










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) v.75 Vicente López ene./dic. 2006
ARTÍCULOS ORIGINALES
Efecto de la densidad y estructura del dosel de maíz en la penetración de la radiación solar
(Con 3 Tablas)
Density and structure of corn population on solar radiation penetration
(With 3 Tables)
Montemayor Trejo1* José Alfredo, Alejandro Zermeño González 2, Jesús Olague Ramirez1, Rogelio Aldaco Nuncio1, Manuel Fortis Hernández1, Enrique Salazar Sosa1,3, José Cruz Rodríguez Rios1, Cirilo Vázquez-Vázquez3
1 Subdirección de Investigación y Graduados Agropecuarios, Instituto Tecnológico Agropecuario N° 10. Carretera Torreón - San Pedro km. 7.5 C.P 27170 Torreón, Coah. México. Tel. 01(871) 7 50 71 98 y 99. montemayorja@hotmail.com.
2 Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo Coahuila México. Tel (844) 4 11 03 53 y 54.
3 Facultad de agricultura y Zootecnia de la Universidad Juárez del Estado de Durango (FAZ-UJED). ENMAGEEL1@YAHOO.ES
* Investigador responsable.
**Se agradece el apoyo del personal de campo, administrativo y mayordomo del Rancho "Cerro del Agua".
Recibido 16.06.2004: aceptado 21.12.2006
Resumen. El objetivo de esta investigación fue analizar el efecto de la densidad de población y la estructura del follaje del maíz en la penetración de la radiación solar fotosintéticamente activa y su relación con algunas características del rendimiento. El trabajo se desarrolló durante el ciclo Primavera-Verano del 2000 en el Rancho "Cerro del Agua" ubicado en el kilómetro 154 de la carretera Saltillo - Matehuala. Se estableció el híbrido de maíz Prime Time en un arreglo factorial de tres por dos, bajo un diseño de bloques completos al azar con arreglo en franjas y tres repeticiones. Como factor "a" se usaron tres densidades de población (5.5, 7.4 y 11.1 plantas m-2) y como factor "b" se utilizaron diferentes inclinaciones del follaje mediante la colocación de una malla de polietileno a ambos lados de los surcos. La penetración de la radiación fotosintética activa (RFA) se midió utilizando una línea con 10 sensores cuánticos, de 70 cm de longitud. Las mediciones se realizaron sobre el dosel y en cuatro posiciones verticales dentro del follaje, realizando una lectura por segundo y promedios de un minuto en cada posición. Para evaluar la penetración de la RFA, se obtuvieron los coeficientes de extinción (K) en los diferentes tratamientos utilizando la ecuación de Lambert Beer. Los coeficientes de extinción obtenidos fueron -0.9, -1.9 y -2.44 sin malla y -0.63, -0.95 y -1.74 con malla en las densidades de 5.5, 7.4 y 11.1 plantas m-2 respectivamente. Estos resultados indican que a mayor densidad de plantas hay menor penetración de la RFA. Sin embargo, la malla indujo un menor valor de K en las densidades de población que representó una mayor penetración de la RFA. Esta mayor penetración por efecto de la malla tuvo un efecto estadístico significativo en la altura de planta, diámetro y peso del elote.
Palabras clave: Zea mayz L.; Dosel; Estructura; Coeficiente de extinción
Abstract. The objective of this study was to analyze the effect of different population densities and corn foliage structure in the penetration of photosynthetically active radiation, and its relationship to some yield parameters. The work was developed in the spring-summer cycle 2000 at the Ranch Hill Water, located in the kilometer 154 in the highway Saltillo - Matehuala. The Prime Time maize hybrid was established in a 3x2 factorial arrengement under a completely randomized block design and three replications. Two factors were studied: as factor a, three population densities (5.5, 7.4 and 11.1 plants m-2), and as factor b, different foliage inclinations to light interception. This was achieved by placement of agricultural meshes at both sides of the furrows. Penetration of the photosynthetically active radiation (PAR), was measured using a quantum sensor line, with 10 sensors instaled in a bar of 70 cm length. Measurements were taken at a second scale, and averaged every minute. Extinction coefficients in each treatments were calculated using the Lambert Beer equation. The extinction coefficients were -0.9, -1.9 and -2.44 without mesh and -0.63, -0.95 and -1.74 with mesh in the densities of 5.5, 7.4 and 11.1 plants m-2, respectively. The higher penetration of solar radiation in the lowest crop stratas in the treatments with mesh produced taller plants, of bigger diameter and weight ear.
