










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkPhyton (Buenos Aires)
On-line version ISSN 1851-5657
Phyton (B. Aires) vol.78 no.1 Vicente López Jan./June 2009
ARTÍCULOS ORIGINALES
Nitrogen metabolism in roots and leaves of green bean plants exposed to different phosphorus doses
Metabolismo del nitrógeno en raíces y hojas de plantas de frijol ejotero expuestas a diferentes dosis de fósforo
Sánchez E.1, G. Ávila-Quezada1, A.A. Gardea1, E. Muñoz1, J.M. Ruiz2, L. Romero2
1Centro de Investigación en Alimentación y Desarrollo, A.C. Av. 4ta. Sur 3820, Fracc. Vencedores del Desierto. Cd. Delicias, Chihuahua. México. 33089. Tel.: +52 (639) 4748400. Fax: +52 (639) 4748704.
2 Departamento de Fisiología Vegetal, Facultad de Ciencias, Universidad de Granada, 18071-Granada, España.
Address Correspondence to: Esteban Sánchez-Chávez; e-mail: esteban@ciad.mx
Recibido-Received 2.09.2008.
Aceptado-Accepted 14.10.2008.
Abstract. The objective of this work was to determine the effect of different P doses on nitrogen assimilation in roots and leaves of green beans plants (Phaseolus vulgaris L. cv. Strike). Phosphorus was applied in the nutrient solution as H3PO4, at the nutrient doses of: P1 = 0.40 mM; P2 = 0.80 mM; P3 = 1.60 mM; P4 = 3.20 mM; P5 = 4.80 mM, and P6 = 6.40 mM P. Our results indicate that both P toxicity and deficiency gave similar responses to N assimilation. Phosphorus and NO3 - interacted on the absorption and translocation processes affecting N assimilation. The deficiency (P1), and toxicity (P6) treatments, diminished root absorption of NO3- in 15% and 36%, respectively, respect to the optimum dose (P3), thus reducing nutrient availability for assimilation. This result may explain the minimum enzymatic activities observed in NO3 - assimilation in P1 and P6. The minimum N assimilation observed in these treatments will eventually translate into a minimum synthesis of major N organic compound involved in plant growth and development. This will lead to a reduced plant biomass production and productivity of green beans plants.
Key words: Phaseolus vulgaris L.; Nitrogen metabolism; Ggreen bean; Phosphorus; Deficiency; Toxicity.
Resumen. El objetivo de este trabajo fue determinar el efecto de diferentes dosis de fósforo sobre la asimilación de nitrógeno en raíces y hojas de frijol ejotero Phaseolus vulgaris L. cv. Strike). El fósforo fue aplicado a la solución nutritiva en la forma de H3PO4; se usaron las siguientes dosis: P1 = 0.40 mM; P2 = 0.80 mM; P3 = 1.60 mM; P4 = 3.20 mM; P5 = 4.80 mM y P6 = 6.40 mM de P. Nuestros resultados indican que tanto los tratamientos de deficiencia y toxicidad de P presentaron respuestas similares en la asimilación de N. Estos resultados definen la interacción P-NO3- que se produce a nivel de absorción y movilización como el principal efecto que ejerce este macronutriente sobre la asimilación de N. Tanto la deficiencia (P1) como la toxicidad (P6) de P redujeron la absorción radical de NO3- en un 15% y 36% respectivamente en relación a la dosis óptima (P3), reduciendo así la disponibilidad de NO3- para ser asimilado. Nuestros resultados explicarían las actividades enzimáticas mínimas observadas para la asimilación de NO3- en P1 y P6. La mínima asimilación de N que se produce en estos tratamientos se traducirá eventualmente en una mínima síntesis de los principales compuestos nitrogenados orgánicos implicados en el crecimiento y desarrollo vegetal. Finalmente, esto conducirá a una menor producción de biomasa y productividad en plantas de frijol ejotero.
Palabras clave: Phaseolus vulgaris L.; Metabolismo nitrogenado; Ffrijol ejotero; Ffósforo; Deficiencia; Toxicidad.
