



![Similitude Pattern and Botanical Origin of the Chilean Propolis: [with 2 figures & 4 tables]](/img/es/prev.gif)






Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) v.73 Vicente López ene./dic. 2004
ARTÍCULOS ORIGINALES
Chemical growth regulators on postharvest sprout development of Sechium edule Swartz
Aung Louis H1, 2*, C Max Harris2*, Joel F Jenner1, 2*
* USDA, ARS, Postharvest Quality and Genetics Research Unit, 9611 S. Riverbend Avenue, Parlier, CA 93648, U.S.A.; We thank Judy Overstreet for typing the manuscript and Dennis Margosan for preparing the photograph.
1Author to whom correspondence should be addressed; Fax (559) 596-2791; E-mail joverstreet@fresno.ars.usda.gov
2Retired
Received 11.II.2004: accepted 23.III.2004
Abstract. Chayote is viviparous and has a tendency to sprout under postharvest storage conditions. The precocious sprout development shortens storage life, lessens fruit quality (texture) and diminishes its value for sale and consumption.Based on the hypothesis that gibberellins (GAs) are essential in initiating chayote sprout development, GAs and GA-antagonists on postharvest sprout development were determined by injection of the chemical regulators via a punched cavity in the fruit adjacent to the embryo.Exogenous application of gibberellin A1 and A3 of 1 mM promoted, while 1 mM tetcyclacis (a GA inhibitor) inhibited sprout development.Prohexadione of 1 mM also inhibited sprout emergence in semi-mature fruits and retarded the development of emergent sprout of mature fruits.The promotive action of gibberellin A1 and A3, and the inhibitory action of GA-antagonists support the view that GAs play an essential role in chayote sprout development.
Key words: Chayote, gibberellins, GA-antagonists, sprouting
The chayote, Sechium edule Sw. is a perennial valued for its immature fruits, tender sprouts, and subterranean tubers.It is unique among the cultivated cucurbits by bearing single-seeded fruit and exhibiting vivipary (5).In contrast to other multi-seeded cucurbitaceous fruit: watermelon, pumpkin, cucumber, and melon which are propagated by seeds, chayote is propagated by planting the fruit. The seed enclosed within the fleshy fruit has no dormant period, undergoes no dehydration, and sprouts readily at 20º-35 ºC temperatures (Plate 1A, B, & C).The readiness of chayote fruit to sprout under postharvest storage conditions poses a major impediment to its long-term preservation, marketing, and utilization.

Ogawa (17) demonstrated the presence of gibberellin-like substances in chayote seed extracts and found a high level of growth-promoting activity from developing seed about 5-weeks from anthesis. The peak of GA-like substances preceded the growth of the fruit and seed which suggested that these substances are involved in the development of the organs.Later, Aung and Flick (3) showed exogenous application of gibberellin A3 and A4/7 in lanolin on the stigmatic surfaces of chayote flowers stimulated the growth of the ovarian tissues, but inhibited embryo/seed development which resulted in parthenocarpic fruit. Using gas chromatography and mass spectrometry, Lorenzi and Ceccarelli (14) found gibberellin A4 and A7 were major growth substances in the seed and flesh of chayote fruit during different stages of development. Additionally, a cell-free preparation of chayote endosperm and cotyledonary tissues possessed the necessary biochemical factors to convert 14C-mevalonic acid to entkaurene and gibberellins (9).Subsequently, Albone et al. (1) identified 20 gibberellins in chayote embryo, endosperm and seedcoat, and showed gibberellin A1, A3,A4 , and A7 to be major gibberellins in chayote fruit.The observations support the view that gibberellins are important in the development of chayote seed and fruit.Therefore, it was of interest to determine the effect of gibberellins and gibberellin-antagonists on chayote sprout development under postharvest conditions.
MATERIALS & METHODS
Commercial chayote fruits of Costa Rican and Mexican origin, and locally-grown (Fresno county, California) fruits were used for investigations.The fruits of different maturity as indexed by size ranged from 250 g (less mature) to over 400 g (more mature). The Costa Rican fruits were light-green in color and broadly obovoid shape (Plate 1D). The local fruits were light-green to creamy color and thickly obovoid (Plate 1E) and the Mexican fruits were slightly darker-green on elongated obovoid shape (Plate 1F).