Key words: Zea mayz L.; Canopy; Structure; Extinction coefficient
INTRODUCCIÓN
El dosel de un cultivo es una estructura compleja de un determinado arreglo espacial de las hojas que forman el cultivo. Éste actúa como un captador de radiación y como un intercambiador de vapor de agua y CO2 (Daughtry et al., 1983). La estructura de un dosel, ejerce influencia en la temperatura, concentración de vapor y régimen de radiación en el medio ambiente de la planta; afecta los procesos de fotosíntesis, transpiración, alargamiento de células, crecimiento y competencia entre especies (Russell et al., 1989). La inclinación, distribución y orientación de la hoja pueden ser importantes para determinar la intercepción de radiación (Roderick, 1981, Girardin y Tollenaar, 1994). Por ejemplo, un trébol con hojas planofilas, necesita menos área foliar para interceptar la mayoría de la radiación incidente comparado con un pasto de dosel erecto (Loomiss y Williams, 1969). En teoría, un índice de área foliar óptimo, es definido como aquel que soporta el incremento máximo de producción de materia seca. Éste es alcanzado cuando las últimas capas inferiores de hojas son capaces, en promedio, de mantener un balance positivo de carbono; es decir, cuando el cultivo virtualmente intercepta toda la radiación fotosintética activa incidente (Roderick, 1981). Se ha observado que los doseles de maíz con hojas con tendencia vertical arriba de la mazorca, pueden ser más productivos comparados con aquellos que sus hojas tienen una posición horizontal (Pendleton y Hammond, 1969). El patrón de hojas verticales en la parte superior y más horizontales en la parte inferior permite que las hojas superiores intercepten menos radiación y dejen pasar más luz hacia las hojas inferiores, logrando que la radiación sea distribuida más equitativamente sobre el total del área foliar (Loomis y Williams, 1969). La predominancia de hojas erectas en maíz justo antes de la inflorescencia origina una mayor penetración de luz, dando las tasas de producción más altas (Williams et al., 1968). Las hojas verticales fueron más eficientes en producción por unidad de índice de área foliar. Por lo tanto, el ángulo de la hoja podria ser una de las muchas características consideradas en un programa de mejoramiento (Whigham y Woolley, 1974). En líneas altas de trigo, se demostró que éstas tuvieron un 20 por ciento más de intercepción de luz y fotosíntesis durante el desarrollo vegetativo; las hojas erectas tuvieron una mayor capacidad de fotosíntesis comparadas con las caídas (Martín y Gent, 1995). Estos antecedentes sugieren que la producción del cultivo de maíz puede mejorarse incrementando la densidad de población y la inclinación de las hojas. El objetivo de esta investigación fue evaluar la penetración de la radiación solar fotosintéticamente activ en tres densidades de población, modificando la orientación normal de las hojas hacia una posición más vertical, y medir su efecto en algunas características de rendimiento.
