










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkPhyton (Buenos Aires)
On-line version ISSN 1851-5657
Phyton (B. Aires) vol.78 no.1 Vicente López Jan./June 2009
ARTÍCULOS ORIGINALES
Efecto del tamaño de celdas y citoquininas en el crecimiento de plantas de petunia
Effect of cell size and cytokinins on growth of petunia plants
Lagoutte1 S, M Divo de Sesar2, F Vilella3
1 Cátedra de Jardinería, Facultad de Agronomía, Universidad de Buenos Aires.
2 Cátedra de Producción Vegetal, Facultad de Agronomía, Universidad de Buenos Aires.
3 Cátedra y Programa de Agronegocios, Facultad de Agronomía, Universidad de Buenos Aires.
Address Correspondence to: M. Divo de Sesar, Av. San Martín 4453, Ciudad Autónoma de Buenos Aires. CP1417DSQ. e-mail: divomart@agro.uba.ar
Recibido-Received 14.08.2008.
Aceptado-Accepted 09.01.2009.
Resumen. Los cultivos de plantas florales anuales se realizan en bandejas alveoladas o contenedores. La variación en el tamaño de la celda de la bandeja de germinación altera el volumen donde se desarrollan las raíces, lo que afecta el crecimiento de la planta. Este trabajo evaluó el efecto del tamaño inicial del contenedor en el crecimiento de plántulas de Petunia x híbrida cv Dream Mix, suplementadas, luego del repique, con citoquininas. Plántulas obtenidas en bandejas de 90 y 288 alvéolos, se trasplantaron a macetas de 1L y fueron tratadas con dosis de 6-bencilaminopurina (BAP), resultando 4 tratamientos: 2 tamaños de contenedor por 2 BAP (con y sin). El tamaño de la celda condicionó el desarrollo de la parte aérea y radical, ya que en las de menor tamaño disminuyó (p<0,001) el ritmo de crecimiento y la relación tallo/raíz. Al final del ensayo, las diferencias en el número de ramificaciones en las plantas con BAP (independientemente del tamaño del alvéolo del cual provenían) fue altamente significativo (p<0,001) con respecto al testigo sin BAP (14,7 vs 6,9). El peso final y la tasa de crecimiento de las plantas en macetas, provenientes de bandejas de 288 celdas, fueron menores a las provenientes de bandejas de 90 celdas. El tamaño de las celdas utilizadas en los períodos tempranos de crecimiento, condicionó el tamaño y la calidad final de plantas de petunia. La suplementación con BAP permite lograr plantas de mayor tamaño con muy buena estructura de planta.
Palabras clave: Tamaño de la bandeja de germinación; Reguladores de crecimiento; Calidad de planta; BAP.
Abstract. Annual floral plant crops are produced in cells or pots of various sizes. Varying container size alters the volume which plant roots can explore, which in turn affect plant growth. This work assesed the effect of the initial container size on growth of Petunia x híbrida cv Dream Mix, supplemented with cytokinins after potting. Seedlings obtained in trays of 90 or 288 cells, were transplanted to 1l pots. They were then supplemented with 6-bencyladenine (BA). The assay consisted of 4 treatments: two container sizes by two BA conditions (with and without). Cell size conditioned growth above ground and below ground; growth rate and shoot/root ratios decreased (p<0.001) as cell size decreased. At the end of the assay, branching differences on plants supplemented with BA were highly significant (p<0.001; 14.7 versus 6.9) with respect to those without BA. This occurred independently of cell size the plant came from. Final weight and growth rate of potted plants, coming from trays of 288 cells, were smaller than those coming from trays of 90 cells. Cell size employed in early growth stages conditioned size and final quality of petunia plants. Supplementation with BA allows obtaining better developed, greater size plants.
Key words: Cell size; Growth regulators; Plant quality; BA.
INTRODUCCIÓN
Petunia es una especie que pertenece a la familia de las Solanáceas. Es muy utilizada como plantín de bordura y de las más populares entre las florales anuales. Posee un largo período de floración, observándose en los jardines durante la primavera y el verano. Son muy fáciles de cultivar y hay genotipos de diferentes formas y colores.
