










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) v.73 Vicente López ene./dic. 2004
ARTÍCULOS ORIGINALES
Perennial grass species richness and diversity on Argentine rangelands recovering from disturbance (with 3 figures & 5 tables)
Busso CA 1*, HD Giorgetti2, OA Montenegro2, GD Rodríguez2
1Departamento de Agronomía and CERZOS (Consejo Nacional de Investigaciones Científicas y Técnicas: CONICET), Universidad Nacional del Sur, Altos del Palihue, Bahía Blanca (8000), Argentina;
2Chacra Experimental de Patagones, Ministerio de Asuntos Agrarios, cc 118, Carmen de Patagones (8504), Argentina;
*Corresponding author, E-mail cebusso@criba.edu.ar.
Received 21.IV.2003: accepted 25.V.2003
Abstract. The influence of types of disturbance (hereon referred to as treatments) on natural vegetation was determined on richness and diversity of perennial grasses in semiarid rangelands of Argentina (40º 39'S, 62º 54'W). Studies were performed in adjacent areas which received different treatments: (1) untreated control; (2) burning; (3) herbicide application (shrub control); (4) Old Field 1 or (5) Old Field 2 (areas previously subjected to mechanical soil disturbance for 25 years; Old Field 1 had been exposed to a more severe grazing than Old Field 2 previous to cropping); and (6) overgrazing. During 1984-1992, annual aboveground biomass was determined per perennial grass species within each treatment after clipping permanent plots (n=30). Grass species richness and diversity were higher (p<0.05) on the Control than on sites receiving a more severe or continuous grazing (Old Field 1 and Overgrazing). With a few exceptions, the Control, Shrub Control and Old Field 2 treatments had a similar (p>0.05) perennial grass species richness and diversity. Changes in grass species diversity among years within each treatment were due to changes in both richness and relative biomasses of the studied species. The declining trends in grass species richness or diversity during 1984-1992 had equal slope in all treatments, indicating that the reduction rate of these variables over time was the same independent of disturbance type. Principal Coordinate Analysis showed that the similarity of species composition was greater among years within any treatment than among treatments in any given year. However, more than 69% of all perennial grass species were desirable perennial grasses in all treatments and years.
Key words: Argentina grasslands, rangeland disturbances, perennial grasses, species richness and diversity.
Resumen. Se determinó la influencia de tipos de disturbios (desde aquí en más llamados tratamientos) de la vegetación natural sobre la riqueza y diversidad de gramíneas perennes en pastizales semiáridos de Argentina (40º 39'S, 62º 54'O). Los estudios se efectuaron sobre áreas adyacentes que recibieron diferentes tratamientos: (1) control no tratado; (2) quema; (3) Aplicación de herbicidas (control de arbustos); (4) Campo abandonado 1 o (5) Campo abandonado 2 (áreas expuestas previamente a disturbio mecánico del suelo durante 25 años); el Campo abandonado 1 había sido expuesto a un pastoreo más severo que el Campo abandonado 2 previo a su cultivo; y (6 ) Sobrepastoreo. Durante 1984-1992, se determinó la biomasa aérea anual por especie de gramínea perenne dentro de cada tratamiento después de cortar parcelas permanentes (n=30). La riqueza y diversidad de especies de gramínea fueron mayores (p<0.05) en el Control que en sitios que recibieron un pastoreo más severo o continuo (Campo abandonado 1 y Sobrepastoreo). Con unas pocas excepciones, los tratamientos Control, Control de arbustos y Campo abandonado 2 tuvieron una riqueza y diversidad de especies similar (p>0.05). Los cambios en la diversidad de especies de gramíneas entre años dentro de cada tratamiento fueron debidos a cambios en la riqueza y biomasa relativa de las especies. La tendencia decreciente tanto en la riqueza como en la diversidad de especies de gramíneas durante 1984-1992 tuvo la misma pendiente en todos los tratamientos. Esto indica que la tasa de reducción de estas variables en el tiempo fue la misma independientemente del tipo de disturbio. El análisis de coordenadas principales mostró que la similitud de composición de especies fue mayor entre años dentro de cualquier tratamiento que entre tratamientos en cualquier año dado. Sin embargo, más del 69% de todas las especies de gramíneas perennes fueron gramíneas perennes deseables en todos los años y tratamientos.
Palabras clave: pastizales de Argentina, disturbios en pastizales naturales, gramíneas perennes, riqueza y diversidad de especies.