INTRODUCTION
Beans are grown and consumed in nearly all the world. In many developing countries, 20% of the available protein is provided by beans. Beans represent also an integral part of dietary protein for 50% of the world's population (Deshpande et al., 1984). Beans are produced in large quantities in the American Continent and East Africa (Singh, 1999).
Phosphorus (P) is the second nutritional element after nitrogen that limits plant growth, having a concentration of about 0.2% of the total plant dry weight (Jeschke et al., 1996; Raghothama, 1999). This macronutrient is a key component in many molecules (i.e., nucleic acids, phospholipids and ATP) that participate in basic plant processes (Schachtman et al., 1998). In its inorganic form (Pi) it is involved in the control of many enzymatic reactions, which regulate different metabolic processes (Theodorou & Plaxton, 1993). Main effects of P deficiency include (1) the reduction in leaf number (Lynch et al., 1991) and (2) loss of photosynthetic efficiency (Lauer et al., 1989).
Nitrogen (N) and P are intimately involved in plant metabolism and growth: they interact in different biochemical processes. N participates directly in amino acid, protein, nucleic acid and other cellular component syntheses, which are needed for plant growth and development. N assimilation by plants requires NO3- absorption, its reduction and subsequent conversion to NO2- , and NH4+, respectively, and the final NH4+ incorporation into organic compounds (Sivasankar & Oaks, 1996; Stitt, 1999). Factors influencing the enzymatic regulation responsible for N assimilation include: plant phenological stage (Ireland & Lea, 1999); light-darkness periods (Migge et al., 1996); sucrose concentration (Lam et al., 1996); N source: NO3- and NH4+ (Ruiz & Romero, 1999); CO2 level (Edwards & Coruzzi, 1989); temperature (Woodall et al., 1996); nutrients (Lopez-Lefebre et al., 2000); growth regulators (Ruiz et al., 2000), products of nitrogen assimilation (Padget & Leonard, 1996) and genetic variability (Ruiz & Romero, 1998).
Phosphorus role in N metabolism has been studied in detail, mostly related to its deficiency. Some of the effects of P deficiency on NO3- assimilation are: (1) reduction of NO3- absorption by roots (Rufty et al., 1991; Pilbeam et al., 1993; Jeschke et al., 1997). This might be likely caused by limited root ATP availability and limitations in the transport membrane system to NO3- absorption (Rufty et al., 1993); (2) Decline in NO3- translocation from roots to shoots (Rufty et al., 1991; Pilbeam et al., 1993; Jeschke et al., 1997). This is associated with the decrease of water pressure through roots and xylem (Rufty et al., 1991; Rufty et al., 1993; Jeschke et al., 1997). All these factors may diminish nitrate reductase activity, and therefore cause a decrease of NO3- assimilation.
In general, N assimilation is affected when P-deficient plants are supplied with NO3- . However, the regulatory mechanism associated with N assimilation under nutrient (P) stress has not been completely clarified. The objective of this study was to determine the effect of different P doses on nitrogen assimilation of roots and leaves in green beans plants (Phaseolus vulgaris L. cv. Strike).
MATERIALS AND METHODS
Crop management and experimental design. Seeds of Phaseolus vulgaris cv. Strike were sown and grown in a growth chamber under controlled environmental conditions: 60-80% relative humidity, 30/20°C (day/night) temperature and 16/8 h photoperiod under a photosynthetic photon flux density of 350 μmol/m2/s (measured at the top of the plants with a 190 SB quantum sensor, LI-COR Inc., Lincoln, NE). Four plants were grown in 8 l pots (25 cm upper diameter, 17 cm lower diameter, 25 cm height), filled with vermiculite. During 20 days, before the experimental treatments, all plants received a nutrient solution consisting of 6 mM NH4NO3; 1.6 mM H3PO4; 4.0 mM K2SO4; 4 mM CaCl2· 2H2O; 1.4 mM MgSO4· H2O; 5 μM Fe-EDDHA; 2 μM MnSO4· H2O; 1 μM ZnSO4· 7H2O; 0.25 μM CuSO4· 5H2O; 0.3 μM Na2MoO4· 2H2O; and 25 μM H3BO3. The nutrient solution (pH= 6.0 ± 0.1) was renewed every 3 days.
