










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) v.76 Vicente López ene./dic. 2007
ARTÍCULOS ORIGINALES
Germinación como parámetro de selección en Karwinskia por su potencial de productividad en peroxisomicina A1 (Con 3 Figuras y 3 Tablas)
Germination as a selection parameter in Karwinskia for its productivity potential in peroxisomicine A1 (With 3 Figures & 3 Tables)
Luján-Rangel1 R, E Olivares-Sáenz2, RE Vázquez-Alvarado2, L Garza-Ocañas3, O Torres-Alanís3
1 División de Estudios de Posgrado, Facultad de Agronomía, U.A.N.L., Carr. Zuazua-Marín, Km. 17.5, Marín, N. L. C.P. 66700. Address Correspondence to: M.C. Rubén Luján Rangel. Departamento de Farmacología y Toxicología de la Facultad de Medicina de la U. A. N. L., Av. Gonzalitos 235 Norte, C. P. 64460, Monterrey, N. L., México. Tel. +52 (81) 8329-4201, Ext. 2734; Fax: (81) 8348-7763. e-mail: rlujanran@yahoo.com; rlujan@fm.uanl.mx
2 División de Estudios de Posgrado, Facultad de Agronomía UANL, Carr. Zuazua-Marín, Km. 17.5, Marín, N. L. C.P. 66700.
3 Departamento de Farmacología y Toxicología de la Facultad de Medicina de la U. A. N. L., Av. Gonzalitos 235 Norte, Monterrey, N. L., C. P. 64460.
Recibido/Received 23.03.2007. Aceptado/Accepted 15.08.2007.
Resumen. Karwinskia parvifolia (Kp) y Karwinskia humboldtiana (Kh) son plantas venenosas de la familia rhamnácea. Los estudios toxicológicos y clínicos realizados con uno de los compuestos presentes en el fruto [peroxisomicina A1 (PA1)) sugieren su posible utilidad farmacológica en el tratamiento de algunos tipos de cáncer. Esta es la razón por la cual estas plantas se cultivan para incrementar su productividad en PA1. Se investigó el efecto del nitrógeno y de la especie sobre la viabilidad de las semillas, y se determinó la curva de germinación para Kp con (grupo Kp-CN) y sin (grupo Kp-SN) nitrógeno, y para Kh sin nitrógeno (grupo Kh-SN) a través del modelo probit. Este modelo se caracteriza por la probabilidad de germinación en función del tiempo. Los resultados sobre promedios de cavidades con una o más plántulas por endocarpio fueron de 26,4 y 15,8 % para los grupos Kp- CN y Kp-SN, respectivamente, y 32,7 % en el grupo Kh-SN. Se observaron diferentes velocidades de germinación entre los grupos de plantas estudiados contenido y entre especies. El orden fue Kp-CN > Kp-SN > Kh-SN. El mayor de PA1 se obtiene de los endocarpios y testas de las semillas de Karwinskia, y los endocarpios con un mayor número de embriones tienen un mayor número de testas. Por lo tanto, a través de los porcentajes de germinación es posible estimar el número de embriones viables y por lo tanto el potencial de productividad de PA1, tan solo por tener un mejor llenado de grano. Como resultado, las pruebas de germinación serían una forma de obtener evidencia sobre el contenido embrionario de los endocarpios, y un parámetro a considerar para la preselección de plantas de alto rendimiento de PA1.
Palabras Clave: Viabilidad; Karwinskia parvifolia; Peroxisomicina A1; Nitrógeno.
Abstract. Karwinskia parvifolia (Kp) and Karwinskia humboldtiana (Kh) are poisonous plants of the rhamnacea family. Toxicological and clinical studies performed with one of the compounds present in the fruit [peroxisomicine A1 (PA1)) suggest its possible pharmacological usefulness in the treatment of certain types of cancer. This is why these plants are cultivated: to increase their productivity of PA1. We studied the effects of nitrogen and species on seed viability. The germination curve was determined for Kp with (Kp-WN) and without (Kp-NN) nitrogen, and for Kh without nitrogen (Kh-NN) using the probitmodel. This is characterized by the probability of germination in relation to time. The results of the average amount of cavities with one or more seedlings per endocarp were 26.4 and 15.8 % for the Kp-WN and Kp-NN groups, respectively; and 32.7 % for the Kh-NN group. Different germination speeds were observed between the study plants and between species, and the order was Kp-WN > Kp-NN > Kh-NN. The greatest amount of PA1 is obtained from the endocarps and testae of Karwinskiaseeds, and the endocarps with the greatest number of embryos have a greater number of testae. As a result, using the germination percentages it is possible to estimate the number of viable embryos and therefore the potential productivity of PA1, by only having better grain filling. Thus, the germination tests would be a way of obtaining evidence for the embryo content of the endocarps, and a parameter to consider for the selectivity of plants with high PA1 yield.
