironmental conditions of the temperate Atlantic coast of Argentina, as inferred from multiple proxy lacustrine records. Journal of Paleolimnology 34:445-469. [ Links ]
16. Fontana, S.L., 2007. Radiocarbon chronologies of Holocene lacustrine sediments from the southern coast of Buenos Aires province, Argentina. Radiocarbon 49:103-116. [ Links ]
17. Geraldi, A.M., M.C. Piccolo y G.M.E. Perillo, 2011. Lagunas bonaerenses en el paisaje pampeano. Ciencia Hoy 21:17-22. [ Links ]
18. Hassan G.S., 2013. Diatom-based reconstruction of Middle to Late Holocene paleoenvironments in Lake Lonkoy, southern Pampas, Argentina. Diatom Research 28:473-486. [ Links ]
19. Hilt, S., J. Köhler, R. Adrian, M.T. Monaghan y C.D. Sayer, 2013. Clear, crashing, turbid and back - long-term changes in macrophyte assemblages in a shallow lake. Freshwater Biology 58:2017-2036. [ Links ]
20. Hogg, A.G., Q. Hua, P.G. Blackwell, M. Niu, C.E. Buck, T.P. Guilderson, T.J. Heaton, J.G. Palmer, P.J. Reimer, R.W. Reimer, C.S.M. Turney y S.R.J. Zimmerman, 2013. SHCal13 southern hemisphere calibration, 0 - 50,000 cal BP. Radiocarbon 55:1889-1903. [ Links ]
21. Irurzún, A., C.S.G. Gogorza, A.M. Sinito, M.A.E. Chaparro, A.R. Prieto, C. Laprida, J.M. Lirio, A.M. Navas y H. Nuñez, 2014. A high resolution palaeoclimate record for the last 4800 cal. years BP on lake La Brava, SE Pampa plains, Argentina. Geofisica Internacional 53:365-383. [ Links ]
22. Izaguirre I., L. Allende, R. Escaray, J. Bustingorry, G. Pérez y G. Tell, 2012. Comparison of morpho-functional phytoplankton classifications in human-impacted shallow lakes with different stable states. Hydrobiologia 698:203-216. [ Links ]
23. Jeppesen, E, M. Sondergaard, J.P. Jensen, P. Mortensen, A.M. Hansen y T.S. Jorgensen, 1998. Cascading trophic interactions from fish to bacteria and nutrients after reduced sewage loading: A 18-year study of a shallow hypertrophic lake. Ecosystems 1:250-267. [ Links ]
24. Labraga, J.C., B. Scian y O. Frumento, 2002. Anomalies in the atmospheric circulation associated with the rainfalk excess or deficit in the Pampa Region in Argentina. Journal of Geophysical Research: Atmospheres 107:2-15. [ Links ]
25. Lagomarsino, L, G.L. Pérez, R. Escaray, J. Bustingorry y H.E. Zagarese, 2011.Weather variables as drivers of seasonal phosphorus dynamics in a shallow hypertophic lake (Laguna Chascomús, Argentina). Fundamental Applied Limnology 178:191-201. [ Links ]
26. Laprida, C. y B. Valero-Garcés, 2009. Cambios ambientales de épocas históricas en la pampa bonaerense en base a ostrácodos: historia hidrológica de la laguna Chascomús. Ameghiniana 46:95-111. [ Links ]
27. Laprida, C, M.J. Orgeira y N. García Chapori, 2009. El registro de la Pequeña Edad de Hielo en lagunas pampeanas. Revista Asociación Geológica Argentina 65:603-611. [ Links ]
28. Laprida, C., M.S. Plastani, A. Irurzún, C. Gogorza, A.M. Navas, B. Valero-Garcés y A.M. Sinito, 2014. Mid-late Holocene lake levels and trophic states of a shallow lake from the southern Pampa plain, Argentina. Journal of Limnology 73:325-339. [ Links ]
29. León, R.J.C., 1991. Setting and vegetation. En Coupland, R.T. (Ed.), Natural Grassland: Introduction and Western Hemisphere. Elsevier, Amsterdam:371-389. [ Links ]