Twenty days after sowing, different P treatments were applied as H3PO4 during 40 days (until harvest): P1: 40 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM, and P6: 6.40 mM). The optimal N dose for Phaseolus vulgaris under our experimental conditions was the P3 treatment; this result is similar to previous studies (Sánchez et al., 2004). A completely randomized block experimental design was used with 6 replicates (individual pots: 24 plants in each of them) per treatment.
Sampling and plant analysis. Plants were sampled 60 days after sowing, at full pod development and maturity. Roots and leaves were sorted out for analysis. Plant material was first rinsed three times in distilled water after disinfecting with a non-ionic detergent at 1% (Wolf, 1982) and then blotted on filter paper. A fresh subsample of roots and leaves was used for analysis of nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), glutamate synthase (GOGAT), phosphoenolpyruvate carboxilase (PEPC), amino acids and proteins. These subsamples were dried in a forced air oven at 70°C for 24 h, ground in a Wiley mill and placed in plastic bags until analysis for nitrate (NO3- ) and ammonium (NH4+ ). Dry weights (DW) were recorded and expressed as mg DW per root or leaf. All determinations were performed in triplicate.
Statistical analysis. Data were analyzed using ANOVA. When F tests were significant, differences between treatment means were compared using LSD at the 0.05 probability level. Also, correlation analyses were made between the different variables. Levels of significance were represented by * at p<0.05, ** at p<0.01, *** at p<0.001, and NS: not significant. Data shown are mean values ± SE.
RESULTS AND DISCUSSION
Phosphorus is an important nutritional element that may limit plant growth (Raghothama, 1999). In our experiment, P treatments affected root biomass production (Fig. 1). Leaf biomass production was also influenced by increasing phosphorus concentrations. P3 showed the highest leaf biomass, with a value 15% greater than that in P6 (Fig. 1); while root biomass production attained its highest value in P1, with an increment of 61% in relation to P6, P3 was considered optimum for greater root and foliar biomass production than P6. This result coincides with those of Carbonell-Barrachina et al. (1997). Treatments below P3 concentrations (i.e., P1 and P2) can be considered P deficient, having a negative effect on leaf biomass production. Doses higher than P3 may have caused P toxic effects, which diminished production of both leaf and root bean biomasses.

Fig. 1. Root and foliar biomass in response to H3PO4 treatments (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM and P6: 6.40 mM P) in green bean plants. Data are means + 1 s.e. (n = 6).
Fig. 1. Biomasa radical y foliar en plantas de frijol ejotero en respuesta a los tratamientos de H3PO4 (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM y P6: 6.40 mM de P). Los datos son promedio + 1 e.e. (n =6).
In general, N assimilation is disturbed when plants growing with an adequate NO3- concentration are limited by inadequate P levels (Rufty et al., 1993). Three different adverse effects have been identified in this regard: (i) diminished NO3- root absorption (Rufty et al., 1991); (ii) reduction of NO3- translocation from roots to shoots, with a subsequent NO3- concentration increment in roots (Lee, 1982; Rufty et al., 1990); and (iii) increased amino acid accumulation in leaves (Israel & Rufty, 1988). Accumulation of amino acids has also been shown in roots as a rare event (Rabe & Lovatt, 1984; Rufty et al., 1990).
The effect of the different P doses on NO3- concentrations in roots and leaves are shown in Fig. 2. The maximum NO3- concentration was observed in P3. This concentration was 29% and 31% greater than concentrations on P1 and P6, respectively. These findings are similar to those reported by Lee (1982) and Rufty et al. (1990). P limitation produces a reduced both NO3- absorption and translocation to leaves, causing a NO3- accumulation in roots (Fig. 2). Rufty et al. (1993) showed that the diminished root NO3- absorption and its subsequent translocation to leaves are probably caused by a drastic decrease in the Pi and ATP concentrations, which are essential in the NO3- absorption and transportation processes.