Key Words: Viability; Karwinskia parvifolia; Peroxisomicine A1; Nitrogen.
INTRODUCCIÓN
Hasta ahora la exploración de especies vegetales con fines medicinales se ha basado principalmente en plantas que de acuerdo con la cultura popular tienen propiedades curativas. No obstante lo anterior, hay otro grupo de plantas que también ha sido utilizado desde tiempos remotos, pero éste lo ha sido por sus propiedades tóxicas. Conocida la sentencia que la dosis hace al veneno, se podría decir que a ciertas dosis, las plantas conocidas como medicinales también podrían causar toxicidad. Por el contrario, las plantas conocidas como venenosas, a ciertas dosis podrían ser de utilidad medicinal. Tal es el caso de las plantas del género Karwinskia.
Las plantas de Karwinskiason perennifolias y producen severas intoxicaciones, parálisis y aun la muerte en animales y humanos que consumen sus partes vegetativas o frutos. El principio neurotóxico fue estudiado y reportado inicialmente por Dreyer et al. (1975). Estos autores identificaron cuatro sustancias sintetizadas por Karwinskia humboldtiana, a las que denominaron en función de su peso molecular como: T-496 (C30H24O7); T-514 (C30H26O8); T-516 (C30H28O8) y T-544 (C32H32O8). En años recientes se han identificado otros compuestos de Karwinskia parvifolia(Ramírez et al., 2002; Rivas et al., 2005).
Se han llevado a cabo diversos estudios toxicológicos y clínicos en relación a la T-514, sustancia que también ha sido reportada en la literatura como peroxisomicina A1 (PA1), y que ha mostrado citotoxicidad selectiva in vitro, con mayor intensidad en células tumorales (Garza et al., 1992; Piñeyro et al., 1994; Rivas y Waksman, 2001; Gómez et al., 2005).
Las investigaciones toxicológicas y clínicas se diversificaron, generando con ello una demanda creciente de PA1. Así se dio inicio al proceso de introducción y domesticación de plantas del género Karwinskia, por su posible utilidad contra algunos tipos de cáncer. Esto dio lugar al establecimiento de un vivero de Karwinskia, a partir del cual Luján et al. (1991, 1992, 1993a) reportaron la propagación por semilla y por injerto, así como la floración y fructificación de K. parvifoliay K. humboldtiana. También se observó en K. humboldtiana su moderada tolerancia al ataque de Phymatotrichum omnivorum, causante de la enfermedad conocida como "pudrición texana". Por esta razón se le utilizó como patrón en las pruebas de reproducción vegetativa de Karwinskiae injertó con K. parvifolia, especie que se observó altamente susceptible a "pudrición texana" (Luján et al., 1993b). Entre los resultados de éstos y otros estudios, se observó que en K. humboldtiana los frutos cosechados durante los meses de noviembre a abril produjeron el mayor peso seco por endocarpio. Sin embargo, el mismo fue menor durante los meses de agosto a septiembre (Luján, 1999).
Los resultados de diversos investigadores relacionados con el análisis y cuantificación de toxinas en Karwinskia[también ha sido reportado que PA1 posee actividad antimicrobiana (Salazar et al., 2006)], coinciden que su fruto es donde se han observado las concentraciones más elevadas de PA1 (Natham, 1987; Guerrero et al., 1987; Luján et al., 1995, Hanácková et al., 1997; Hanácková, 1999; Lisková et al., 1999; Piñeyro y Waksman, 2000).
En lo general, las especies de Karwinskia sintetizan diferentes tipos y cantidades de toxinas en frutos de origen silvestre y cultivado como una cualidad genética. Esto puede ser modificable por el ambiente, y bajo esa perspectiva resulta interesante conocer los aspectos agronómicos de las especies bajo estudio.