Concern is increasing about the effects of range improvement practices on species richness and diversity (33, 44). Cooperrider (9) pointed out that range improvement projects typically emphasize increasing livestock forage while reducing plant species diversity. Other studies have reported that both selective and non-selective grazing or soil disturbance associated with grazer activity can potentially increase or decrease plant species diversity (25). Furthermore, grazers may interact with other processes operating at different spatial and temporal scales (e.g., fire, drought, species interactions) resulting in scale dependent effects (10, 22). However, range improvement practices (e.g., woody plant control, appropriate grazing management) can be designed and planned to maintain or increase plant species richness and diversity (35,17). Maintaining biodiversity, a concept which includes species richness and diversity, is a desirable goal for range managers. Primary production may be more resistant to and recover more fully from disturbances (e.g., drought) in more diverse plant communities (34, 43). Biodiversity components can also influence structural and functional attributes of grassland ecosystems (2,43), but factors affecting biodiversity components in rangelands remain poorly understood (44).
Effects of various management alternatives on perennial grass standing crop have been examined in the south of the Phytogeographical Province of the Monte (13). In this region of Argentina, however, information is lacking on grass species richness and diversity measured during several consecutive years on fields disturbed at different times in the past, or on fields all disturbed at approximately the same time. In this paper, we use the Simpson diversity index as a measure of plant species diversity. Our objective is to report on nine years of grass species richness and diversity sampling in six areas with different disturbance histories previous to the sampling period, but undisturbed during the sampling period. This information is important in answering questions on long-term dynamics of recovery of semi-arid vegetation after disturbance.
MATERIALS & METHODS
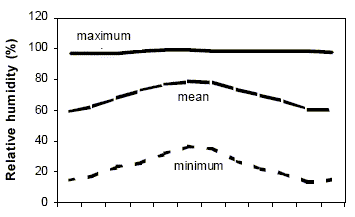
Study site. Research was conducted at the field site of the Experimental Farm of Patagones (40º 39'S, 62º 54'W; 40 m a.s.l.) which is located 22 km north of Carmen de Patagones. This site is within the Phytogeographical Province of the "Monte" (6). Long-term (1901-1950) mean annual values are 331 mm for rainfall, 14.6ºC for temperature, 60% for relative humidity and 13 km h-1 for wind speed. Absolute minimum (August) and maximum (January) temperatures are -7.6 and 43ºC, respectively. A meteorological station at the study site provided various climatic parameters during 1984-1992 as shown in Fig. 1. Regional topography is typically a plain. In ninety one percent of samples (n=44), soil texture was from sandy to sandy loam in all disturbance types (hereon referred to as treatments). Soil physical and chemical characteristics, determined following Allison et al. (1),were similar at all study sites (Table 1).





Monte vegetation is a steppe scrub dominated by microphyllous, xerophytic shrubs 1 to 3 m high (15). Perennial grasses are dominant within the herbaceous layer. In this study, perennial grasses were classified following Cano (7) according to their degree of acceptance by livestock as desirable [Bromus brevis Nees, Koeleria permollis Nees, Pappophorum subbulbosum Arech., Piptochaetium napostaense (Speg.) Hack., Poa lanuginosa Poir., P. ligularis Nees ex Steud., Sporobolus cryptandrus (Torr.) A. Gray, Stipa clarazii Ball, S. papposa Nees and S. tenuis Phil.]; intermediate (Aristidapallens Cav., A. spegazzinii Arech., A. subulata Henrard, A. trachyantha Henrard, Stipaspeciosa Trin. & Rupr., grazed when desirable species are not available); or undesirable [Sporobolus rigens (Trin.) Desv., Stipa ambigua Speg., S. brachychaeta Godron, S.trichotoma Nees, only grazed when a better forage is not available]. Except for the warm-season Sporobolus, Aristida and Papphophorum species, which show the reproductive developmental phase from mid to late summer, all other C3 perennial grasses resume growth in fall, stay vegetative during winter, and flower and set seed during spring and early summer (18).
The plant community in the study area was characterized (n=20 stands) on 1 November 1977 by using Braun Blanquet's (36), abundance-dominance/sociability index. Species with the highest index were Chuquiraga erinacea D. Don (mean=2.3), Condalia microphylla Cav. (mean=1.1), Schinus fasciculatus (Gris.) I.M. Johnston (mean=1.1), Lycium chilensis Miers ex DC. var. minutifolia (mean=1.1) and islands of Geoffroea decorticans (Gill. ex Hook. & Arn.) Burkart (mean=4.4) for the shrubby layer; Baccharis ulicina Hook. & Arn. (mean=1.2) for the forb layer, and S. tenuis (mean=4.4), S. clarazii(mean=2.3), P.ligularis(mean=1.2) and P.napostaense (mean=1.2) for the grass layer. The community was then classified as an open shrubland of Ch. erinacea and C. microphylla with islands of G. decorticans within a continuous herbaceous layer of S.tenuis (13). All study areas were similar in initial vegetation structure and botanical composition, topography and soil, supporting our assumption that areas representing different treatments were sufficiently similar in site characteristics to interpret any differences between them mainly as resulting from treatments rather than site effects.