Fig. 2. Root and foliar NO3- concentrations in response to H3PO4 treatments (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM and P6: 6.40 mM P) in green bean plants. Data are means + 1 s.e. (n = 6).
Fig. 2. Concentración de nitratos radical y foliar en plantas de frijol ejotero en respuesta a los tratamientos de H3PO4 (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM y P6: 6.40 mM de P). Los datos son promedio + 1 e.e. (n =6).
Higher than optimum P doses showed a reduction in root and foliar NO3- concentrations. This may be due, at least partially, to the antagonism between PO4= and NO3- absorption (Marschner, 1995).
Although statistically not significant (p>0.05), both deficient and toxic P doses resulted in negative effects on root and leaf NH4+ concentrations (Fig. 3).
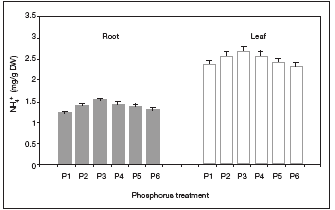
Fig. 3. Root and foliar NH4+ concentrations in response to H3PO4 treatments (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM and P6: 6.40 mM P) in green bean plants. Data are means + 1 s.e. (n = 6).
Fig. 3. Concentraciones de NH4+ radical y foliar en plantas de frijol ejotero en respuesta a los tratamientos de H3PO4 (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM y P6: 6.40 mM de P). Los datos son promedio + 1 e.e. (n =6).
The role of P in N assimilation has been widely studied, although mostly in experiments related to P deficiency. The results obtained so far as in our study, indicate that NO3- assimilation diminishes with limited P availability. Nitrate absorption and translocation have been strongly affected curtailing root and shoot biomass accumulation (Rufty et al., 1991).
Regarding the enzyme responses influencing NO3- reduction (i.e., NR and NiR), we found that the activity of both enzymes was similar to the trend of NO3- concentrations observed in roots and leaves (Table 1): P3 showed the highest values and the enzymatic activity increased more than 25% in both organs compared to P1 and P6. The direct relationship between both parameters (P concentration versus enzyme activity) is reflected in the high correlation coefficients obtained in different study plant parts (roots: NO3- -NR, r = 0.94***; NO3- -NiR, r = 0.90***; leaves: NO3- -NR, r = 0.84**; NO3- -NiR, r = 0.80**; **p<0.01, ***p<0.001).
Table. 1. Effect of H3PO4 treatments (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM and P6: 6.40 mM P) on the key enzymes of nitrogen metabolism in roots and leaves of green bean plants. Data are means ± 1 s.e. (n = 6).
Tabla. 1. Efecto de los tratamientos de H3PO4 (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM y P6: 6.40 mM de P) sobre las enzimas claves de la asimilación de nitrógeno en raíces y hojas de plantas de frijol ejotero. Los datos son promedio ± 1 e.e. (n =6).

The GS/GOGAT activities showed a similar behavior, both in roots and leaves, as the activity shown by the NR and NiR enzymes. The maximum activities for GS and GOGAT were also observed in the optimum treatment (P3): increases were over 30% in roots and 35% in leaves compared to the minimum concentrations obtained in the extreme treatments (P1 and P6).
Several research works have shown that the PEPC activity is stimulated under P deficiency conditions (Kondracka& Rychter, 1997). However, Biddinger et al. (1998) found that lack of P diminishes CO2 assimilation and, therefore, PEPC is reduced as well. Results of the current experiment (Table 1) are in agreement with this statement. The maximum activities of PEPC were obtained in P3 in both roots and leaves, while the minimum values were found in P1 and P6.
In relation to high molecular weight N compounds, such as amino acids and proteins (Table 2), the concentration in roots and leaves followed a similar behavior to all study enzymes activities. While P3 showed the highest concentrations of these compounds, the minimum was observed in P1 and P6.