Está en curso una investigación en K. parvifolia donde se evaluó el efecto del nitrógeno y la variabilidad entre plantas sobre la productividad de PA1. Los resultados preliminares mostraron que las plantas producen frutos de diversa forma, tamaño y peso; el análisis químico sobre la concentración de PA1 sigue en curso (Luján et al., 2005, 2006).
Se consideró que a través de la viabilidad y velocidad de germinación de las semillas se podría estudiar el comportamiento individual de las plantas e incorporar esa información al conocimiento que ya se tiene sobre la domesticación de estas plantas. Esto permitiría ampliar el criterio de evaluación para su posible mejoramiento genético a través de la selección por alto rendimiento de PA1.
El fruto de Karwinskia es una drupa y en su endocarpio contiene hasta 4 embriones viables. Altos porcentajes de germinación indicarían un mayor contenido de embriones y por lo tanto mayor contenido de testas y de PA1, puesto que dentro del fruto, las testas son el sitio de mayor contenido de toxinas (Natham, 1987; Hanácková et al., 1997; Hanácková, 1999), entre las cuales se encuentra la T-514 (PA1).
Los objetivos de esta investigación fueron: 1) comparar los porcentajes de germinación en frutos cosechados de K. parvifoliacultivadas en vivero con y sin suplemento nitrogenado, y de K. humboldtianaversus K. parvifoliasin nitrógeno en ambas especies, y 2) caracterizar las curvas de germinación para K. parvifolia y K. humboldtiana.
MATERIALES Y MÉTODOS
Para las pruebas de germinación se utilizaron frutos de Karwinskia cosechados de plantas establecidas en 1991 en el vivero de Karwinskiadel Departamento de Farmacología y Toxicología de la Facultad de Medicina de la Universidad Autónoma de Nuevo León. Se diseñó un experimento para evaluar el efecto del nitrógeno y la variabilidad entre plantas sobre la productividad de PA1. Se utilizó un diseño cruzado estándar 2x2 [dos secuencias de tratamiento x dos ciclos de producción (2005 y 2006)] (Jones y Kenward, 1989). Para este experimento se seleccionaron 16 plantas de K. parvifoliay se formaron dos grupos de 8 plantas. Uno de estos grupos recibió N y el otro no. Como fertilizante nitrogenado se utilizó sulfato de amonio, [(NH4)2SO4] a razón de 2,44 kg por planta. El nitrógeno se aplicó e incorporó en una zanja de 15 cm de ancho x 15 cm de profundidad por dentro de la zona de goteo, distribuido en dos aplicaciones. El 70% se aplicó en la segunda quincena de diciembre de 2004, y el 30% restante en la segunda quincena de mayo de 2005. Después de cada fertilización se aplicaron riegos en todas las plantas, y fertilizados o no, éstos se repitieron de manera constante cada tres semanas.
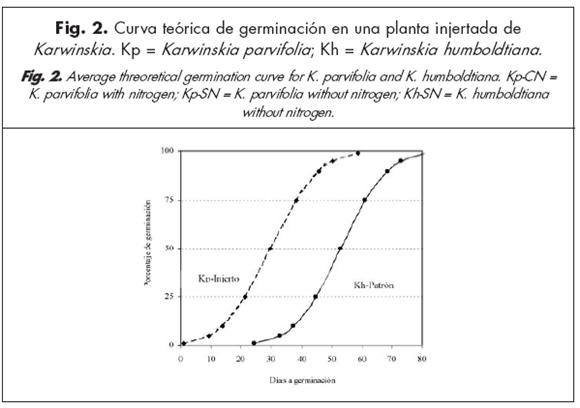
Las pruebas de germinación se realizaron en los invernaderos de la Facultad de Agronomía de la Universidad Autónoma de Nuevo León. Se trabajó con 20 plantas de Karwinskia, tomándose una muestra por planta de 100 endocarpios provenientes de frutos maduros. Dieciséis de las 20 muestras (1 a 16) se tomaron de plantas cosechadas durante septiembre y octubre de 2005; 8 de esas 16 muestras procedían de plantas con suplemento nitrogenado (grupo Kp-CN); mientras que las otras 8 muestras procedían de plantas sin dicho suplemento (grupo Kp-SN). También se utilizaron 4 muestras (17 a 20) cosechadas un día antes de la siembra (4 de agosto del 2006), tres de las cuales (18, 19 y 20) fueron de K. humboldtiana(grupo Kh-SN). Por otro lado, una planta de Kp se injertó en una planta de Kh, permitiéndose ambos crecimientos en un mismo sistema radical, etiquetándose como Kp el injerto (muestra 17) y como Kh el patrón (muestra 18). Se utilizaron 20 cajas de petri y 10 cajas de propagación de 200 cavidades cada una, usándose turba de musgo como sustrato.