Chemical regulators sources and preparation. Gibberellin A3 (Sigma Chemical Company, St. Louis, Missouri, USA), gibberellin A4/7(Merck, Rahway, New Jersey, USA), gibberellin A1(Kyowa Hakko Kogyo Company, Ltd, Japan; sample courtesy of Dr. Y. Sumiki), abscisic acid (Sigma Chemical Company, Missouri, USA), Tetcyclacis (BASF, 96%, Aktiengesellschaft, Ludwigshafen, Germany) and prohexadione (BASF, 10% powder, Research Triangle Park, North Carolina, USA; sample courtesy of Dr. Reid Evans) were used. The chemical regulants at the specified concentrations were initially dissolved in 50 µl of alcohol and diluted to the desired volume with distilled water. Tween-80 (polyoxyethylenesor-bitan monoleate) was added as a surfactant. The final reagent solution contained 1% alcohol (v/v) and 0.05% Tween-80 (v/v).
Method of chemical regulators application. Gibberellins (GAs, 1mM), abscisic acid (AbA; 50 µM), tetcyclacis (TC; 1mM), and prohexadione (1mM) were injected with a Hamilton syringe via a punched fruit cavity of 5 mm diameter and 25 mm depth adjacent to the embryonic shoot of the seed.Solution volume of 400 µl of each regulant was injected per fruit cavity which was than covered with a piece of adhesive tape.Each treatment consisted of 4 replicates with 3 fruits per replicate.The treated fruits were maintained in environmental rooms at 25 ºC conducive to sprout development or at 15 ºC which retards sprouting.
Data analysis. Sprout length and emergence were used to gauge development in response to the treatments. The data obtained were analyzed using the analysis of variance procedure (18) and the least significant difference at probability level of 5% or 1% was used for comparing treatment means.Percentage data were transformed using arc sine √% before analysis of variance computation. Linear regression of sprout length and development time was calculated to depict prohexadione sprout retarding effect.
RESULTS
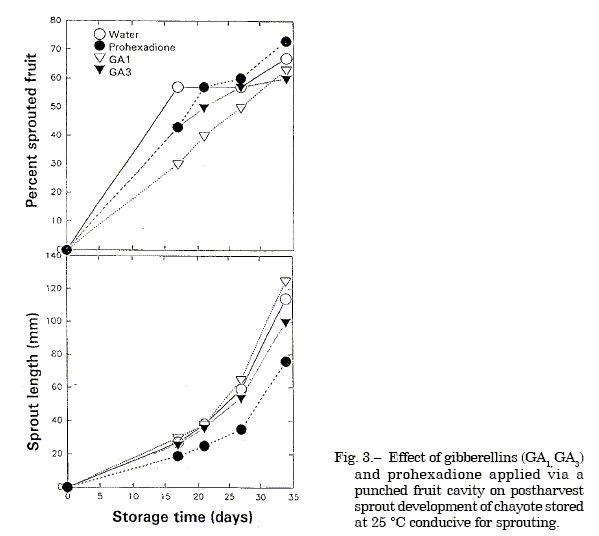
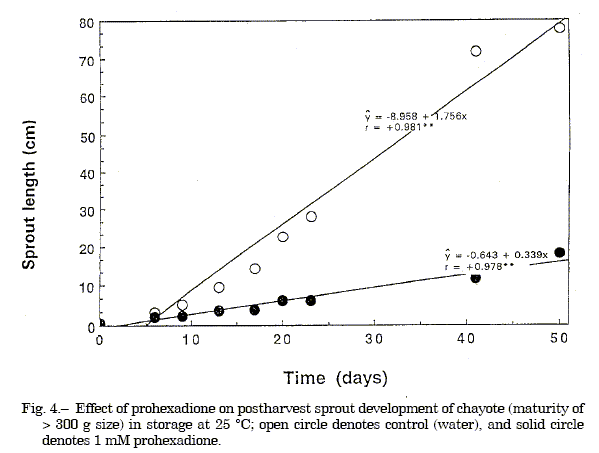
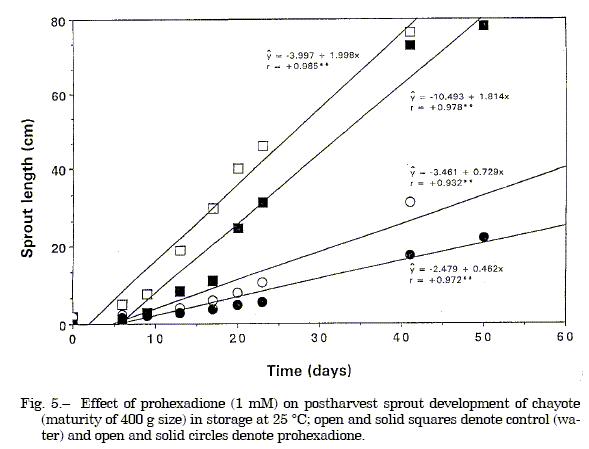
The gibberellin antagonist tetcyclacis at 1.0 mM (equivalent to 105.6 µg/fruit) injected via a punched cavity adjacent to the embryo retarded sprout development at 25 ºC compared to the untreated reagent control, but caused some tissue injury.Abscisic acid at 50µM (equivalent to 5.3 µg/fruit) injected into the fruit via a punched cavity had no inhibitory or promotive effect on sprout development at 25 ºC (Figure 1 and 2). Injection of gibberellin A3 at 1.0 mM (equivalent to 138 µg/fruit) stimulated sprout development at 15 ºC compared to the untreated reagent control which remained unsprouted (Figure 2). Shoot length was also slightly increased by GA1 application, but not by GA3 compared to control which sprouted at 25 ºC (Figure 3). Application of prohexadione at 1.0 mM concentration through a punched cavity in fruits of different maturity based on size showed that prohexadione inhibited sprout emergence in semi-mature fruits and retarded the development of emergent sprouts of mature fruits. Sprout development was reduced 50% 1-week after treatment and 78% after 3-weeks; thereafter, the inhibitory effect diminished (Figure 4 and 5). Also, the prohexadione treated fruits had compacted sprouts, leaves with enlarged laminae, short stout internodes, and reduced tendrils.


Fig. 3. Effect of gibberellins (GA1, GA3) and prohexadione applied via a punched fruit cavity on postharvest sprout development of chayote stored at 25 ºC conducive for sprouting.