Treatments. Several disturbances (treatments) were imposed on areas which had been exposed to continuous grazing by cattle and sheep before 1951 (Table 2). Trees and undergrowth were cleared in 1951 from two fenced areas which were at least 1 km apart. Surfaces of these areas were 4.6 (Old field 1) or 15.1 (Old field 2) hectares. Before fencing in 1951, Old field 1 was closer than Old field 2 to a watering point for cattle. Thus, since grazing effects on vegetation can be associated with distance from water (16, 45), the smaller area supported heavier grazing than the largest, more distant area before cultivation. Both of these areas were then cropped with wheat during 1951-1975; grazing was allowed on these areas once every two years (Table 2). During this period (1951-1975), an area (95 hectares) between both Old Fields was kept uncleared and grazed by cattle and sheep. Access of domestic herbivores was then excluded from both Old Fields, and the area in between, during 1975-1993 (Table 2). The 95 ha area, however, was exposed to three different disturbances between December 1977 and March 1978: one site of 34 ha remained untreated (Control); another site of 37 ha was burned (Burnt), and herbicides were applied on a 24 ha third site (Shrub control) (Table 2).

Burning was performed on 3 March 1978 when maximum and mean air temperatures were 23.5 and 14.4º C, respectively, mean relative humidity was 49%, mean wind speed was 22 km h-1, and dry weight of fine fuel load was 438±56.5 kg ha-1. At this time, mean regrowth height was 12 cm for perennial grasses; shrubs were at the maturation and dissemination stage of development. One year after burning, ≥50% of plants of G.decorticans, C.microphylla, L.chilensis, Ch.erinacea, Larrea divaricata Cav. and S.fasciculatus had produced basal regrowth.
Digiuni (13) has already reported details ofchemical shrub control in this study. Briefly, an aerial application of Tordon 213 (2 l ha-1) and 2,4,5-T (4 l ha-1) was performed on 29 December 1977. At this time, minimum, maximum and mean air temperatures were 13.6, 21.2 and 18.7º C, respectively, mean air relative humidity was 58%, mean wind speed at 4 m height was 14 km h-1, accumulated rainfall during December was 108.2 mm, and shrubby vegetation was at the reproductive developmental stage. Immediately after their application, herbicides produced death or total defoliation with no basal regrowth in C.microphylla, G.decorticans, L. chilensis and L. divaricata, and less than 50% defoliation in Ch. erinacea. Sixteen months later, regrowth had not been produced by 80-90% of C. microphylla and G. decorticans plants. The remaining plants, however, and those of L. chilensis, Ch. erinacea and L. divaricata were less than 50% defoliated. Lack of manpower prevented study ofshrub recovery afterwards.
Another adjacent site (168 ha) had been severely overgrazed until 1981 (2.5-4.5 ha/cow equivalent), and it was then excluded from domestic herbivory until 1993 (Overgrazed) (Table 2). In 1981, visible signs of severe overgrazing by sheep were noticeable in this paddock: basal area of perennial grass tussock had been grazed to the soil surface and bare soil percentage cover was large with almost no litter cover (Giorgetti, pers. comm.). None of the treatments was grazed during 1981-1992.
Sampling procedures. Unfortunately, change of research scientists at the Experimental Farm of Patagones in 1978 and economic constraints precluded data collection on all treatments during 1978-1983. During 1984-1992, maximum aboveground standing crop was obtained in all treatments by hand clipping live plus recently dead herbage by late December or early January, when major perennial grass species reached maturity. At harvesting time during this period, perennial grasses were clipped to 3-5 cm stubble height on each of 30 randomly distributed, permanent plots (0.5 ´ 0.5 m) in each treatment. Because treatments were unreplicated, the results will be treated as a case study, with statistical inferences applying only to the study area in order to avoid misinterpretation (26). Since there is only one large-scale plot for each treatment, the sampling quadrats within each plot are pseudo-replicates and not true replicates. Perennial grasses were separated by species, dried in a forced draft oven at 70º C until constant weight, and weighted.