30. Lotter, A.F., 2003. Multi-proxy climatic reconstructions. En Mackay A., R.W. Batarrbee, H.J.B. Birks y F. Olfield (Eds.), Global Change in the Holocene. Hodder Arnold, London:373-383. [ Links ]
31. Mancini, M.V., M.M. Paez, A.R. Prieto, Stutz, M.S. Tonello e I. Vilanova, 2005. Mid-Holocene climatic variability reconstruction from pollen records (32°-52°S, Argentina). Quaternary International 132:47-59. [ Links ]
32. Pelatachy, M., E. Pronin y A. Pukacz, 2014. Charophyte occurrence in Ceratophyllum demersum stands. Hydrobiologia 737:111- 120. [ Links ]
33. Piovano, E., 2011. Laguna Mar Chiquita: a unique sensor of past and present hydroclimatic variability in south-eastern South America. 11th International Conference on Salt Lake Research:23, Miramar. [ Links ]
34. Piovano, E.L., D. Ariztegui, F.E. Córdoba, M. Cioccale y F. Sylvestre, 2009. Hidrological variability in South America below the tropic of Capricornio (Pampas and Patagonia, Argentina) during the last 13.0 Ka. En F. Vimeux (Ed.), Past climate variability in South America and surrounding regions. Developments in Paleoenvironmental Research:323-351. [ Links ]
35. Quirós, R. y E. Drago, 1999. The environmental state of Argentinean lakes: An overview. Lakes and Reservories: Research and Management 4:55-64. [ Links ]
36. Quirós, R., A.M. Rennella, M.A. Boveri, J.J. Rosso y A. Sosnovsky, 2002. Factores que afectan la estructura y el funcionamiento de las lagunas pampeanas. Ecología Austral 12:175-185. [ Links ]
37. R Core Team, 2014. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/ [ Links ]
38. Ritcher, D. y E.M. Gross, 2013. Chara can outcompete Myriophyllum under low phosphorous supply. Aquatic Sciences 75:457-467. [ Links ]
39. Sánchez, M.L., H. Pizarro, G. Tell e I. Izaguirre, 2010. Relative importance of periphyton and phytoplankton in turbid and clear vegetated shallow lakes from the Pampa Plain (Argentina): a comparative experimental study. Hydrobilogia 646:271-280. [ Links ]
40. Sayer, C.D., A. Burgess, K. Kari, T.A. Davidson y N. Rose, 2010. Long-term dynamics of submerged macrophytes and algae in a small and shallow, eutrophic lake: implications for the stability of macrophyte-dominance. Freshwater Biology 55:565-583. [ Links ]
41. Scheffer, M. y E. Jeppesen, 2007. Regime shifts in shallow lakes. Ecosystems 10:1-3. [ Links ]
42. Scheffer, M. y E.H. van Nes, 2007. Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 584:455-466. [ Links ]
43. Scheffer, M., S. Carpenter, J.A. Foley, A. Folke y B. Walker, 2001. Catastrophic shifts in ecosystems. Nature 413:591-596. [ Links ]
44. Sosnovsky, A. y R. Quirós, 2006. El estado trófico de pequeñas lagunas pampeanas, su relación con la hidrología y el uso de la tierra. Ecología Austral 16:115-124. [ Links ]
45. Stutz, S., A.R. Prieto y F.I. Isla, 2002. Historia de la vegetación de la laguna Hinojales, sudeste de la provincia de Buenos Aires, Argentina. Ameghiniana 39:85-94. [ Links ]
46. Stutz, S., A.R. Prieto y F.I. Isla, 2006. Holocene evolution of the Mar Chiquita coastal lagoon area, Argentina, indicated by pollen analysis. Journal of Quaternary Science 21:17-28. [ Links ]
