










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) v.73 Vicente López ene./dic. 2004
ARTÍCULOS ORIGINALES
Axillary bud viability and dry matter production of Poa ligularis in Patagonian grasslands (with 1 table & 4 figures)
Souto CP1, GF Becker2, GL Siffredi2, CA Busso3*, M Sternberg2
1Laboratorio Ecotono, Univ. Nac. del Comahue, Centro Regional Universitario Bariloche, Quintral 1250, 8400 - Bariloche - Argentina;
2INTA EEA Bariloche, Secretaría de Agricultura, Ganadería, Pesca y Alimentación, Paraje Villa Verde s/n, 8400 - Bariloche;
3Departamento de Agronomía y CERZOS, Universidad Nacional del Sur, Altos del Palihue, 8000 - Bahía Blanca, Argentina
4Department of Plant Sciences, Faculty of Life Sciences, Tel Aviv University 69978 Tel Aviv, Israel
*To whom all correspondence should be addressed. Email: cebusso@criba.edu.ar
Received 21.IV.2003; accepted 29.V.2003
Abstract. Poa ligularis Nees is one of the dominant perennial grass species in Patagonian grasslands. This is the first field study which investigated the effects of defoliation frequency on its number of viable axillary buds, and subsequent dry matter production during one growing season. Experiments were arranged in a randomized complete block design. Treatments included one defoliation height (10 cm), from one to five times a year, and undefoliated controls. Bud respiratory activity was examined usingthe tetrazolium test and the vital stain Evan's blue.
At the end of the growing season, plants that have been defoliated more than three times showed significatively (p<0.05) lower number of respiratory active buds than unclipped controls. A high defoliation frequency increased bud death in stem bases. Both defoliated and undefoliated plants produced more (p<0.05) axillary buds at the periphery than at their center. Defoliation treatments increased shoot weight by 13.4% on average compared with undefoliated controls, but reduced in by 10% the average number of metabolically active axillary buds. This indicates that this reduction, however, was not high enough to constrain dry matter production.
It appears that Poa ligularis could tolerate moderate grazing frequency and intensity without compromising its potential regrowth capacity. Longer-term studies of the effects of defoliation on dry matter production and bud metabolic activity are needed to understand the cumulative effects of grazing of P. ligularis in natural grasslands.
Key words: defoliation, Poa ligularis, axillary buds, semiarid Argentina, dry matter production, Patagonia.
Resumen. Poa ligularis Nees es una de las especies de gramíneas perennes dominantes en los pastizales naturales de la Patagonia. Este es el primer estudio que investigó los efectos de la frequencia de defoliación sobre su número de yemas axilares viables, y subsiguiente producción de materia seca durante una estación de crecimiento. Los estudios se efectuaron utilizando un diseño de bloques completamente al azar. Los tratamientos incluyeron una altura de defoliación (10 cm), de una a cinco veces por año, además de controles no defoliados. La actividad respiratoria de las yemas fue examinada usando el método del tetrazolio y el colorante vital azul de Evans.
Hacia el final de la estación de crecimiento, las plantas que habían sido defoliadas más de tres veces mostraron un número significativamente menor (p<0.05) de yemas con actividad respiratoria que los controles no defoliados. Una alta frecuencia de defoliación incrementó la muerte de yemas en las bases de tallos. Todas las plantas- defoliadas o no, produjeron más (p<0.05) yemas axilares en su periferia que en su centro. La defoliación incrementó 13.4% el peso de los tallos en promedio, pero redujo en 10% el número promedio de yemas axilares metabólicamente activas. Esto indica que esta reducción , sin embargo, no fue suficiente como para limitar la producción de materia seca. Parece que P. ligularis toleraría una frecuencia e intensidad moderada de pastoreo sin comprometer su capacidad potencial de rebrote. Se necesitan estudios a más largo plazo de los efectos de la defoliación en la producción de materia seca y actividad metabólica de las yemas para entender los efectos acumulativos del pastoreo de Poa ligularis en los pastizales naturales.
Palabras clave: defoliación, Poa ligularis, yemas axilares, Argentina semiárida, producción de materia seca, Patagonia.