Table. 2. Amino acid and protein accumulation in roots and leaves of green bean plants in response to H3PO4 treatments (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM and P6: 6.40 mM P). Data are means ± 1 s.e. (n = 6).
Tabla. 2. Acumulación de aminoácidos y proteínas en raíces y hojas de frijol ejotero en respuesta a los tratamientos de H3PO4 (P1: 0.4 mM; P2: 0.80 mM; P3: 1.60 mM; P4: 3.20 mM; P5: 4.80 mM y P6: 6.40 mM de P). Los datos son promedio ± 1 e.e. (n =6).

CONCLUSION
Results proved that both P toxicity and deficiency gave similar responses in N assimilation. Phosphorus and NO3- interacted on the absorption and translocation processes affecting N assimilation. The deficiency (P1) and toxicity (P6) treatments diminished root NO3- absorption by 15% and 36%, respectively, in comparison to P3, the optimum dose. These treatments also reduced nutrient assimilation within the plant, which may help explain the minimum enzymatic activity for NO3- assimilation observed in P1 and P6. The minimum N assimilation shown in these treatments, and the subsequent minimum synthesis of N organic compounds, which are keys to plant growth and development, will influence the reduction of green bean plant biomass and production.
REFERENCES
1. Biddinger, E.C., C. Liu, R.J. Joly & K.G. Raghothama (1998). Physiological and molecular responses of aeroponically grown tomato plants to phosphorus deficiency. Journal of the American Society for Horticultural Sciences 123: 330-333. [ Links ]
2. Bradford, M.M. (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dyebinding. Annals of Biochemistry 72: 248-254. [ Links ]
3. Cataldo, D.A., M. Haroon, L.E. Schrader & V.L. Young (1975). Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Communications in Soil Science and Plant Analysis 6: 71-80. [ Links ]
4. Deshpande, S., S.K. Styhe & D.K. Salunkhe (1984). Interrelationships between certain physical and chemical properties of dry bean. Quality Plant Foods Human Nutritional 34: 53-65. [ Links ]
5. Edwards, J.W. & G.M. Coruzzi (1989). Photorespiration and light act in concert to regulate expression of the nuclear gene for chloroplast and cytosolic glutamine synthetase. Plant Cell 1: 241-248. [ Links ]
6. Farnden, K.J.F. & J.G. Robertson (1980). Methods for studying enzymes envolved in metabolism related to nitrogenase. In: Bergersen, F.J. (ed.), pp: 265-314. Methods for Evaluating Biological Nitrogen Fixation. John Wiley& Sons, New York. [ Links ]
7. Geladopoulus, T.P., T.G. Sotiroudis & A.E. Evangelopoulus (1991). A Malachite green colorimetric assay for protein phosphatase activity. Annals of Biochemistry 192: 112-116. [ Links ]
8. Groat, R.G. & C.P. Vance (1981). Root nodule enzymes of ammonia assimilation in alfalfa (Medicago sativa L.). Plant Physiology 32: 169-204. [ Links ]
9. Hageman, R.H. & D.P. Hucklesby (1971). Nitrate reductase. Methods in Enzymology 23: 497-503. [ Links ]
10. Ireland, R.J. & P.J. Lea (1999). The enzymes of glutamine, glutamate, asparagina and aspartate metabolism. In: Singh, B.K. (ed.), pp. 49-109. Plant amino acids. Biochemistry and Biotechnology. Marcell Dekker, Inc., New York. [ Links ]
11. Israel, D.W. & J.R. Rufty (1988). Influence of phosphorus nutrition on phosphorus and nitrogen utilization efficiencies and associated physiological responses in soybean. Crop Science 28: 954-960. [ Links ]