En cada caja de propagación se colocaron e identificaron dos muestras de 100 endocarpios cada una y se prepararon con el sustrato (húmedo), colocándose un endocarpio por cavidad. Posteriormente, los endocarpios se cubrieron con una capa de turba de musgo de 5 milímetros de espesor, y se aplicó agua hasta saturar el sustrato. La siembra se realizó el 5 de agosto de 2006 y se llevó registro de datos desde esta fecha hasta el 18 de septiembre de 2006. En cada registro se tomaron los siguientes datos: 1) número de cavidades con emergencia de una, dos, tres o cuatro plántulas por endocarpio; 2) total de cavidades por muestra con emergencia de una o más plántulas por endocarpio y total de plántulas por muestra.
Las temperaturas que se presentaron entre las 8:30 y 10:30 horas durante el desarrollo de la prueba fluctuaron entre los 26.3 y 29.0 °C; mientras que la humedad relativa (HR) fluctuó entre 41.0 y 77.0 %. Por la tarde (12:30 a 14:30 horas), las temperaturas fluctuaron entre 43.9 y 47.6 °C; mientras que la HR fluctuó entre 17.0 y 24.0 %.
La medición de la velocidad de germinación se hizo a través de conteos periódicos después de la siembra. Para ello se contabilizó el número de plántulas por cavidad a los 4, 7, 10, 13, 16, 19, 21, 24, 26, 37, 38 y 44 días, considerando una curva sigmoidea como el prototipo de la germinación (Hartmann y Kester, 1979), también llamada curva de crecimiento de la población (Villee, 1985). Se utilizó el modelo de la curva probit (Salsburg, 1986), adaptado para caracterizar la curva de germinación en Karwinskia. Es decir: p ( germinación) = f ( tiempo), lo cual significa que la probabilidad de germinación estaría dada en función del tiempo.
Una vez obtenidos los datos generados a través de los conteos periódicos por cada muestra de 100 endocarpios, se procedió a obtener las pro babilidades teóricas de germinación en función del tiempo (días) y sus respectivos intervalos de confianza a través del modelo probit. Para ello se utilizó el paquete estadístico SPSS® 13.0.
Los porcentajes de germinación se compararon a través de tablas de contingencia 2x2 y pruebas de Ji-cuadrada ( x2 ). También se hizo un análisis de correlación en los materiales de K. parvifolia (muestras 1 a 16) para las variables: peso de endocarpio seco versus total de plántulas por muestra.
RESULTADOS
El peso promedio de endocarpio seco fue mayor en el grupo de plantas que recibieron suplemento nitrogenado que en el grupo de plantas que no lo recibieron (Tabla 1).

El total de cavidades en la caja de propagación con emergencia de 1 plántula por endocarpio fue de 181 para el grupo Kp-CN, 109 para el grupo Kp-SN y 92 para el grupo Kh-SN. En estos mismos grupos y en ese orden, se observaron otras 29, 16 y 2 cavidades con emergencia de 2 plántulas por endocarpio, respectivamente. La emergencia de 3 plántulas por endocarpio se observó en 1 cavidad sólo en los grupos Kp-CN y Kp-SN. En el grupo Kh-SN no se observaron cavidades con 3 plántulas. En ninguna cavidad de las 10 cajas de propagación se observó la emergencia de 4 plántulas por endocarpio.
Los valores de emergencia ya informados totalizaron 242 [(181 cavidades x 1=181) + (29 cavidades x 2=58) + (1 cavidad x 3=3)], 144 y 100 plántulas para los grupos Kp-CN, Kp-SN y Kh-SN, respectivamente.
Por otro lado, los resultados promedio de cavidades por muestra con emergencia de una o m ás plántulas por endocarpio fueron de 26,4 y 15,8 % por grupo de plantas de K. parvifoliacon nitrógeno y sin nitrógeno, respectivamente (Tabla 2). En la comparación entre ambas proporciones resultó una x2 (Jí-cuadrada) = 27,16. Esto indica que los porcentajes de germinación entre ambos tratamientos son estadísticamente diferentes.