Patagonian grasslands are located within the 150 to 300 mm annual rainfall zone in Southern Argentina.This semiarid region occupies 35% of the Provinces of Río Negro (> 20 million ha) and Neuquén (> 9 million ha). Its main economic activity is animal production, mainly sheep and goats, which use natural grasslands as the major food source. Therefore, this system needs to be well understood to achieve appropriate management.
Grassland utilization essentially consists in removal of live shoots on palatable perennial grasses due to plant organ selectivity by grazers (11). Hence, to maintain shoot production under periodic defoliation is essential for both sustained grassland production and survival of grazed plants. It is then necessary to understand the fundamental processes that are directly involved in production of new shoots, and how they might be controlled by management.
Plant response to herbivory may vary between species. When plants decrease under grazing, it may be because they are either intolerant of defoliation or tolerant to this stress but highly preferred by herbivores, thus being grazed more heavily than other plants (3). On the other hand, perennial grasses which persist or increase under severe grazing have often evolved resistance mechanisms which allow them to evade herbivory (8).
Poa ligularis is a native perennial grass (34) that has become a key species for sward management in many local plant communities of Argentinean semiarid rangelands (42, 5, 43). This species is considered a good herbage during cool seasons, but it is tending to disappear under continuous grazing.Ecophysiological factors that determine its recovery ability after various defoliation frequencies have not yet been addressed.
Deciding when and to what extent plants should be defoliated are considered a critical and difficult task in range management. Understanding plant responses to defoliation allows managers to choose the proper timing for plant use and rest, which ultimately leads to a better management of range plant communities (29).
Tillering allows annual tiller replacement and thus contributes to perennation in grasses (30). Tillering and subsequent re-establishment of a photosynthetic canopy after defoliation are dependent upon the activation and subsequent growth of axillary buds if active meristematic tissues are removed by grazing (26, 48, 7, 9). Defoliation might interfere with axillary bud production, viability and/or activation, thereafter constraining the potential regrowth of a particular species (14). If frequency of this biotic stress is high enough during a growing season, it could reduce plant vigor andpersistence of desirable grass species in a community thereby leading to deterioration of rangeland vegetation (28, 52, 17). Reduction of tillering after defoliation could be due to its inhibitory effects on axillary bud activation (36, 14, 39).
In this study we tested the hypothesis that axillary bud metabolic activity and subsequent regrowth decrease as defoliation frequency increase in Poa ligularis. This is the first study which evaluates the effects of various defoliation frequencies on axillary bud viability and subsequent regrowth in perennial grasses.
MATERIALS & METHODS
Study area.During the 2001-2002 growing season, field studies were conducted at the National Institute for Agricultural Technology (INTA), Pilcaniyeu Experimental Station, located80 Km east of San Carlos de Bariloche, Patagonia, Argentina (41o8'S, 71o19'W). The study site (1 ha exclosure) is at 1000 m a.s.l., and is characterized by a flat to gently rolling topography with less than 7 % slope. It was subjected to long-term grazing until 1980, and thereafter was protected from domestic herbivory and any other agricultural use.
Climate.During 2001, hourly data on air and soil (20 cm depth) temperatures, precipitation, relative humidity and wind speed were provided by a meteorological station located at the study site (at 50 cm height). Long-term data on the same variables were provided by C. Bustos, EEAA, INTA Bariloche (personal communication). Mean air temperature was 10.7ºC, with a mean maximum in December of 16.4ºC and a mean minimum in July of -2.5ºC (Fig. 1 (a)). There were 23 freezing days in 2001, with a maximum in July of 16 days and a minimum in December of 1 day. The long-term number of freezing days for this area is 13, with a maximum in July of 18 and a minimum in December of 3. The site does not have a frost free period. Mean relative humidity is 70%, with a maximum in July of 90% and a minimum in February of 54% (Fig. 1 (b)). Winds are more intense during spring, and during 2001 had an annual mean of 7 Km h-1(Fig. 1 (b)). The long-term mean annual precipitation for the site ranges from 150 to 300 mm, and is concentrated in winter. Precipitation during 2001 was 334 mm (Fig. 1 (c)). Mean soil temperature in 2001 was 15ºC at 15 cm depth.