Si bien los cultivos de plantas florales anuales se realizan en contenedores con diferentes funciones y/o tamaño, tradicionalmente la producción de plantines de bordura se realizaba en bandejas de germinación. En la actualidad los productores adoptaron, ampliamente, la producción en bandejas alveoladas. El plantín obtenido de las mismas se conoce como plug. Un plug es una plántula producida en un pequeño volumen de sustrato contenido en un alvéolo o celda (Hartmann et al., 2002).
Cuando las plantas desarrollan sus raíces en un volumen de sustrato restringido, se observan cambios morfológicos y fisiológicos que afectan la calidad del plantín y su productividad. El crecimiento de vástagos y raíces, la acumulación de biomasa y su partición, la fotosíntesis y el contenido de clorofila foliar, las relaciones hídricas, la captación de nutrientes, la respiración y la floración son todos afectados por el tamaño del contenedor (NeSmith y Duval, 1998). Por lo tanto, el tamaño de las celdas condicionaría el desarrollo y el tiempo de permanencia en las bandejas, y el crecimiento del sistema radical. La partición de fotoasimilados entre diferentes partes de la planta podría ser distinto de acuerdo al grado de restricción que ofrece la celda (Xu y Kafkafi, 2001).
A diferencia de otras plantas ornamentales, los plantines de bordura deben realizar la mayor parte de su crecimiento y floración luego de la venta (Holbcomb, 1994). Ello se conecta en forma directa con la longevidad y la calidad de las mismas. Si bien no existe una definición exacta de planta de calidad, ya que la misma involucra la subjetividad del observador, se pueden señalar ciertos parámetros cuantitativos, además de los estéticos (cualitativos). Éstos, aunque no estén estandarizados, son perceptibles por el consumidor y se relacionan positivamente con un producto superior: número y tamaño de las ramificaciones, adecuada relación tallo/raíz, turgencia e intensidad del color en hojas y flores, forma y compacidad de la planta (entendiéndose como tal la relación armónica y específica entre altura y diámetro de planta) (Divo de Sesar, 2005). Carey et al. (2007) afirman que, desde el punto de vista de la producción, una planta de alta calidad y buena estructura es aquella que no está ahilada, posee buen número de ramificaciones y abundante floración, y es lo suficientemente compacta como para minimizar el deterioro cuando se embalan a altas densidades. Los reguladores de crecimiento, que inhiben la síntesis de las giberelinas, pueden controlar la altura pero no mejorar la ramificación (Carey et al., 2007).
Las citoquininas fueron descubiertas en el Laboratorio del Profesor Skoog, en la Universidad de Wisconsin, en 1954, y se definieron como una clase de sustancias que, en presencia de una concentración óptima de auxina, inducen la división celular en el tejido parenquimático de la médula del tallo de tabaco (Nicotiana tabacum L.) (Mok y Mok, 1994). Sin embargo, los efectos de las mismas no están limitados a la división celular. Estas hormonas han demostrado ser muy importantes en otras fases del desarrollo de las plantas. En ensayos anteriores se demostró que plantas tratadas con citoquininas poseían más ramificaciones, de mayor longitud y peso, y se observaba mayor concentración de clorofilas (Divo de Sesar et al., 2003).
El objetivo de este trabajo fue evaluar el efecto residual del tamaño de los alvéolos en el crecimiento inicial de las plántulas de Petunia x híbrida cv Dream Mix, y de la suplementación con citoquininas luego del repique a macetas.
MATERIALES Y MÉTODOS
Los ensayos se realizaron en el Laboratorio e Invernáculo de la Cátedra de Producción Vegetal, Facultad de Agronomía, Universidad de Buenos Aires (34,35° S y 58,29° O).