Fig. 4. Effect of prohexadione on postharvest sprout development of chayote (maturity of> 300 g size) in storage at 25 ºC; open circle denotes control (water), and solid circle denotes 1 mM prohexadione.
Fig. 5. Effect of prohexadione (1 mM) on postharvest sprout development of chayote (maturity of 400 g size) in storage at 25 ºC; open and solid squares denote control (water) and open and solid circles denote prohexadione.
DISCUSSION
The phenomenon of vivipary in chayote, a herbaceous perennial plant, finds few equals in the plant kingdom. The growth habit of chayote can be contrasted with other seed-bearing crop plants where seed germination occurs only after undergoing a requisite period of dehydration (13), whereas the enclosed seed of chayote within the fleshy pericarp maintains a high water content of 80-90% on a fresh wt. basis (7) continues to grow and mature throughout ontogeny. Even in plants that do not undergo a period of dehydration for germination, their seeds are normally detached from the fruit for planting. In contrast, chayote seed remains intact within the fleshy fruit and the entire structure is used for propagation.In commercial practice, chayote fruit is harvested immature for sale and consumption.Thus, the endogenous seed continues to grow under postharvest conditions utilizing the stored water and nutrients existing at the time of severance from the mother plant (Plate 2). We have observed that a sprouted chayote fruit grew a 1-2 m long shoot during a 6-8 week period using primarily its stored water and nutrients, and with only its embryonic roots immersed in distilled water at 25 ºC under dim light. Under these conditions, the developing shoot completely resorbed and utilized the entire fruit content leaving only the epicarpic shell.Chayote seed grows continuously without dormancy or dehydration within the fleshy fruit, and it appears that exogenous postharvest application of hormonal compounds must in effect influence or modulate on-going metabolic/cellular processes rather than on the processes initiating sprouting.Furthermore, the exogenous substances could interact with endogenous hormonal substances which have a primary/direct role(s) in the initiation of sprout growth and its development.

The biological activity of endogenous GA-like substances in chayote fruit tissues was shown by Ogawa (17).Subsequently, the chemical identity of the GAs in the seed and fruit tissues of chayote was demonstrated unequivocally by GC/MS (14, 15). However, the specific function(s) of GAs in chayote growth and development is still lacking. Ceccarelli and Lorenzi (10) showed GA conjugates and 16, 17-dihydro-16-hydroxy-GA15 alcohol glucoside of cotyledonary tissues promoted shoot growth of plantlets grown under short photoperiod of 8-h in a greenhouse at 15º-25ºC. Also, a foliar spray application of GA4/7 restored shoot growth of the decotylized plantlets.From these observations, they inferred that GA-conjugates or the products of hydrolysis of the cotyledons were involved in shoot growth.Given the fact that both cytokinins (16) and indole-3-acetic acid (11) are also present in chayote seed tissues, it is not unreasonable to suggest that GAs acting synergistically or interactively with these compounds probably are involved in shoot growth (3).On the other hand, application of exogenous GA4/7 or GA3 in lanolin on the stigma of chayote flowers stimulated the development of parthenocarpic fruit with the abolishment of the seed tissues (i.e. the cotyledons and embryo).Thus, depending of the developmental stage of the tissues, the presence of the various kinds and concentrations of the bioactive substances, different growth responses may result.