Soils. Soils are classified as Xerollic Haplargids, loam fine mix messic, shallow. They have the following description: A (0-5 cm) single grained sandy loam, AC (5-15 cm) dry loam sandy brown. They are structured in week subangular blocks, with abundant roots (80% of total soil volume is root biomass), and a C (15-32 cm) loam sandy clay, which is massive, with frequent pebbles, and few roots (35).
Vegetation.The area consists of a gramineous steppe of Poa ligularis-Stipa speciosa var. speciosa with isolated shrubs of Mulinum spinosum. Coverage of dominant species may vary between 40-60% (35). Poa ligularis is a native perennial, dioecious forage grass (34) highly distributed in center and southern Argentina. It is considered resistant to hervibory and drought, and forms dense bunches with cartridge-like and piercing leaves (27). It regrows in late summer-early autumn, remains vegetative during winter, elongates internodes by the end of September, flowers at the end of spring and disperses seeds in summer (6). It can be found associated with shrubs as Mulinum spinosum and Senecio spp., and to grasses of the genus Stipa. Poa ligularis prefers flat and convex topography. In natural rangelands, it is usually found severely eaten and with a low aerial coverage (= 5%). However, it may constitute up to 40% cover in areas far away from watersheds.
Experimental design and defoliation treatments. In June 2001, plants of Poa ligularis having a similar basal diameter (mean ± SD; 40.02 ± 3.00 mm; N =96) were randomly assigned to different treatments in a natural stand.Treatments included clipping to 10 cm stubble height from one (T1) to five (T5) times a year (T1: clipped on 08/01/01; T2: clipped at T1 + 10/10/01;T3: clipped at T1 and T2 + 11/10/01; T4: clipped at T1 - T3 + 12/15/01; T5: clipped at T1 - T4 + 02/15/02), in addition to an unclipped control. There were 8 replicates per treatment which were arranged in a randomized complete block design. All plants were defoliated at 5-7 cm stubble height at the end of the season (02/15/02).
Soil moisture and leaf water potential. Soil moisture was determined gravimetrically at two soil depths (0-10 and 10-20 cm) every 2 weeks during the growing season following Brown (12). At the same time, midday leaf water potentials were measured with a pressure chamber (41) using youngest fully expanded leaves. Leaf water loss was reduced by maintaining leaves in a plastic bag from excision to the end of measurement (47).
Size, position and number of metabolically active axillary buds. The effect of defoliation on the number, size, position and metabolic activity of axillary buds of Poa ligularis was determined for parent tillers (stem bases from the previous year) with green tillers, and for daughter tillers produced during the growing season. Two parent tillers with green tillers were harvested on each of 4 plants per replicate. There were 4 replicates per treatment. On each plant, one tiller was sampled from the periphery and the other one from the plant center. These harvests were made immediately before and two weeks after each defoliation treatment. After sampling, tillers were maintained at 4ºC dissected.
Axillary buds were mapped and identified on grass crowns using Mueller & Richards ´s (37) terminology. For any stem base, basal will refer to the two lowermost buds, distal to the upper bud, and intermediate to all those buds in between. Supradistal were those buds occasionally present on elongated internodes.
Observations on size and metabolic activity of axillary buds followed Busso et al. (14). Bud length was measured from its most distal portion to the point of origin on the stem using an ocular micrometer. The stem base was then cut longitudinally, leaving entire buds on each side of the cutting. Both halves were then incubated in darkness at 30ºC for 15 h in a 0.6% (w/v) solution of triphenyl tetrazolium chloride (TTC)-phosphate buffer 0.05 M plus 0.05 ml of a wetting agent (Tween 20). Staining of buds with TTC indicates enzymatic reduction of the tetrazolium salt to red, insoluble formazan. A bud was considered metabolically active when its apex stained red or pink (44). Longitudinal bud sections which did not stain with TTC and were not visibly necrotic were incubated in Evan's blue (0.25% w/w) for 30 min at room temperature (20ºC). Evan's blue is a vital stain which does not penetrate intact semipermeable membranes (24). These sections were then examined under a microscope, and buds which remained unstained or stained dark blue were considered as dormant or dead, respectively. The use of TTC and Evan's blue thus allowed the separation of axillary buds into three categories of viability: respiratorily active, dormant or dead (14).