Etapa en bandeja: En Enero de 2006, semillas de Petunia x híbrida cv Dreams, F1, serie Mix de Ball, se sembraron al voleo, en cajas de PVC con tapa, en un sustrato compuesto por perlita y turba (6:2; v/v). Las mismas se incubaron en Cámara de Crecimiento, a 23 ± 2 °C, con régimen lumínico de 16 horas de luz y 8 de oscuridad, logrados con tubos de luz blanca fría, con 57 μmol/m2/s. A los 15 días desde la siembra, se realizó el repique a bandejas alveoladas con 90 ó 288 celdas (270 plántulas para cada tamaño de alvéolo) conteniendo un sustrato comercial (Dinamics 1®), a base de perlita, turba, vermiculita y corteza de pino. El mismo se caracterizó por poseer un pH de 5,85, una conductividad eléctrica de 175 μS/cm y 63 ppm de sólidos disueltos. Las plántulas se distribuyeron al azar. Las bandejas se mantuvieron en invernáculo, bajo media sombra (saram 30%) y riego intermitente; la frecuencia y tiempo de mojado se programaron según la demanda ambiental.
En el momento del repique, y a los 10 y 20 días del mismo (1ª, 2ª y 3er fecha, respectivamente) se realizaron muestreos totalmente al azar (7 plántulas de cada tratamiento para cada momento de muestreo). Las variables cuantificadas fueron peso fresco y seco de la parte aérea y de las raíces (mg), calculándose las relaciones entre el peso fresco ó el peso seco de la parte aérea y las raíces (relación Pa/R) y la tasa de crecimiento del cultivo (TCC) (mg/día). Este último consistió en el cociente entre la diferencia de pesos (mg) de 2 momentos de muestreo consecutivos y el tiempo entre los mismos (días).
Etapa en maceta: A las 5 semanas desde la siembra, los plantines se trasplantaron a macetas de 1 l, con un sustrato compuesto por tierra, resaca, perlita, vermiculita y turba rubia (5:2:2:0,5:0,5; v/v). El pH inicial de la mezcla fue 5, la conductividad eléctrica 25 μS/cm y los sólidos disueltos 11 ppm. La fertilización se realizó en forma quincenal con 2 g/l de sustrato de Nitrofoska® Elite, de Compo®, (12:10:20; con S, Ca, Mg y micronutrientes). Las macetas se mantuvieron en las mismas condiciones ambientales que las bandejas; el control de malezas y el riego se realizaron en forma manual. La mitad de las plantas, provenientes de cada tamaño de celda, fue suplementada 3 veces, quincenalmente, con 6-bencilaminopurina (BAP) (5mg/l, 5 ml/ pl) y la otra mitad se utilizó como tratamiento testigo.
Las variables cuantificadas fueron: número de ramificaciones, número de flores finales, peso fresco y seco de la parte aérea y de las raíces (g), calculándose las relaciones entre el peso fresco ó el peso seco de la parte aérea y las raíces (relación Pa/R), la tasa de crecimiento del cultivo (TCC) (mg/día) y el perfil de clorofilas. Al igual que en la etapa anterior; la TCC fue el resultado del cociente entre la diferencia de los pesos de plantas (mg) finales (a los 98 días) e iniciales (momento del trasplante) (mg) para la etapa en maceta y la duración de la misma (días) (63 días).
El perfil de clorofilas se evaluó de la siguiente forma: durante las últimas 4 semanas de cultivo se realizaron tres mediciones en distintos momentos (en la 5ª, 7ª y 9ª semana desde el trasplante a maceta, coincidiendo la última medición con la finalización del ensayo). En cada momento se midieron 8 ejemplares de cada tratamiento (seleccionados aleatoriamente), se tomaron 5 muestras de hojas totalmente expandidas de distintas partes de la planta, y se realizaron lecturas manuales de cada hoja (3 lecturas por hoja) con un Minolta SPAD 502 Meter.
Al finalizar el ensayo se tomaron al azar 7 plantas de cada tratamiento y se midieron la altura y el diámetro mayor y menor, promediándose la sumatoria de los 3 valores. A cada valor obtenido, se le restó el coeficiente de dispersión (σn-1) correspondiente, y se le sumó el número de ramificaciones; el promedio de estos últimos 3 valores definió un Índice de Calidad de Planta (ICP). Mayores valores para el número de ramificaciones y el promedio de los diámetros conducen a un mayor ICP. El σn-1 se relaciona en forma directa (negativamente) con la uniformidad de cada planta, es decir que un menor valor del σn-1 implica una mayor uniformidad de planta, por lo tanto, corresponde restarlo. Un valor absoluto de ICP mayor indica que la planta es de mayor calidad.