12. Jeschke, W.D., E.A. Kirkby, A.D. Peuke, J.S. Pate & W.C. Hartung (1997). Effects of P deficiency on assimilation and transport of nitrate and phosphate in intact plants of castor bean (Ricinus communis L.). Journal of Experimental Botany 48: 75-91. [ Links ]
13. Jeschke, W.D., A. Peuke, E.A. Kirkby, J.S. Pate & W. Hartung (1996). Effects of P deficiency on the uptake, flows and utilization of C, N and H2O within intact plants of Ricinus communis L. Journal of Experimental Botany 47: 1737-1754. [ Links ]
14. Kaiser, J.J. & O.A.H. Lewis (1984). Nitrate reductase and glutamine synthetase activity in leaves and roots of nitrate fed Helianthus annuus L. Plant and Soil 70: 127-130. [ Links ]
15. Kondracka, A. & A.M. Rychter (1997). The role of Pi recycling processes during photosynthesis in phosphate-deficient bean plants. Journal of Experimental Botany 48: 1461-1468. [ Links ]
16. Krom, M.D. (1980). Spectrophotometric determination of ammonia: a study of a modified berthelot reaction using salicylate and dichloroisocyanurate. Analyst 105:305-316. [ Links ]
17. Lam, H.M., K.T. Coschigano, I.C. Oliveira, R. Melo-Oliveira & G.M. Coruzzi (1996). The molecular genetics of nitrogen assimilation into amino acids in higher plants. Annual Review of Plant Physiology and Plant Molecular Biology 47: 569-593. [ Links ]
18. Lauer, M.J., D.G. Blevins & H. Sierzputowska-Gracz (1989). 31P-nuclear magnetic resonance determination of phosphate compartmentation in leaves of reproductive soybeans (Glycine max L.) as affected by phosphate nutrition. Plant Physiology 89: 1331-1336. [ Links ]
19. Lee, R.B. (1982). Selectivity and kinetics of ion uptake by barley plants following nutrient deficiency. Annals of Botany 50: 429-449. [ Links ]
20. Lillo, C. (1984). Diurnal varations of nitrite reductase, glutamine synthetase, glutamate synthase, alanina aminotransferase and aspartate in barley leaves. Physiologia Plantarum 61: 214-218. [ Links ]
21. López-Lefebre, L.R., J.M. Ruiz, R.M. Rivero, P.C. García, E. Sánchez & L. Romero (2000). Role of CaCl2 in ammonium assimilation in roots of tobacco plants (Nicotiana tabacum L.). Journal of Plant Physiology 156: 672-677. [ Links ]
22. Lynch, J., A. Läuchli & E. Epstein (1991). Vegetative growth of the common bean in response to phosphorus nutrition. Crop Science 31: 380-387. [ Links ]
23. Marschner, H. (1995). Mineral Nutrition of Higher Plants. Second Edition. Academic Press Inc., San Diego, CA. pp: 245-281. [ Links ]
24. Migge, A., G. Meya, E. Carrayol, B. Hirel & T.W. Becker (1996). Regulation of the subunit composition of tomato plastidic glutamine synthetase by light and the nitrogen source. Planta 200: 213-220. [ Links ]
25. Padget, P.E. & R.T. Leonard (1996). Free amino acid levels and the regulation of nitrate uptake in maize cell suspension cultures. Journal of Experimental Botany 47: 871-883. [ Links ]
26. Pilbeam, D.J., I. Cakmak, H. Marschner & E.A. Kirkby (1993). Effect of withdrawal of phosphorus on nitrate assimilation and PEP carboxylase activity in tomato. Plant and Soil 154:111-117. [ Links ]
27. Rabe, E. & C.J. Lovatt (1984). De novo arginine biosynthesis in leaves of phosphorus-deficient Citrus and Poncirus species. Plant Physiology 76: 747-752. [ Links ]
28. Raghothama, K. G. (1999). Phosphate acquisition. Annual Review of Plant Physiology and Plant Molecular Biology 50: 665-693. [ Links ]
29. Ress, T.A.V., T.R. Larson, J.W.C. Heldens & F.G.J. Huning (1995). In situ glutamine synthetase activity in a marine unicellular alga. Development of a sensitive colorimeric assay and the effects of nitrogen status on enzyme activity. Plant Physiology 109: 1405- 1410. [ Links ]