Biomass production. Total biomass produced by P. ligularis plants (N = 72), which had been defoliated different number of times (once: T1; twice: T2; three times: T3; four times: T4; five times:T5), was determined at the time of the defoliation treatments (C, clipping). At the end of the growing cycle, the amount of dry weight produced by nondefoliated plants during the whole study period, and that accumulated by defoliated plants from the time/s of treatment to the end of the growing cycle (R, regrowth) were measured. Total dry weight produced by defoliated plants was calculated as C+R. Current-year live and recently dead materials were separated from older, previous-year grey. This separation based on leaf colour was clear and reliable and it has been reported in other studies on perennial grasses (i.e., 16). Production of dry matter per plant was measured after drying at 65ºC for 72 h. Regrowth will be used here to refer to the production of new tissue after clipping.
Statistical analysis. Information obtained from parent and daughter tillers coming either from the plant center or the periphery, or which have been collected before or after clipping, was compared using t-test for paired samples. Analysis of variance was used to compare treatments within each tiller or bud category. Comparison among treatments at each sampling date was made using nested ANOVA of four levels: five clipping treatments (main factor), four replicates per treatment, four plants per replicate and two tillers per plant. Interactions among effects were open as suggested by Willems & Raffaele (49). Dry matter production data were analyzed using one-way ANOVA. The Student-Newman-Keuls test was used to separate means when statistical differences were found at p<0.05 (46).
RESULTS & DISCUSSION
Soil humidity and leaf water potential: Soil humidity increased (p < 0.05) at greater dephts (Fig.1d).Values for this parameter were greater (p < 0.05) duringOctober-November than during December 2001 (Fig. 1d). Leaf water potentials were greater (p < 0.05) for defoliated than for undefoliated controls (Fig. 2). This result is similar to that of other authors (10, 12, 51) for perennial grasses. The positive response to herbivory has been attributed to a reduced transpiratory leaf surface area after defoliation which would conserve soil water (2, 12). Other studies, however, have reported similar leaf water potentials on defoliated and undefoliated perennial tussock grasses (14, 4, 22).

Size, position and number of metabolically active axillary buds. Before and after the defoliation treatments and for undefoliated controls, bud size was similar (p> 0.05) at the plant center (n, mean± 1 standard error: 219, 2.33±0.10 mm) than at the plant periphery (249, 2.54±0.11mm) on parent and daughter tillers. Busso et al. (14) suggested that even small bud sizes in droughted Agropyron species would not impose a limitation to regrowth following defoliation. This was because in their study even the basal buds (the smallest within any stem base) produced tillers. Our results indicate that independently of tiller location within the plant, a greater defoliation frequency will not modify the potential regrowth capacity of individual buds. In general, bud size was similar (p>0.05) for supradistal, distal, intermediate or basal buds on tillers of Poa ligularis. Overall mean bud size was 2.4 mm (N= 468, SD=1.67). This result contrasts with previous studies on perennial grasses which showed that the upper and lowermost buds were the longest and shortest, respectively in any stem base (37,14, 15, 4, 22).
Within the total number of metabolically active buds (441) studied during the whole growing season, 48 were basal, 51 intermediate, 341 distal and 1 supradistal.In our study, and those of Mueller & Richards (37), Busso et al. (14, 15), Becker et al. (4) & Flemmer et al. (22) on perennial grasses, tillering took place mostly from distal buds.