Principal coordinate analysis was used to detect possible perennial grass species associations among years and/or treatments. With this purpose, biomasses of each of the 19 perennial grass species were averaged over the 30 quadrats, and its relative contribution to total biomass was calculated giving a proportion vector for each treatment and year. The Morisita-Horn (41) index was then calculated for any pair of proportion vectors as follows:
 (41)
(41)
where (pi1,...,pi19) and (pj1,...,pj19) corresponded to any combination of treatment and year for species (k) 1 through 19. This index takes into account species dominance: it detects greater association between vectors sharing a dominant species than between those which share species of similar importance. Vectors are represented as points within a space where distance among them represents a measure of their association; closer points then represent a greater association.
Species richness refers to the number of perennial grass species sampled within each quadrat and is expressed for each treatment and date. Species diversity, which includes both richness and evenness or equitability of species'relative biomass components, was calculated for each quadrat within each treatment and year, using the Simpson's index ![]() where p is the relative biomass of species k (41, 3).
where p is the relative biomass of species k (41, 3).
Simpson's index was used since it is implicitly considered in the algorithm of the association index of Morisita-Horn. Species present at any one treatment and year will be referred as botanical composition. Species composition will refer to relative biomasses of each species.
Statistical analysis. Statistical comparisons for grass species richness or diversity were made between the Control versus any of the other disturbance types using the non-parametric Dunnett's test (46).Contrasts were also tested between pairs or groups of treatments that had a similar disturbance history in most periods but differed in one treatment factor in one period. With this purpose, Scheffé's contrasts were applied on rank averages (46) to test the following comparisons: Old Fields versus all others, Old Field 1 versus Old Field 2, and Overgrazed versus Control+Burnt+Shrub Control. These analysis were performed using BMDP New System for Windows, Version 1.Changes in grass species richness or diversity with time (years) within each treatment were tested using linear regression analysis (37) to detect any trend over years.Regression lines were compared between treatments, and data were pooled if they were not significantly different (p>0.05 ).When regression lines were significantly different (p<0.05), slopes of these lines were tested for equality following the procedure of Neter & Wasserman (37). Briefly, a 95 percent confidence interval for the difference between any two slopes was constructed; whenever a confidence interval covered that the difference between any two slopes was equal zero, this suggested that the two slopes were the same (37).
RESULTS & DISCUSSION
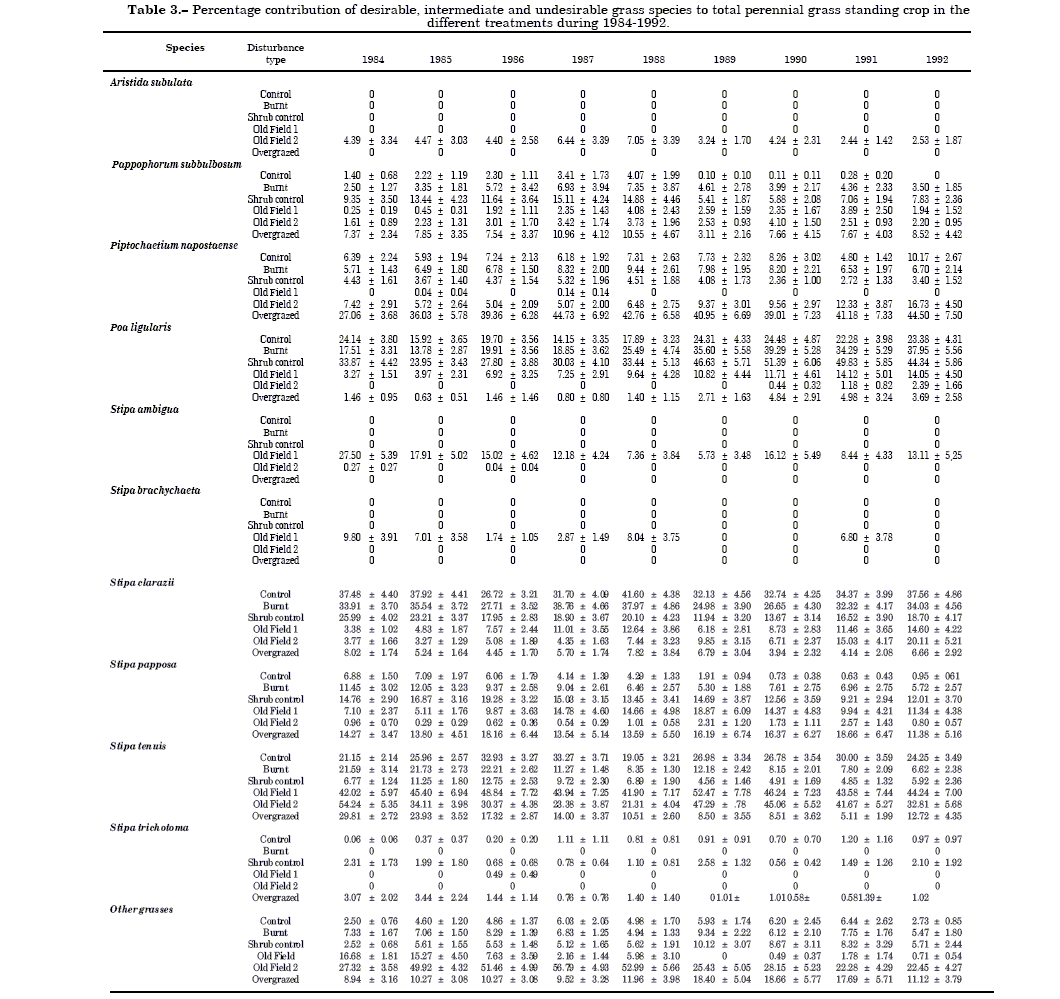
Species relative contribution to total standing crop biomass.Poa ligularis and S. clarazii contributed the least to total perennial grass standing crop in the Overgrazed treatment in our study (Table 3). Busso (5) reported that these species are the mostpreferred by cattle in semi-arid rangelands of Central Argentina, and tend to disappear from the pastures with heavy grazing. Proper grazing management aids recruitment and persistance of desired species, whereas poor management hastens their replacement by other, less desirable species (28).