30. Rufty, T.W. Jr., D.W. Israel, J.R. Volk, J. Qui & S.A. Tongmin (1993). Phosphate regulation of nitrate assimilation in soybean. Journal of Experimental Botany 44: 879-891. [ Links ]
31. Rufty, T.W.Jr., C.T. Mackown & D.W. Israel (1990). Phosphorus stress effects on assimilation at nitrate. Plant Physiology 94: 328- 333. [ Links ]
32. Rufty, T.W.Jr., M.Y. Siddiqi, A.D. Glass & T.J. Ruth (1991). Altered 15NO3- influx in phosphorus limited plants. Plant Science 76: 43- 48. [ Links ]
33. Ruiz, J.M., N. Castilla & L. Romero (2000). Nitrogen metabolism in pepper plants applied with different bioregulators. Journal of Agricultural Food Chemistry 48: 2925-2929. [ Links ]
34. Ruiz, J.M. & L. Romero (1998). Calcium impact on phosphorus and its main bioindicators: response in the roots and leaves of tobacco. Journal of Plant Nutrition 21: 2273-2285. [ Links ]
35. Ruiz, J.M. & L. Romero (1999). Cucumber yield and nitrogen metabolism in response to nitrogen supply. Scientia Horticulturae 82: 309-316. [ Links ]
36. Sánchez, E., R.M. Rivero, J.M. Ruiz, & L. Romero (2004). Changes in biomass, enzymatic activity and protein concentration in roots and leaves of green bean plants (Phaseolus vulgaris L. cv. Strike) under high NH4NO3 application rates. Scientia Horticulturae 99: 237-248. [ Links ]
37. SAS (1987). SAS/STAT Guide for Personal Computers. In: pp: 1028- 1056. Version 6 Statistical Analysis System Institute, Inc; Cary, NC. [ Links ]
38. Schachtman, D.P., R.J. Reid & S.M. Ayling (1998). Phosphorus uptake by plants: from soil to cell. Plant Physiology 116: 447-453. [ Links ]
39. Singh, S.P. (1999). Production and utilization. In: Sing, S.P. (ed.), pp. 1-24. Common Bean Improvement in the Twenty-First Century. Developments in Plant Breeding. Vol. 7. Kluwer Acad., Dordrecht, The Netherlands. 210 p. [ Links ]
40. Singh, R.P. & H.S. Srivastava (1986). Increase in glutamate synthase activity in maize seedlings in response to nitrate and ammonium nitrogen. Physiologia Plantarum 66: 413-416. [ Links ]
41. Sivasankar, S. & A. Oaks (1996). Nitrate assimilation in higher plants: the effect of metabolites and light. Plant Physiology and Biochemistry 34: 609-620. [ Links ]
42. Slawyk, G. & M. Rodier (1988). Biosynthetically active glutamine synthetase in the marine diatom Phaeodactylum tricornutum: optimization of the forward-reaction assay. Marine Biology 97: 269-274. [ Links ]
43. Stitt, M. (1999). Nitrate regulation of metabolism and growth. Current Opinion in Plant Biology 2: 178-186. [ Links ]
44. Theodorou, M.E. & W.C. Plaxton (1993). Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant Physiology 101: 339-344. [ Links ]
45. Vance, C.P. & S. Stade (1984). Alfalfa root nodule carbon dioxide fixation. II. Partial purification and characterization of root nodule phosphoenolpyruvate carboxilate. Plant Physiology 75: 261- 264. [ Links ]
46. Wolf, B. (1982). A comprehensive systems of leaf analysis and its use for diagnosing crop nutrients status. Communications in Soil Science and Plant Analysis 13: 1035-1059. [ Links ]
47. Woodall, J., J.B. Boxall, B.G. Forde & J. Pearson (1996). Changing perspectives in plant nitrogen metabolism: The central role of glutamine synthetase. Science Progress 79: 1-26. [ Links ]
48. Yemm, E.W. & E.C. Cocking (1955). The determination of amino acids with ninhydrin. Analyst 80: 209-213. [ Links ]