As multiple factor interactions were found in the nested ANOVA when testing the effects of several of defoliations events on the total number of metabolically active buds, we opened the interaction by treatment (49). Plants which were defoliated once during the growing season did not show significant differences (p < 0.05) in the total number of metabolically active axillary buds when compared with undefoliated controls (Fig. 3).Similar results were obtained when comparing parent versus daughter tillers, or tillers located at the plant periphery with those at the plant center (Fig. 3). Since all active meristems remained on the plants after the first defoliation, it appears that carbon resources were not limiting to maintain a metabolically active bud pool in any stem base. Busso et al. (14) indicated that previous-year stems of Agropyron species with at least one green tiller has mostly metabolically active tiller buds. Their results suggested that when one bud grew out it allowed maintenance of other buds in the same stem base. However, tillers on undefoliated controls had on average 22.4% more (p<0.05) metabolically active buds than parent and daugher tillers on plants that were clipped 4 times (Fig. 3), and 9.8% more active buds than daughter tillers on plants that were defoliated 5 times (Fig. 3). Availability of carbon resources might have been a constraint for having a greater number of metabolically active buds on these defoliated plants. Previous-year stems without green tillers have had mostly dead or dormant axillary meristems in Agropyron, Stipa and Piptochaetium species (14, 4).

Numbers of metabolically active buds were between 20 and 84.6% greater (p<0.05) on daughter than parent tillers on plants which were defoliated from 2 to 5 times (Fig. 3), and on undefoliated controls sampled at the time of the 2-, 3- or 5-time (Fig. 3) clipping treatments. Lower metabolic activity in older bud tissue, such as parent (previous-year stems) versus daughter (current-year stems) tillers, has been attributed to tissue senescence or necrosis (36, 18) and/or deterioration of vascular bundles to the buds (23, 50, 25). These factors may help to explain that the number of dead buds per stem base increased with the advancement of the growing season (Table 1). The number of dead buds per stem base, however, doubled after plants were defoliated four times in comparison to undefoliated controls (Table 1). This is the first study showing that a high defoliation frequency can increase bud death in grass stem bases.

Tillers at the plant periphery had 33% more (p<0.05) metabolically active axillary buds than those at the plant center on plants that were defoliated 4 times (Fig. 3). This result, which agrees with that of Olson & Richards (38), could be associated with differences in light quality and quantity reaching the stem base at these two locations in the plant (20, 21, 45, 19, 18).
During 2002, the total number of buds on stem bases of P. ligularis (Table 1) was higher than that reported by Becker et al. (4) for S. tenuis (2.1 ± 0.1, N = 546)and Piptochaetium napostaense (1.6 ± 0.1, N = 545). This number is similar to that observed on Agropyron desertorum and A. spicatum which can have from three to nine axillary buds per stembase (37).
Biomass production. Defoliated and undefoliated plants of P. ligularis showed a similar (p>0.05) dry matter production when yields were compared at any given time (Fig. 4); thus showing its defoliation tolerance. A rapid photosynthetic surface area reestablishment after grazing is characteristic of perennial grasses tolerant to this biotic stress (11). It appears that selective herbivory could be an important mechanism leading to local disappearance of P. ligularis from plant communities under continuous, long-term grazing. Saint Pierre (40) reported that selective herbivory of the highly competitive and grazing resistant Stipa clarazii would be the major factor driving to its replacement by less palatable perennial grasses in rangelands of Central Argentina. Anderson & Briske (2) arrived at a similar conclusion for Schizachyrium scoparium. Plants defoliated twice had a lower (p<0.05) dry matter production than those defoliated four or five times or which remained undefoliated at the time of the third or fourth defoliation (Fig. 4). These results contrast with those obtained by Klich et al. (32) in this species under greenhouse conditions, where light to severe defoliation reduced dry matter production by 30 and 44 %, respectively compared with undefoliated controls. Adjei et al. (1) also showed that dry matter production increased linearly with less frequent defoliation in several perennial grasses.