En el grupo de plantas de K. humboldtiana(sin nitrógeno) la germinación promedio fue de 32,7 % (Tabla 2). Este valor, comparado con el 15,8 % de germinación de las muestras de K. parvifolia (sin nitrógeno), permitió obtener una x 2 = 38,5. Esto indica que las diferencias entre especies fueron estadísticamente significativas, siendo mayor el porcentaje de germinación en K. humboldtiana.
El análisis de correlación ( r ) entre las variables: peso de endocarpio seco por muestra versus total de plántulas emergidas por muestra resultó en un r = 0,74 ( n = 16). Este resultado indicaría que el mayor peso seco por endocarpio obtenido en las plantas que recibieron suplemento nitrogenado influyó positivamente sobre los porcentajes de germinación observados en K. parvifolia.
En las muestras obtenidas del injerto de K. parvifolia(muestra 17), se obtuvieron porcentajes de germinación de 18 %. Al mismo tiempo, en la muestra 18 de la planta patrón ( K. humboldtiana) se observó un 17 % de germinación. En estas 2 muestras no se observaron cavidades con emergencia de 2 o más plántulas por endocarpio.
En cuanto a la velocidad de germinaci ón entre especies, (muestras 1 a 16) se observó que el período de mayor crecimiento poblacional de plántulas de K. parvifolia estuvo comprendido entre los días 4 al 26 a partir de la siembra.
Para el mismo per íodo de los conteos en K. parvifolia, en K. humboldtiana se observaron 2, 3 y 6 plántulas en las muestras 18, 19 y 20, respectivamente. Sin embargo, a partir de esa fecha la emergencia de plántulas se intensificó en estas 3 muestras, y se prolongó hasta prácticamente los 44 días después de la siembra.
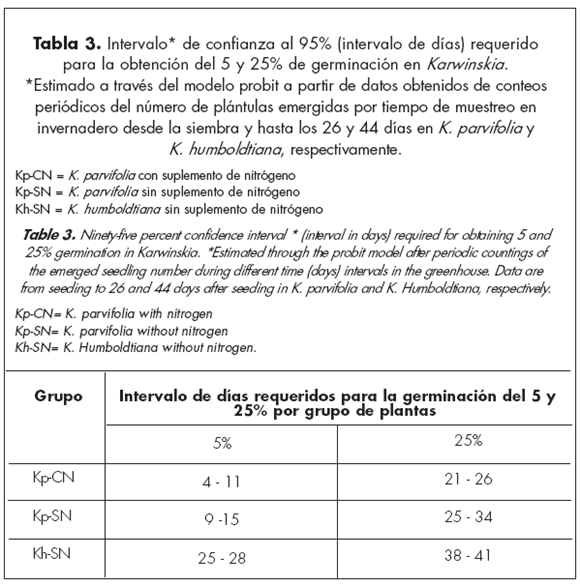
En las Figs. 1 y 2 se observa que para obtener un 100 % de germinaci ón serían necesarios más de 60 días. Esto no ocurrió en esta investigación. Tal situación sólo se obtendría suponiendo que todos los endocarpios tuviesen embriones viables y que los mismos no fuesen afectados por plagas, enfermedades, temperaturas extremas, humedad u otra adversidad ambiental. Habría que suponer además que la velocidad de germinación observada desde la siembra y hasta los 26 y 44 días, para K. parvifolia y K. humboldtiana, respectivamente, se mantuviese sin cambios por tiempo indefinido. Sin embargo, a partir de los datos de los conteos periódicos realizados en el invernadero y por medio del modelo probit, fue posible caracterizar las curvas teóricas desde el 1 y hasta el 99 % de la germinación para cada uno de los grupos estudiados y considerando los resultados promedio de germinación en los tres grupos de plantas. Se obtuvo así el número de días promedio y los intervalos de confianza al 95 % que serían requeridos para dos valores teóricos de referencia seleccionados: el 5 y 25 % de germinación.