MATERIALES Y MÉTODOS
El trabajo fue realizado en el ciclo Primavera - Verano del 2000, en el rancho denominado "Cerro del agua", ubicado en el kilómetro 154 de la carretera Saltillo - Matehuala. El clima de la región según el sistema de clasificación de Koppen, modificado por García (1981) es de tipo Bo(h´) W(e´). Este pertenece al grupo de los climas secos con una temperatura media anual de 22.8°C, con meses extremos en el invierno. El híbrido establecido fue "Prime Time", cuya característica principal es que se destina para consumo fresco por ser clasificado dentro de los maíces dulces. Tiene una altura promedio a la hoja bandera de 0.95 m. El diseño experimental fue un bloques al azar con arreglo en franjas y tres repeticiones; el arreglo de los tratamientos fue un factorial de tres por dos. En el factor "a" estuvieron tres densidades de población (5.5, 7.4 y 11.1 plantas m-2 ), el factor "b" consistió en permitir un desarrollo normal de las hojas e inducir un desarrollo vertical forzado del follaje. Para ésto se colocó una malla agrícola de un metro de ancho a ambos lados del surco, para favorecer la penetración de la radiación a los estratos inferiores y mantener una posición más vertical de las hojas.
Penetración de la radiación fotosintéticamente activa en el dosel del cultivo.
La penetración vertical de la radiación fotosintéticamente activa a través del follaje del cultivo en cada tratamiento se midió utilizando una línea de sensores cuánticos de 70 cm de longitud, Modelo Apogee Instruments, con 10 sensores. Ésta fue colocada y nivelada manualmente en forma perpendicular entre dos surcos vecinos de la unidad experimental metodología propuesta por Gallo y Daughtry (1986). Las profundidades en las que fue colocada la barra con respecto a la altura de la planta fueron: en la parte superior del dosel del cultivo, y a las profundidades relativas de 0.15, 0.33, 0.66 y 0.96, iniciando en la parte superior del dosel hacia la superficie del suelo. En la primera posición, se tomaron dos lecturas. La primera fue colocando la barra orientada hacia el zenit para medir la RFA incidente, y la segunda orientada hacia la superficie para medir la radiación reflejada por el cultivo. Las lecturas fueron tomadas y almacenadas cada segundo y promediadas por minuto, en días claros, sin nubes y a diferentes horas del día usando un Data Loger 23 X de Campbell Scientific, Inc.
Coeficientes de extinción por el cultivo. Con los valores obtenidos de la radiación fotosintéticamente activa incidente sobre el dosel del cultivo (RFAo) y la radiación medida en cada uno de los estratos o profundidades relativas en el perfil del cultivo (RFAz), donde (z) es la profundidad relativa de 0.15, 0.33, 0.66 y 0.96 de cada estrato, se obtuvieron los coeficientes de extinción del cultivo para cada tratamiento a partir de la ecuación de Lamber-Beer.
RFAz = RFAo e-KZ (1)
Donde (K) es el coeficiente de extinción de la radiación solar por el perfil del cultivo y e es la base de logaritmo natural. Rescribiendo la ecuación uno y aplicando logaritmos obtenemos:
Ln [RFAz/RFAo] = -KZ (2)
De la ecuación dos podemos observar que la pendiente de la relación de los valores de Ln[RFAz/RFAo) versus Z corresponde al valor de K. De esta forma se obtuvieron los valores de K en los diferentes tratamientos, al igual que los coeficiente de determinación (r2) de las ecuaciones. La cosecha de elote, se realizó a los 117 días después de la siembra, en forma manual, eligiendo diez elotes al azar por cada unidad experimental. Posteriormente se midió el peso, diámetro, longitud del elote y peso de la planta. El índice de área foliar (IAF) expresado como metros cuadrados de follaje por unidad de superficie de suelo, se evaluó a los 105 días después de la siembra.