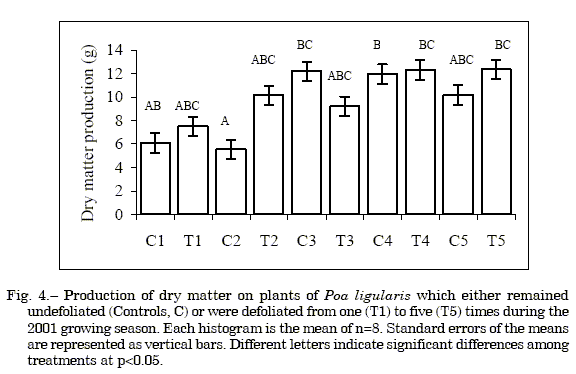
Although defoliation increased average dry matter production by 13.4% compared with undefoliated controls (Fig. 4), it reduced in 10% the average number of metabolically active axillary buds. A positive correlation in tiller populations, an expression of bud outgrowth, has been found with an increase in frequency of defoliation in perennial rangeland grasses (31, 33). Our results contrast with the overall and widely supported concept that frequent defoliations are detrimental to plant vigor (28, 52, 17). Therefore, it appears that Poa ligularis could tolerate moderate grazing frequency of moderate grazing intensity (10 cm stubble) without compromising its potential regrowth capacity. Longer-term studies of the effects of defoliations on biomass production and bud metabolic activity are needed to better understand the effects of grazing in natural grasslands.
1.Adjei MB, P Mislevy, RS Kalmbacher, P Busey, Soil Crop Sci Soc Fl Proc 48 (1989) 1 [ Links ]
2.Anderson VJ, DD Briske, Ecol Appl 5 (1995) 1014 [ Links ]
3.Archer SR, LL Tieszen, In G Olafur, ed, Plant response to defoliation: hierarchical considerations. Grazing Research at Northern Latitudes, Plenum Publishing Corp (1986) 45 [ Links ]
4.Becker GF, CA Busso, T Montani, J Arid Environ 35 (1997)233 [ Links ]
5.Bonvissuto GL, R Somlo, Guías de condición para los campos naturales de Precordillera y Sierras y Mesetas de Patagonia. Argentina: INTA-GTZ (1998) [ Links ]
6.Bran D, D Tangir, Especies silvestres patagónicas. El Coirón Poa. Argentina: INTA EEA Bariloche (1985) [ Links ]
7.Briske DD, In PJ Joss, PW Lynch, OB Williams, eds, Proc Symp Rangelands: A resource under siege. Cambridge University Press (1986) 425 [ Links ]
8.Briske DD, In RK Schmidt, JW Stuth, eds , Grazing Management: An ecological perspective. Timber Press, Portland, Oregon (1991) 85 [ Links ]
9.Briske DD, In J Hodgson,AW Illius, eds, Strategies of plant survival in grazed systems: A functional interpretation.The ecology and management of grazing systems CAB International (1996) 37 [ Links ]
10.Briske DD, VJ Anderson, Vegetatio 103 (1990)41 [ Links ]
11.Briske DD, JH Richards, In DJ Bedunah, RE Sosebee, eds, Wildland Plants: Physiological Ecology and Developmental Morphology. Soc Range Manage, Denver, Colo (1995) 635 [ Links ]
12.Brown RW, In DJ Bedunah,RE Sosebee, eds, Wildland plants: Physiological Ecology and Developmental Morphology. Soc Range Manage, Denver, Colo (1995) 291 [ Links ]
13.Busso CA, JH Richards, Acta Oecol 14 (1993) 3 [ Links ]
14.Busso CA, RJ Mueller, JH Richards, Ann Bot 63 (1989)477 [ Links ]
15.Busso CA, RM Bóo, DV Peláez, Ann Bot 71 (1993)377 [ Links ]
16.Busso C A, DD Briske, V Olalde-Portugal, Oikos 93 (2001) 32 [ Links ]
17.Cuomo GJ, BE Anderson, LJ Young, J Range Manage 51 (1998) 32 [ Links ]
18.Davies A, E Evans, Ann Bot 66 (1990) 349 [ Links ]
19.Deregibus VA,MJ Trlica, Acta Ecol 11 (1990)693 [ Links ]
20.Deregibus VA, RA Sánchez, JJ Casal, Plant Physiol 72 (1983) 900 [ Links ]