Abscisic acid (AbA) generally inhibits many developmental processes and acts as an antagonist of GAs. However, the application of AbA at 50 µg concentration had no effect on postharvest shoot development of chayote fruit stored at 25EC (Figure 1). Thus, further work involving a range of AbA concentrations need to be determined. The lack of chayote response to AbA may be due to low tissue AbA sensitivity of viviparous plants (8).In chayote, the occurrence of AbA has not been shown, and thus it would be intriguing to determine if AbA occurs in viviparous chayote fruit/seed tissues, and whether the AbA content increases with dehydration as in non-viviparous plant tissues (12).
Postharvest storage temperature conditions significantly affect sprouting of chayote as shown by quicker fruit sprouting at 25 ºC than at 15 ºC (6). If a GA-conjugation mechanism which is temperature-sensitive (2), Figure 3) exists in chayote fruit, it may provide an explanation concerning its differential sprouting behavior to temperature and exogenous GAs. Thus, if the hypothesis is correct, more GA-conjugates can be expected from chayote fruit stored at 15 ºC than at 25 ºC. This needs to be determined.
Antagonists of GA, tetcyclacis, and prohexadione were effective in retarding shoot development of chayote (Figure 1 and 3). Since these compounds interfere with gibberellin biosynthesis, it was inferred that chayote shoot development which required a supply of gibberellins has been interrupted. Furthermore, if in fact a supply of GAs needed for shoot development is derived from a storage pool in the cotyledons, it would be of interest to inquire in the light of the antagonists inhibitory action whether the hydrolytic, catabolic GA process(es) or both was affected. Clarification of these aspects would provide insight regarding source(s) of GA (i.e. release from stored conjugates, transformation from precursors or from de novo synthesis in embryonic tissues) for chayote shoot development.
1.Albone KS, P Gaskin, J MacMillan, VM Spourel, Planta 162 (1984) 560 [ Links ]
2.Aung LH, AA DeHertog, GL Staby, Plant Physiology 44 (1969) 403 [ Links ]
3.Aung LH, GL Flick, HortScience 11(1976) 460 [ Links ]
4.Aung, LH, JM Byrne, J Am Soc Hort Sci 101 (1976) 189 [ Links ]
5.Aung LH, A Ball, M Kushad, Economic Botany 44 (1990) 157 [ Links ]
6.Aung LH, RE Rij, DC Fouse, JE Lindegren, FUTON 53 (1992) 125 [ Links ]
7.Aung, LH, CM Harris, RE Rij, JW Brown, J Hort Sci 71 (1996) 297 [ Links ]
8.Black M, In: WJ Davies, HG Jones, eds, Abscisic Acid Physiology and Biochemistry. Bios Scientific Publishers, Oxford, UK (1991) p 99 [ Links ]
9.Ceccarelli N, R Lorenzi, Phytochemistry 22 (1983) 2203 [ Links ]
10.Ceccarelli H, R Lorenzi, J Plant Physiology 149 (1992) 190 [ Links ]
11.Gregorio S. P Passerini, P Picciarelli, N Ceccarelli, J Plant Physiology 145 (1995) 736 [ Links ]
12.Hartung W, WJ Davies, In: WJ Davies, HG Jones, eds, Abscisic Acid Physiology and Biochemistry.Bios Scientific Publishers, Oxford, UK (1991) p 63 [ Links ]
13.Kermode AR, MY Oishi, JD Bewley, In: PC Stanwood, MB McDonald, eds, Seed Moisture.Crop Sci Soc Am Special Publ. No. 14 (1989) p 23 [ Links ]
14.Lorenzi R, N Ceccarelli, Phytochemistry 22 (1983) 2189 [ Links ]
15.Lorenzi R, N Ceccarelli, Phytochemistry 25 (1986) 817 [ Links ]
16.Lorenzi R, P Vernieri, N Ceccarelli, J Plant Physiology 133 (1988) 310 [ Links ]
17.Ogawa Y, Botanical Magazine (Tokyo) 79 (1966) 1 [ Links ]
18.Steel RGD, JH Torrie, Principles and Procedures of Statistics.McGraw-Hill, New York (1960) [ Links ]


{kind=link}
{kind=link}
{kind=link}