Para pesar se utilizó una balanza analítica Mettler H80 (Precisión 0,1 mg). El material se secó en estufa a 75°C hasta peso constante. Los datos se analizaron de acuerdo a un modelo de análisis de varianza para dos factores, con un diseño completamente aleatorio. Los factores fueron: Tamaño de alvéolo x Momento de muestreo, y Tamaño de alvéolo x Tratamiento hormonal para las etapas en bandeja y en maceta, respectivamente. Los promedios se compararon según el Test de Tukey. Se utilizó el software estadístico InfoStat 2002 (Di Rienzo et al., 2002). El ensayo se finalizó a las 14 semanas desde la siembra.
RESULTADOS Y DISCUSIÓN
Etapa en bandeja: En las Tablas 1 y 2 se describe el crecimiento a través del tiempo de las plántulas de petunia y la TCC. Desde el segundo muestreo, el tamaño de la celda redujo (p<0,001) el crecimiento y desarrollo de la parte aérea y radical; las plántulas provenientes de celdas más grandes crecieron más que las de celdas más pequeñas. Esta tendencia fue similar tanto para el peso fresco (Tabla 1) como para el peso seco (Tabla 2). La síntesis y el balance de carbono estarían limitados por el volumen de la celda que, en el caso de las bandejas de 288 celdas, aportaría menor cantidad de agua y nutrientes a la plántula (Hartmann et al., 2002), condicionando el tamaño de la parte aérea.
Tabla 1. Peso Fresco (mg) de la parte aérea y las raíces de plántulas de petunia, provenientes de bandejas con distinto volumen de alvéolos y diferentes momentos de muestreo (a los 15, 25 y 35 días desde la siembra, correspondientes a la 1ª, 2ª y 3ª fecha respectivamente), y Tasa de crecimiento del cultivo (TCC) (mg/día) entre la siembra y los respectivos momentos de muestreo.
Table 1. Shoot and root Fresh weight (mg) of petunia seedlings, coming from different cell size trays and different sampling times (at 15, 25 and 35 days after seeding, corresponding to the 1st, 2nd, and 3rd dates, respectively), and crop Growth Rate (TCC) (mg/day) between seeding and the subsequent sampling times.

Tabla 2. Peso seco (mg) de la parte aérea y las raíces de plántulas de petunia, provenientes de bandejas con distinto volumen de alvéolos y diferentes momentos de muestreo (a los 15, 25 y 35 días desde la siembra, correspondientes a la 1ª, 2ª y 3ª fecha, respectivamente), y Tasa de crecimiento del cultivo (TCC) (mg/día) entre la siembra y los respectivos momentos de muestreo.
Table 2. Shoot and root Dry weight (mg) of petunia seedlings, coming from different cell size trays and different sampling times (at 15, 25 and 35 days after seeding, corresponding to the 1st, 2nd, and 3rd dates, respectively), and crop Growth Rate (TCC) (mg/day) between seeding and the subsequent sampling times.