21.Deregibus VA, RA Sánchez, JJ Casal, MJ Trlica, J Appl Ecol 22 (1985) 199 [ Links ]
22.Flemmer AC, CA Busso, OA Fernández, J Range Manage 55 (2002) 150 [ Links ]
23.Fletcher GM, JE Dale, Ann Bot 38 (1974) 63 [ Links ]
24.Gaff DF, Okong´O-Ogola, J Exp Bot 22 (1971) 756 [ Links ]
25.Gifford RM, LT Evans, Annu Rev Plant Physiol 32 (1981) 485 [ Links ]
26.Hyder DN, In VB Younger, CM Mckell, eds. The Biology and Utilization of Grasses. Academic Press, New York, New York (1972) 304 [ Links ]
27.INTA, Flora patagónica. Parte III. Gramineae. Argentina: Instituto Nacional de Tecnología Agropecuaria (1978) [ Links ]
28.Jameson DA, Bot Rev 29 (1963) 532 [ Links ]
29.Jameson DA, DL Huss, J Range Manage 12 (1959) 122 [ Links ]
30.Jewiss OR, J Brit Grass Soc 27 (1972) 65 [ Links ]
31.King JW, WIC Lamb, MT McGregor, Grass Forage Sci 34 (1979)107 [ Links ]
32.Klich MG, AD Perez Mauri, RE Brevedan, In NE West, ed, Rangelands in a Sustainable Biosphere, Soc Range Manage, Salt Lake City, Utah (1995) 294 [ Links ]
33.Korte CJ, New Zeal J Agr Res 29 (1986)629 [ Links ]
34.Latour MC, Identificación de las principales gramíneas forrajeras de Chubut, Santa Cruz y Tierra del Fuego por sus caracteres vegetativos. Argentina,INTA (1978/79) [ Links ]
35.Lores RD, CA Ferreira, J De Anchorena, V Lipinski, AA Marcolín, Gaceta Agronómica 16 (1983) 660 [ Links ]
36.Mitchell KJ, Physiol Plantarum 6 (1953) 425 [ Links ]
37.Mueller RJ, JH Richards, Ann Bot 58 (1986) 911 [ Links ]
38.Olson BE, JH Richards, Oikos 76 (1988) 7 [ Links ]
39.Painter EL, JK Detling, DA Steingreber, Vegetatio 106 (1993) 37 [ Links ]
40.Saint Pierre C, Capacidad competitiva y tolerancia a la defoliación en Stipa clarazii, Stipa tenuis y Stipa ambigua. M.Sc. Thesis, Bahía Blanca, Argentina. (2002) 81 [ Links ]
41.Scholander PF, HT Hammel, ED Bradstreet, EA Hemmingsen, Science 148 (1965) 339 [ Links ]
42.Siffredi G, J Ayesa, GF Becker, J Mueller, GL Bonvissuto, Efecto de la carga animal sobre la vegetación y la producción ovina en Río Mayo Patagonia. X Reunión Nacional CAPERAS. Universidad Nacional del Sur. Bahía Blanca. (1991)183 [ Links ]
43.Siffredi GL, GF Becker, Guías de evaluación forrajera de campos para la determinación de la carga animal. Argentina: INTA-GTZ. (1998) [ Links ]
44.Smith FE, Science 113 (1951) 751 [ Links ]
45.Solangarchichi S, JL Harper, Oecologia 72 (1987) 372 [ Links ]
46.Steel RGD, JH Torrie, Bio estadística: Principios y procedimientos. Bogotá: McGraw hill (1985) [ Links ]
47.Turner NC, Proceedings International Conference of Measurements of Soil and Plant Water Status. Logan. Utah(1987) 13 [ Links ]
48.Van Loo EN, Ann Bot 70 (1992) 511 [ Links ]
49.Willems P, E Raffaele, Ecología Austral 11 (2001) 131 [ Links ]
50.Williams RF, BC Sharman, RHM, Aust J Bot 23 (1975)715 [ Links ]
51.Williams KJ, BJ Wilsey, SJ McNaughton, FF Banyikwa, Oikos 81 (1998) 463 [ Links ]
52.Youngner VB, In VB Youngner, CM McKell, eds, The Biology and Utilization of Grasses. Academic Press, New York (1972) 292 [ Links ]