RESULTADOS Y DISCUSIÓN
Coeficientes de extinción de radiación solar por el cultivo. La Tabla 1 muestra los coeficientes de extinción a la radiación solar por el dosel del cultivo a los 96, 98, 103 y 106 días después de la siembra en diferentes horas del día, y los coeficientes de determinación obtenidos en cada tratamiento. El coeficiente de extinción representa la eficiencia con que la cubierta vegetal absorbe la radiación en los diferentes estratos; depende de la estructura del dosel y de la elevación solar (Russell et al., 1989; Pauline y Seppo, 1982). Los coeficientes obtenidos fueron mayores a menor distancia entre plantas, y disminuyen conforme la separación aumenta. Este comportamiento es similar para los tratamientos con malla y sin malla. Sin embargo, en los tratamientos con malla se reducen los coeficientes de extinción, lo cual explica una mayor penetración de la radiación solar a los estratos inferiores del cultivo, hojas verticales en la parte superior y más horizontales en la parte inferior permite una más efectiva penetración y distribución de luz (Russell et al., 1969). Este incremento de la radiación en los estratos inferiores, puede resultar en un incremento en el intercambio de CO2 a nivel de planta e incrementar la producción de biomasa por unidad de superficie (Pauline y Seppo, 1982; Farquhar, 1991). La tendencia de estos resultados coinciden con los reportados por (Flenet et al., 1996) donde los coeficientes de extinción mostraron una disminución lineal cuando el espaciamiento entre hileras se incrementó. Madonna y Otegui (1996) encontraron diferentes coeficientes de extinción en diferentes híbridos y concluyeron que éstos fueron relacionados con el ángulo y la densidad de las hojas.

Efecto de la densidad de población en el índice de área foliar y altura de plantas. Los resultados del índice de área foliar, altura de la planta, diámetro, peso, longitud de elote y peso de la planta sin elote con respecto a la densidad de plantas se presentan en la Tabla 2. El mayor índice área foliar de 1.83 correspondió a la densidad de 11.1 plantas m-2 y no existieron diferencias estadísticas entre las densidades de 7.4 y 5.5 plantas m-2 (Tabla 2). Esta característica tiene una influencia directa con la radiación solar interceptada y es un componente importante en el proceso de fotosíntesis que influye en el crecimiento y producción del cultivo (Daughtry et al., 1983). Mayor IAF asociado a la densidad más alta, provoca una competencia entre plantas que favorece el alargamiento de células, la producción y afecta indirectamente los procesos de fotosíntesis y transpiración (Campbell y Norman, 1989). Con respecto a la altura de la planta, se observó una diferencia estadística al incrementar la densidad de las mismas, lo que muestra que el crecimiento del híbrido es susceptible a los cambios en densidad de población. Resultados similares fueron reportados por (Westgate et al., 1997) donde al probar altas densidades de población, observó que las altas densidades foliares inducen una alta competencia por luz, que estimulan el desarrollo vertical de las plantas.

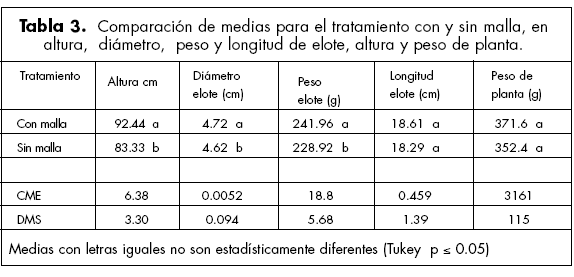
Efecto de la malla en la altura, diámetro y peso del elote. La malla mejoró la penetración y distribución de la radiación hacia los estratos inferiores del follaje, que se manifestó en una mayor altura de la planta, diámetro y peso de elote (Tabla 3). La mayor penetración y distribución de la RFA contribuye a una mayor asimilación de la radiación, que mejora la fotosíntesis del follaje del cultivo (Mock y Pearce, 1975). Boote y Loomis (1991) indicaron que la asimilación de CO2 depende del ángulo de la hoja y dimensiones del dosel. Sin embargo, el uso de la malla no tuvo efecto en la longitud de elote y el peso de la planta (Tabla 3).
CONCLUSIONES
La penetración de la radiación solar es afectada por la densidad de población y la estructura de las hojas, siendo los coeficientes de extinción mayores en la más alta densidad y sin el uso de la malla. La mayor penetración de la radiación fotosintéticamente activa hacia los estratos inferiores de las hojas del cultivo se manifestó en una mayor altura de planta, diámetro y peso de elote en los tratamientos con malla.