Los resultados muestran que en el grupo de plantas de K. parvifolia que recibieron suplemento nitrogenado y que produjeron un 26,4 % de germinación, teóricamente se requerirían intervalos de 4 a 11 y de 21 a 26 días para la obtención del 5 y 25 % de germinación, respectivamente. En el grupo de plantas de K. parvifoliaque no recibió suplemento nitrogenado y que produjo 15,8 % de germinación, se requerirían en teoría, intervalos de 9 a 15 y de 25 a 34 días para la germinación del 5 y 25 %, respectivamente. En K. humboldtiana, que produjo un 32,7 % de germinación, teóricamente se requerirían intervalos de 25 a 28 y de 38 a 41 días para la germinación del 5 y 25 %, respectivamente (Fig. 1, Tabla 3).
DISCUSIÓN
Se observaron diferencias entre especies respecto a la velocidad de germinación, aún cuando los endocarpios provinieron de frutos cosechados el mismo día y de una misma planta (Fig. 2), donde K. humboldtiana funcionó como patrón y K. parvifolia como injerto. Esto indicaría que ambas especies presentan diferentes velocidades de germinación aún cuando compartan el mismo sistema radical (Fig. 3a).

En otras especies como las cerezas dulce y agria, el período de germinación de todas las semillas se prolonga por 15 y 18 semanas, respectivamente (Jankiewicz, 2003). Una de las causas de estos períodos prolongados es que una especie de árbol puede tener semillas tanto latentes como no latentes, o puede tener más de un tipo de latencia (USDA, 1986). El hecho que las semillas frecuentemente tienen un período de latencia en el cual no pueden germinar aún cuando las condiciones ambientales son favorables para el desarrollo, previene la disminución de la población de plántulas por cambios ambientales adversos (Jankiewicz, 2003). En las especies de Karwinskia, esto podría ser una característica de adaptación.
Epstein y Bloom (2005) mencionan que pr ácticamente cada una de las especies tiene su propia forma especial de almacenamiento de proteínas con diferentes vías catabólicas, y se considera que esto le da ventaja contra predadores. Es posible que la síntesis de toxinas como las mencionadas para Karwinskia, así como los períodos de latencia y la velocidad de germinación, sean un ejemplo de esas ventajas. Es probable que esto favorezca en mayor o menor grado la propia adaptación de las diferentes especies de Karwinskia.
En cuanto a los porcentajes de germinaci ón, se sembraron 800 endocarpios del grupo Kp-CN; 800 del grupo Kp-SN y 300 del grupo Kh-SN, de los cuales se obtuvieron totales de 242; 144 y 100 plántulas, respectivamente. Sin embargo, si consideramos que cada endocarpio podría generar hasta 4 embriones viables, entonces los resultados esperados serían de 3200; 3200 y 1200 plántulas, respectivamente. Esto significa que la cantidad de plántulas obtenidas con respecto a las potencialmente esperadas para cada uno de los grupos fue de 7,6 (242/3200*100); 4,5 (144/3200*100) y 8,3 % (100/1200*100), respectivamente. En semillas forestales el número de plántulas producidas por 100 semillas viables ha variado entre 10 y 60 para coníferas, y varía aún más en especies de hoja ancha (USDA, 1986).
Las semillas con baja viabilidad por lo general se asocian con bajos porcentajes y velocidades de germinaci ón (Hartmann y Kester, 1979). Lillo (2004) menciona que algunas veces un suplemento nitrogenado abundante contrarresta la transición hacia la latencia. Por otro lado, la escasez de nitrógeno puede inducir latencia. Masarovicová et al. (2000) investigaron el efecto del nitrógeno sobre la producción de peroxisomicina A1 en plantas de K. parvifolia y K. humboldtiana. Estos autores informaron que el suplemento de nitrógeno influyó significativamente sobre la concentración de peroxisomicina A1 en las hojas de ambas especies. Considerando la alta correlación lineal obtenida entre el peso del endocarpio y el total de plántulas producidas, los porcentajes de germinación obtenidos serían una demostración del efecto del suplemento nitrogenado. Además, estaría indicando que los endocarpios más pesados tendrían mayor volumen seminal y/o mayor número de embriones viables por endocarpio, lo cual se traduciría potencialmente en mayores porcentajes de germinación.