Table 3. Percentage contribution of desirable, intermediate and undesirable grass species to total perennial grass standing crop in the different treatments during 1984-1992.
Undesirable perennial grasses were mostly found in the Old Field 1, which had been heavily grazed before 1951, and Overgrazed treatments (Table 3). Species of this group, such as S. ambigua, S. brachychaeta and S. trichotoma, are indicative of previous grazing mismanagement conducive to rangeland deterioration (7). Intermediate perennial grasses were mostly present in the Old Field 2 treatment (Table 3). These grasses included Aristida species (e.g., A. subulata) which may also indicate rangeland overuse (7). Another factor which may have affected the presence of species almost exclusively found in the Old Field 1 or Old Field 2 treatment is distance to seed sources. Propagules were probably destroyed by years of cultivation so that botanical composition at both Old Fields was very likely influenced by distance to sources of new propagules. The importance of distance to seed sources in revegetation of disturbances on rangeland was emphasized by Glenn & Collins (20) and Samuel and Hart (42). The abundance of seed and other propagules in the soil may be the primary control of secondary succession in grasslands (31).
Disturbance history on grass species richness and diversity. Disturbance type comparisons. The Control had a lower (p<0.05) grass species richness than the Burnt treatment in 1987, and than the Shrub Control treatment during 1985-1987 (Tables 4 & 5). Except for 1991, when grass species diversity was lower (p<0.05) in the Shrub Control than in the Control treatment, the Control, Burnt and Shrub Control treatments had a similar (p>0.05) grass species diversity during 1984-1992 (Tables 4 & 5). Also, the Old Field 2 and Control treatments had a similar (p>0.05) grass species both richness and diversity during 1985-1992, except in 1984 when grass species richness (p<0.01) and diversity (p<0.05) were lower in the Old Field 2 than in the Control treatment (Tables 4 & 5). Our results are similar to those of Nolte et al. (38) in the western Rio Grande Plain where plant species richness and diversity were similar in ephemeral drainages chained (about 40 years earlier) and then root plowed (about 30 years earlier) and in untreated rangeland. Fulbright (17) reported that plant species richness and diversity within a patch created by woody plant control may be similar, lower, or greater than plant species richness and diversity on nonmanipulated rangeland following treatment depending on various factors including the treatment method, composition of the plant community before treatment, soil series, and rainfall before and after treatment. Maintenance of biodiversity allows primary productivity to recover more fully from and be more resistant to disturbances such as drought (34, 43), a common environmental stress in our study region (5). Recent evidence has also shown that plant species diversity and the compositional and production stability of grassland plant communities are positively related (24).
Table 4. Species richness and Simpson species diversity index of perennial grasses on 0.5 x 0.5 m plots in the different treatments during 1984-1992. Each value is the mean ± 1 SE of n=30.