47. Stutz, S., C.M. Borel, S.L. Fontana, L. del Puerto, H. Inda, F. García- Rodriguez y M.S. Tonello, 2010. Late Holocene environmental evolution of Nahuel Rucá freshwater shallow lake, SE Pampa grasslands, Argentina. Journal of Paleolimnology 44:761- 775. [ Links ]
48. Stutz, S., C.M. Borel, S.L. Fontana y M.S. Tonello, 2012. Holocene evolution of three shallow lakes in the SE Pampa plain (Argentina) as evidenced by analyses of pollen, non-pollen palynomorphs and plant macrofossils.The Holocene 22:1215- 1222. [ Links ]
49. Stutz, S., M.S. Tonello, M.A. González Sagrario y D. Navarro, 2015. Evolución ambiental de dos lagos someros del SE de la llanura Pampeana durante el Holoceno. Inferencias paleoclimáticas. VI Congreso Argentino de Cuaternario y Geomorfología Libro de Resúmenes:17, Ushuaia. [ Links ]
50. Tonello M.S., S. Stutz y D. Navarro, 2014. Variabilidad climática durante los últimos 1500 años a partir del análisis de múltiples indicadores en el Sudeste de la llanura Pampeana. XIX Congreso Geológico Argentino, Actas en CD, Córdoba. [ Links ]
51. Torremorell A., J. Bustigorry, R. Escaray y H.E. Zagarese, 2007. Seasonal dynamics of a large, shallow lake, laguna Chascomús: the role of light limitation and other physical variables. Limnologica 37:100-108. [ Links ]
52. Tricart, R., 1973. Geomorfología de la Pampa Deprimida. Base para los estudios edafológicos y agronómicos. XII Colección científica. INTA. 201pp. [ Links ]
53. Vera, C., W. Higgins, J. Amador, T. Ambrizzi, R. Garreaud, D. Gochis, D. Gutzler, D. Lettenmaier, J. Marengo, C.R. Mechoso, J. Nogues-Paegle, P.L. Silva Diaz y C. Zhang, 2006. Towards a unified view of the American Monsoon System. Journal of Climate 19:4977-5000. [ Links ]
54. Vilanova, I., A.R. Prieto, S. Stutz y E.A. Bettis, 2010. Holocene vegetation changes along the southeastern coast of the Argentinean Pampa grasslands in relation to sea-level fluctuations and climatic variability: palynological analysis of alluvial sequences from Arroyo Claromecó. Paleogeography, Paleoecology, Paleoclimatology 298:210-223. [ Links ]
55. Woodward, G., D.M. Perkins y L.E. Brown, 2010. Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philosophical Transactions of the Royal Society 365:2093-2106. [ Links ]
56. Zárate, M.A., 2005. El Cenozoico tardío continental de la provincia de Buenos Aires. En R.E. de Barrio, R.O. Etcheverry, M.F. Caballé y E. Llambías (Eds.), Geología y Recursos Minerales de la provincia de Buenos Aires. 16° Congreso Geológico Argentino:139-158. [ Links ]
57. Zárate, M.A. y A. Tripaldi, 2012. The aeolian system of central Argentina. Aeolian Research 3:401-417. [ Links ]
58. Zárate, M.A., M. Espinosa y L. Ferrero, 1998. Paleoenvironmental implications of a Holocene diatomite. Quaternary of South America and Antarctic Peninsula 12:135-52. [ Links ]
59. Zárate, M.A., R.A. Kemp, M.A. Espinosa y L. Ferrero, 2000. Pedosedimentary and palaeoenvironmental significance of a Holocene alluvial sequence in the southern Pampas, Argentina. The Holocene 10:481-488. [ Links ]
60. Zhao, Y. y S. Harrison, 2012. Mid-Holocene monsoons: a multimodel analysis of the interhemispheric differences in the responses to orbital forcing and ocean feedbacks. Climate Dynamics 39:1457-1487. [ Links ]