Por otro lado, si se considera que en Karwinskia se pueden generar hasta 4 plántulas por endocarpio, entonces los porcentajes de germinación serían una forma de estimar el número de embriones viables (Fig. 3b). A su vez, esto daría una indicación del potencial de productividad de PA1, tan solo por efecto de tener un mejor llenado de grano. Los costos de extracción y purificación fluctúan alrededor de los 1000 dólares americanos por gramo de PA1 sin incluir costos de equipo (Lisková et al., 1999). Por lo tanto, la obtención de endocarpios con 4 embriones bien desarrollados tendría un importante efecto sobre la productividad de PA1. Además, las pruebas de germinación serían una forma de obtener evidencia sobre el contenido embrionario de los endocarpios y un parámetro a considerar para la preselección de plantas de alto rendimiento de PA1.

REFERENCIAS
1. Dreyer, D., I. Arai, C. D. Bachman, W.R. Anderson, R. G. Smith y G. D. Daves (1975). Toxins Causing Noninflammatory Paralytic Neuronophaty. Isolation and Structure Elucidation. Journal of the American Chemical Society 97: 4985-4990. [ Links ]
2. Epstein, E. y A. J. Bloom (2005). Mineral Nutrition of Plants. Principles and perspectives. 2ª ed. Sinauer Associates, Inc. Publishers, Massachusets. 400 p. [ Links ]
3. Garza-Ocañas, L., G.C. Hsieh, D. Acosta, O. Torres-Alanís y A. Piñeyro-López (1992). Toxicity assessment of toxins T-514 and T-544 of Buckthorn ( Karwinskia humboldtiana) in primary skin and liver cell cultures. Toxicology73: 259-267. [ Links ]
4. Gómez-Silva, M., L. Garza-Ocañas, N. Waksman, V. Rivas y A. Piñeyro-López (2005). In vitro metabolism and toxicity assessment of toxin T-514 (Peroxisomicine A1) of Karwinskia humboldtianain microsomes and primary cultured hepatocytes. Toxicology in Vitro19: 47-53. [ Links ]
5. Guerrero, M., A. Piñeyro y N. Waksman (1987). Extraction and quantification of toxins from Karwinskia humboldtiana(tullidora). Toxicon 25: 565-568. [ Links ]
6. Hanácková, Z., N. Waksman, M. Mikus, A. Lux, D. Lisková y E. Masarovicová (1997). Structure, physiology and toxin content in leaves of Karwinskia humboldtiana in vivo and in vitro. Physiologia Plantarum29: 95-100. [ Links ]
7. Hanácková, Z. (1999). Ontogénesis de las estructuras internas y algunas características anatómicas y embriológicas de Karwinskia parvifolia (Rhamnáceae). Tesis Doctoral, Facultad de Ciencias Naturales, Universidad Comeniana, Bratislava, Eslovaquia. 65 p. [ Links ]
8. Hartmann, H.T. y D.E. Kester (1979). Propagación de plantas. Tr. Antonio Marino Ambrosio. 1ª edición de la 2ª edición en inglés. Ed. Cecsa. 810 p. [ Links ]
9. Jankiewicz, L.S. (2003). Desarrollo de semillas y la presencia de latencia en yemas y semillas; el papel de los reguladores de plantas. En: Jankiewicz, L.S. (ed), pp. 193-240. Reguladores del crecimiento, desarrollo y resistencia en plantas. Vol. I. Propiedades y acción. Ediciones Mundi- Prensa, México. 487 p. [ Links ]
10. Jones, B. y M.G. Kenward (1989). Design and Analysis of Cross-Over Trials. Editorial Chapman & Hall/CRC, London, England. 340 p. [ Links ]
11. Lillo, C. (2004). Light regulation of nitrate uptake, assimilation and metabolism. En: Amancio, S. & I. Stulen (eds), pp 149-184. Nitrogen Acquisition and Assimilation in Higher Plants. Ed. Kluwer Academic Publishers, The Netherlands. 299 p. [ Links ]
12. Lisková, D., A. Lux, A. Piñeyro L., y R. Luján R. (1999). Karwinskia species (Buckthorn): in vitro culture, micropropagation, and the production of toxin anthracenone. En: Y. P. S. Bajaj (ed), pp. 223-242. Biotechnology in Agriculture and Forestry, Medicinal and Aromatic Plants XI. Vol. 43. Springler-Verlag, Berlin, Heidelberg. [ Links ]