Grass species richness reached lower values in the Old field 1 than in the Control treatment (p<0.05) during 1986-1992, and in the Overgrazed than in the Control treatment (p<0.01) during 1988-1992 (Tables 4 & 5). Comparisons between the Old field 1 or Overgrazed treatment versus the Control for grass species diversity followed the same pattern than that reported for grass species richness, except that differences (p<0.01) between these treatments were noticed earlier (1984-1992 for Old field 1, and 1986-1992 for Overgrazed) (Tables 4 & 5). Treatments were not only individually compared with the Control. Contrasts were also made between pairs or groups of treatments that had a similar disturbance history in most period but differed in one treatment factor in one period. For example, and except in 1984 when grass species richness was greater (p<0.05) in the Old field 1 than in the Old field 2, grass species both richness and diversity reached higher (p<0.05)values in the Old field 2 than in the Old field 1 treatment during 1987-1992 for species richness and in 1985, 1986, 1989 and 1991 for species diversity (Tables 4 & 5). Also, values were higher (p<0.05) in the Control+Burnt+Shrub Control than in the Overgrazed treatment during 1986-1992 for grass species richness, and from 1985-1992 for grass species diversity (Tables 4 & 5) (32). In Missouri tallgrass prairie sites or (39) in Mediterranean grasslands plant species diversity or richness, respectively, decreased when grazing was very heavyfor many years. In the Nylsvley Nature Reserve in South Africa, low animal stocking densities over two decades have resulted in high species diversities in several plant communities (14). When grazing is moderate, however, species diversity is greater than on ungrazed sites (24), which supports specific predictions of the intermediate disturbance hypothesis and the generalized model of Milchunas et al. (35). Similarly, Noy-Meir (39) obtained a higher mean species richness and diversity at sites grazed (burnt or unburnt) mostly moderately than at those kept ungrazed on Mediterranean grasslands. In sub-humid grasslands, theoretical models also predict greater plant species diversity under moderate grazing than under ungrazed conditions (35). The increase inboth species richness and diversity under moderate grazing has been attributed to the generation of greater habitat heterogeneity. The mosaic of habitat patches generated by herbivory and non-grazing activities likely increases the diversity of colonization and establishment opportunities for several species that are otherwise excluded from the community by the strong competitive effects of the matrix dominants (24). In fact, Huntly (25) reported that both selective and non-selective grazing or soil disturbance associated with grazer activity can potentially increase or decrease plant species diversity. Furthermore, grazers may interact with other procceses operating at different spatial and temporal scales (e.g., fire, drought) resulting in scale dependent effects (32, 10, 8, 22, 39, 24).
Herbivores can mediate species diversity through differentially utilizing plants variously suceptible to defoliation; the capacity of competitive dominants to exclude other species is decreased, and gaps available for occupation by other species are created (2, 24). Initial conditions, climatic regimes, intensity and duration of grazing impact, and grazer selectivity for specific food items can also be important determinants of either increases or decreases in plant species diversity in response to grazing (23, 35). Thus, when reporting the effects of grazing on species richness and diversity we need to be aware that different herbivory types or management conditions may have different effects on these community components (e.g., see 24). Systems managed with cattle within the Phytogeographical Province of the Monte typically involve year-round grazing with minimal or no grazing management (5). In other rangelands managed for livestock production, however, grazing is often seasonal. Thus differences in the temporal and spatial patterns of grazing may differ considerably among studies and contribute to the observed differences in vegetation responses.
The Control+Burnt+Shrub Control+Overgrazed treatments had a similar (p>0.05) grass species richness during the whole study period, and a greater grass species diversity in 1984 (p<0.01), 1986 (p<0.01) and 1989 (p<0.05) than both Old Fields (Tables 4 & 5). D'Angela et al. (12) and Omacini et al. (40) reported that native rangeland species did not return to the community during early successional stages of herbaceous vegetation after interruption of agricultural activities in agroecosystems of central Argentina.This was partially attributed to unusual climatic events, such as severe inundations, in their study region (40).

Variation over time. Grass species both richness and diversity declined (p<0.05) over time in most treatments during 1984-1992, except in the Old field 2 for species diversity where this variable did not show a significant (p=0.317) trend with time (Figure 2). Noy-Meir (39) also found that species richness and diversity varied considerably between years within each category of grazing and fire treatment in Mediterranean grasslands. However, this author reported that the effect of fire on species richness and diversity in pairs of adjacent sites was much less consistent than that of grazing: in ungrazed areas, a single fire was about as likely to decrease species richness as to increase it. Noy-Meir (39) reported that in grasslands with a strong perennial component there was a significant trend of increased richness in the three years after a single fire in grazed areas. This response was attributed to stimulation of germination by fire in some Mediterranean herbaceous species (e.g., 21). Economic constraints during a few years immediately after treatment application, however, precluded us from the possibility of observing this trend. Direct negative effects of hot fires on propagule populations of some species may outweigh the gap creation and stimulation effects of fire (39). Belsky (4) also found a tendency for increased diversity in response to fire in East African grasslands, but the effect was not consistent over communities and grazing treatments.