REFERENCIAS
1. Boote KJ, RS Loomis, Modeling canopy photosynthesis from the biochemistry of C3 chloroplast.American Society of Agronomy Journal and Crop Science Society of America, 677 S. Segoe Rd., Madison, WI 53711, USA. Modeling Crop Photosynthesis - from Biochemistry to Canopy. CSSA Special Publication, 19 (1991). [ Links ]
2. JM Norman, The description and measurement of plant canopy structure. En: Plant Canopies: Their growth, form and function. Cambridge University Press. Cambridge, England. Russell G, B Marshal, PG Jarvis (eds), (1989). [ Links ]
3. Daughtry CST, KP Gallo, ME Bauer, Spectral estimates of solar radiatión intercepted by corn canopies. Agronomy Journal 72 (1983) 527. [ Links ]
4. Evans RJ, GD Farquhar, Modeling canopy photosynthesis from the biochemistry of C3 chloroplast. American Society of Agronomy Journal and Crop Science Society of America, 677 S. Segoe Rd., Madison, WI, USA. Modeling Crop Photosynthesis - from Biochemistry to Canopy. CSSA Special Publication 19 (1991). [ Links ]
5. Flenet F, JR Kiniry, JE Board, ME Westgate, DC Reicosky, Row spacing effects on light extinction coefficients of corn, sorghum, soybean and sunflower. American Society of Agronomy 88 (1996) 185. [ Links ]
6. Gallo KP, CST Daughtry, Techniques for measuring intercepted and absorbed photosynthetically active radiation in corn canopies. Agronomy Journal 78 (1986) 752. [ Links ]
7. García E, Modificaciones al sistema de clasificación climática de Koppen. Cuarta edición. U.N.A.M., México, DF, (1981). [ Links ]
8. Girardin P, M Tollenaar, Effects of interference on maize leaf azimuth. Crop Science Society of America 34 (1994) 151. [ Links ]
9. Loomis RS, WA Williams, Productivity and the morphology of crop stands: patterns and leaves. En Eastin JD, Haskins FA, Sullivan CY, van Bavel CHM (eds), pp 28-45 (1969). Physiological aspects of crop yield. CSSA and ASA. Madison, WI. [ Links ]
10. Maddonni GA, ME Otegui, Leaf area, light interception, and crop development in maize. Field Crops Research 48 (1996) 81. [ Links ]
11. Martin PN, Gent. Canopy light interception, gas exchange, and biomass in reduced height isolines of winter wheat. Crop Science 35 (1995) 1636. [ Links ]
12. Mock JJ, RB Pearce, An ideotype of maize. Euphytica 24 (1975) 613. [ Links ]
13. Pauline OB, K Seppo, Effect of angular distribution of foliage on light absorption and photosynthesis in plant canopy: Theoretical computations. Agricultural Meteorology 26 (1982) 105. [ Links ]
14. Pendleton JW, JJ Hammond, Relative photosyntetic potential for grain yield of various leaf canopy levels of corn. Agronomy Journal 61 (1969) 911. [ Links ]
15. Roderick H, Plant growth analysis. The Institute of Biology´s. Studies in Biology, 96 (1981), University of Sheffield. [ Links ]
16. Russell G, PG Jarvis, JL Monteith, Absorption of radiation by canopies and stand growth. En: Russell G, Marshal B, Jarvis PG, (eds). Plant Canopies: Their growth, form and function. Cambridge University Press. Cambridge, England (1989). [ Links ]
17. Westgate ME, F Forcella, DC Reicosky, J Somsen, Rapid canopy closure for maize production in the northern US corn belt: Radiation-use efficiency and grain yield. Field Crops Research 49 (1997) 249. [ Links ]
18. Whigham DK, DG Woolley, Effect of leaf orientation, leaf area, and plant densities on corn production. Agronomy Journal 66 (1974) 482. [ Links ]
19. Williams WA, RS Loomis, WG Duncan, A Dovrat, F Nuñez, Canopy architecture at various population densities and growth and grain yield of corn. Crop Science 8 (1968) 303. [ Links ]