13. Luján, R. R., R. González y A. Piñeyro L. (1991). Propagación del género Karwinskia. Memorias del IX Encuentro Regional de Investigación Biomédica, Facultad de Medicina, U. A. N. L., Monterrey, N. L., México. p. 6. [ Links ]
14. Luján, R. R., R. González y A. Piñeyro L. (1992). Propagación del género Karwinskia. Memorias del X Encuentro Regional de Investigación Biomédica. Facultad de Medicina de la U. A. N. L., Monterrey, N. L., México. pp. 181-182. [ Links ]
15. Luján, R. R., R. González y A. Piñeyro L. (1993a). Karwinskia humboldtiana y su fructificación. Memorias del XI Encuentro Regional de Investigación Biomédica. Facultad de Medicina de la U. A. N. L., Monterrey, N. L., México. p. 96. [ Links ]
16. Luján, R. R., R. González y A. Piñeyro L. (1993b). Enfermedades del género Karwinskia. Memorias del XI Encuentro de Investigación Biomédica. Facultad de Medicina de la U. A. N. L., Monterrey, N. L., México. p. 112. [ Links ]
17. Luján, R. R., N. Waksman, M. L. Salazar y A. Piñeyro L. (1995). Producción de antracenonas en varias especies de Karwinskia bajo diferentes condiciones de cultivo. Memorias del XIII Congreso Nacional de Investigación Biomédica. Facultad de Medicina de la U. A. N. L., Monterrey, N. L., México. p. 20. [ Links ]
18. Luján, R. R (1999). Efecto del período de cosecha, tamaño del fruto, riego y especie sobre el contenido de Peroxisomicina A1 (T-514) en plantas del género Karwinskia. Tesis de Maestría. Facultad de Agronomía, U. A. N. L., Marín, N. L., México.115 p. [ Links ]
19. Luján-Rangel, R., E. Olivares-Sáenz, R. E. Vázquez-Alvarado, L. Garza-Ocañas y O. Torres-Alanís (2005). Efecto del nitrógeno sobre la producción de T-514 en plantas del género Karwinskia. Memorias del XXIII Ciclo de Seminarios. División de Estudios de Posgrado, Facultad de Agronomía-UANL., Marín, N. L., México. [ Links ]
20. Luján-Rangel, R., E. Olivares-Sáenz, R. E. Vázquez-Alvarado, L. Garza-Ocañas y O. Torres-Alanís (2006). Producción fitoquímica y aplicación de nitrógeno en Karwinskia. Memorias del XXIV Ciclo de Seminarios. División de Estudios de Posgrado, Facultad de Agronomía-UANL., Marín, N. L., México. [ Links ]
21. Masarovicová, E., R. Welschen, A. Lux, H. Lambers, K. Argalásová, E. Brandsteterová y A. Caniova (2000). Photosynthesis, biomass partitioning and peroxisomicine A1 production of Karwinskia species in response to nitrogen supply. Physiologia Plantarum108: 300-306. [ Links ]
22. Natham, P.J. (1987). Revista de la Academia de Ciencias y Artes de Puerto Rico2: 19-23. [ Links ]
23. Piñeyro-López, A., L. Martínez de Villarreal y R. González-Alanís (1994). Toxicology92: 217-227. [ Links ]
24. Piñeyro-López, A. y N.Waksman (2000). Chemistry, Structure and Biological Activity of Anthracenones of the Karwinskia Genus. En: Atta-ur-Rahman (ed), pp. 555-606. Natural Products Chemistry. V 22. Elsevier Science. [ Links ]
25. Ramírez-Durón, R., A. García-Luna, L. Garza-Ocañas, A. Piñeyro-López y N. Waksman de Torres (2002). Pharmaceutical Biology40: 440-447. [ Links ]
26. Rivas, V. y N. Waksman (2001). Natural Product Letters15: 243-251. [ Links ]
27. Rivas, V., G. Cuevas, L. Garza y N. Waksman (2005). Arkivoc 12: 224-233. [ Links ]
28. Salazar, R., V. Rivas, G. González, y N. Waksman (2006). Fitoterapia77: 398-400. [ Links ]
29. Salsburg, D.S. (1986). Statistics for Toxicologists. Marcel Dekker, New York, NY, USA. 196 p. [ Links ]
30. USDA (1986). Semillas. Compañía Editorial Continental, S. A., México. 1020 p. [ Links ]
31. Villee C.A. (1985). Biología. 7a ed. Nueva Editorial Interamericana, S. A., México D. F. 803 p. [ Links ]