The Control+Burnt+Old Field 2 treatments for grass species richness, and the Control+Burnt+Shrub Control treatments for grass species diversity adjusted to the same (p>0.07 in both cases) regression line when the tendency of these variables was investigated over time (Fig. 2). The Control+Burnt+Old Field 2, Old Field 1, Shrub Control and Overgrazed treatments showed different (p<0.05) regression lines in the relationship between grass species richness and time (Fig. 2). Regression lines between grass species diversity against time were also different (p<0.05) among the Control+Burnt+Shrub Control, Old field 1, Old field 2 and Overgrazed treatments (Figure 2). Despite being different, regression lines depicted in Figure 2 for grass species richness had equal slope. Similarly, all four regression lines between grass species diversity and time in Fig. 2 shared the same slope. This means that the reduction rate of grass species either richness or diversity over time was the same independently of disturbance type from 1984 to 1992. Equal slopes on different regression lines during this period also means that values for species richness or diversity in the investigated treatments were already different in 1984 (i.e., regression lines had different ordinates through the origin when observations were started in 1984). Treatment differences, as previously discussed,may then have occurred during the period from establishment of exclosures in 1975 to the time when observations began uninterruptedly in 1984. Houston (27) reported that plant species diversity generally increases during the early stages of succession and decreases during the later stages, with a maximum in mid-succession. The declining trend in species diversity over time under all disturbance types in our study suggests that the stage of mid-succession had already been reached in 1984, after nine years of vegetation exclosure. Judd & Jackson (30) and Judd (29) reported that the climax stage on abandoned farmlands in Nebraska and Montana occurred as early as 8 years but it usually required 10 to 15 years. On the shortgrass prairies of northeastern Colorado, however, Costello (11) concluded that the climax mixed prairie association may appear 20-25 years after abandonment but may require 40-50 years.
Changes in grass species diversity among years within each treatment (Table 4, Fig. 2) were due to changes in grass species both richness (Table 4, Fig. 2) and relative biomasses (Table 3) of the species. However, principal coordinate analysis showed that similarity of the composition of grass species was greater among years within any treatment than among treatments in any given year (Figure 3). Nonetheless,most (>69%) perennial grass species were desirable in all treatments and years (data not shown). Pappophorum subbulbosum, P. napostaense, P. ligularis, S. clarazii, S. papposa and S. tenuis contributed more than 83% to total perennial grass standing crop on average for all treatments and years (Table 3). Within each treatment, however, relative biomasses of each species changed considerably among years (Table 3). Rainfall appears to be the major factor in accounting for most of the variation in grass production among years in these rangelands (19).

1.Allison LE, JW Brown, HE Hayward, LA Richards, L Bernstein, M Fireman, GA Pearson, LV Wilcox, Bower CA., JT Hatcher, RC Reeve, Diagnóstico y rehabilitación de suelos salinos y sódicos. Manual de Agricultura No. 60 del Departamento de Agricultura de los Estados Unidos de América. Secretaría de Agricultura y Ganadería. Instituto Nacional de Investigaciones Agrícolas, México (1962) [ Links ]
2.Archer S, FE Smeins, In RK Heitschmidt, JW Stuth, eds, Grazing management: An Ecological Perspective, Timber Press, Portland (1991) 109 [ Links ]
3.Begon M, JL Harper, CR Townsend, Ecology. Individuals, populations, and communities. Sinauer Associates, Inc., Sunderland, Massachusetts (1986) [ Links ]
4.Belsky AJ, J Veg Sci 3 (1992) 187 [ Links ]
5.Busso CA, J Arid Environ 36 (1997) 197 [ Links ]
6.Cabrera AL, In EF Ferreira Sobral, ed, Enciclopedia Argentina de Agricultura y Jardinería. ACME, Buenos Aires (1976) 1 [ Links ]
7.Cano E, Pastizales naturales de La Pampa. Descripción de las especies más importantes (1988) Tomo 1. Convenio AACREA-Provincia de La Pampa, Buenos Aires [ Links ]
8.Collins SL, Ecology 68 (1987) 1243 [ Links ]
9.Cooperrider A, In WE Hudson, ed, Conservation of biodiversity on western rangelands. Landscape linkages and biodiversity, Island Press, Washington, D.C (1991) 40 [ Links ]
10.Coppock DL, JD Detling, J Wildlife Manage 50 (1986) 452 [ Links ]
11.Costello DF, Ecology 25 (1944) 312 [ Links ]
12.D´Angela E, RJC León, JM Facelli, Flora 178 (1986) 261 [ Links ]
13.Digiuni D, Estación Experimental IDEVI, Serie Técnica 11 (1983) 1 [ Links ]
14.Dorgeloh WG, J Range Manage 52 (1999) 519 [ Links ]
15.Fernández OA, CA Busso, In, O Arnalds, S Archer, eds, Arid and semi-arid rangelands: two thirds of Argentina.Case Studies of Rangeland Desertification. Agricultural Research Institute Report Nº 200, Reykjavik, Iceland (1999) 41 [ Links ]
16.Friedel MH, J Arid Environ 37 (1997) 145 [ Links ]
17.Fulbright TE, J Range Manage 49 (1996)554 [ Links ]
18.Giorgetti HD, Z Manuel, OA Montenegro, GD Rodríguez, CA Busso, FUTON, Int J Exp Botany 69 (2000) 91 [ Links ]
19.Giorgetti HD, OA Montenegro, GD Rodríguez, CA Busso, T Montani, MA Burgos, AC Flemmer, MB Toribio, S Horvitz, Biomasa de especies herbáceas y su correlación con la precipitación en la Provincia Fitogeográfica del Monte. XVII Reunión Argentina de Ecología, 24-28 April , Mar del Plata, Argentina (1995) 166 [ Links ]
20.Glenn SM, SL Collins, Oikos 63 (1992) 273 [ Links ]
21.González-Rabanal F, M Casal, L Trabaud, J Veg Sci 5 (1994) 289 [ Links ]
22.Grime JP, JML Mackey, SH Hiller, DJ Read, Nature 328 (1987)420 [ Links ]
23.Harper JL, The role of predation in vegetation diversity. Brookhaven Symposium in Biology 22 (1969) 48 [ Links ]
24.Hartnett DC, KR Hickman, LE Fischer Walter, J Range Manage 49 (1996) 413 [ Links ]
25.Huntly N,Ann Rev Ecol Syst 22 (1991) 477 [ Links ]
26.Hurlbert SH, Ecol Monogr 54 (1984) 187 [ Links ]
27.Huston MA, Biological diversity. The coexistence of species on changing landscapes. Cambridge University Press, Cambridge, U.K (1998) [ Links ]
28.Jones RM, JJ Mott, Trop Grasslands 14 (1980) 218 [ Links ]
29.Judd BI, J Amer Soc Agron 32 (1940) 330 [ Links ]
30.Judd BI, ML Jackson, J Amer Soc Agron 31 (1939) 541 [ Links ]
31.Kinucan RJ, FE Smeins, Am Midl Nat 128 (1992) 11 [ Links ]
32.Kucera CL, Ecology 37 (1956) 389 [ Links ]
33.Lewis CE, BF Swindel, GW Tanner, J Range Manage 41 (1988)466 [ Links ]
34.McNaughton SJ, EcolMonogr 55 (1985)259 [ Links ]
35.Milchunas DG, OE Sala, WK Lauenroth, Am Nat 132 (1988) 87 [ Links ]
36.Mueller-Dombois D,H Ellenberg, Aims and Methods of Vegetation Ecology. John Wiley and Sons, New York (1974) [ Links ]
37.Neter J, W Wasserman, Applied linear statistical models. Regression, Analysis of variance, and Experimental Designs. Richard D. Irwin, Inc., Homewood, Illinois (1974) [ Links ]
38.Nolte KR, TM Gabor, MW Hehman, MA Asleson, TE Fulbright, J Range Manage 47 (1994) 457 [ Links ]
39.Noy-Meir Y, J Veg Sci 6 (1995) 701 [ Links ]
40.Omacini M, EJ Chaneton, RJC León, WB Batista, J Veg Sci 6 (1995) 309 [ Links ]
41.Pielou E, Mathematical Ecology. John Wiley and Sons, New York (1977) [ Links ]
42.Samuel MJ, RH Hart, J Range Manage 45 (1992)444 [ Links ]
43.Tilman D,JA Downing, Nature 367 (1994) 363 [ Links ]
44.West NE, J Range Manage 46 (1993)2 [ Links ]
45.Whitford WG, DJ Rapport, AG de Soyza. J Environ Manage 57(1999) 21 [ Links ]
46.Zar JH, Biostatistical Analysis. Prentice Hall, New Jersey (1999) [ Links ]


{kind=link}
{kind=link}