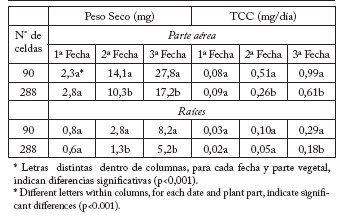
Si bien no siempre las diferencias fueron significativas, la partición a raíces disminuyó entre la primera y segunda medición para luego aumentar (Tabla 3). Asimismo, ésta siguió la misma tendencia que el crecimiento (Tablas 1 y 2). El desarrollo de la parte aérea y la partición a raíces fueron elevadas en las celdas más pequeñas en la primera fecha de medición. Sin embargo, esto se revirtió rápidamente por las limitaciones que ofreció el tamaño de las mismas, ya que las tasas de crecimiento de vástagos y raíces son interdependientes. El desarrollo de las raíces depende de la parte aérea de la planta para el suministro de fotoasimilados y diferentes reguladores de crecimiento, mientras que la parte aérea depende de las raíces para el anclaje y la disponibilidad de agua, nutrientes y hormonas (Tonutti y Giulivo, 1990). Cuando el sistema radical está restringido a un pequeño volumen, el balance entre vástagos y raíces se afecta volviéndose inestable (NeSmith y Duval, 1998). Se ha demostrado que la restricción en el crecimiento de las raíces reduce el peso seco total (follaje y raíces) (Wien, 1997; Mugnai et al., 2000). Vavrina (2002) comprobó que incluso 45 días después del trasplante, plantas de tomate en invernadero provenientes de contenedores con alvéolos grandes, presentaron un crecimiento notablemente mayor que plantas obtenidas en alvéolos pequeños. Los plantines con un sistema radical relativamente grande, generalmente sufren menos el estrés al trasplante y, de esta forma, el crecimiento posterior de los mismos se anticipa al de los que poseen raíces de menor tamaño (Weston y Zandstra, 1986).
Tabla 3. Relaciones tallo/raíz en plántulas de Petunia, bandejas con distinto volumen de alvéolos, para diferentes momentos de muestreo (a los 15, 25 y 35 días desde la siembra, correspondientes a la 1ª, 2ª y 3ª fechas, respectivamente).
Table 3. Shoot/root ratios in Petunia seedlings, different cell size trays, at different sampling times (at 15, 25 and 35 days after seeding, corresponding to the 1st, 2nd, and 3rd dates, respectively).

Si bien las bandejas con alvéolos de menor volumen permiten la utilización más eficiente del espacio y tiempo de cultivo, y los plantines provenientes de los mismos son más fáciles de manipular (Holcomb, 1994), es evidente que plantas de calidad sólo se logran con celdas de mayor tamaño.
Etapa en maceta: Las diferencias en el perfil de clorofilas sólo fueron significativas (p<0,001) en las plantas tratadas con citoquininas en una medición temprana, 15 días después de iniciada dicha etapa (Tabla 4). Se ha mencionado que existirían dos posibles efectos del BAP sobre la RuBP carboxilasa en hojas: uno directo, al inducir la síntesis de la enzima, y otro indirecto al mejorar la efectiva partición de N a hojas (Ookawa et al., 2004). Se podría postular entonces que, al no haberse realizado una fertilización diferencial en las plantas suplementadas con BAP (que crecieron más), hubo un efecto de dilución del N en la hoja. Por otro lado, se ha demostrado, en Vitis labrusca, que el contenido de N en hoja se eleva a medida que la oferta de N aumenta, observándose una relación lineal positiva entre las concentraciones de clorofila, N en hoja y el balance de carbono (Chen y Cheng, 2003). Bryant et al. (1987) demostraron que el valor nutricional de las hojas de Populus tremuloides Michx aumentaba con el incremento de la oferta de N del suelo. Un par de años después, Evans (1989) afirmó que el N de los tilacoides es positivamente proporcional a la concentración de clorofilas. Este autor informó que existe una fuerte correlación lineal positiva entre las concentraciones de clorofila o de RuBP carboxilasa y el N en varias especies vegetales (trigo, espinaca, algodón, poroto, Chenopodium album, Alocasia sp).
Tabla 4. Perfil de clorofilas de plantas de petunia, con y sin suplementación con BAP, evaluadas en diferentes momentos de muestreo (5 y 19 de Abril, y 3 de Mayo, correspondientes a la 5ª, 7ª y 9ª semana desde el trasplante, respectivamente).
Table 4. Chlorophyll profiles of petunia plants, either supplemented or not with cytokinins, at different sampling times (5 and 19 April, and 3 May, corresponding to the 5th, 7th and 9th week from transplanting, respectively).

Las condiciones restrictivas que se obtienen con alvéolos más pequeños tienen un efecto residual sobre todo el ciclo del cultivo, reduciendo (p<0,001) la tasa de crecimiento aún en las plantas tratadas con BAP (Tabla 5), condicionando el tamaño y/o la calidad final de la planta (Tablas 7 y 8). Al analizar la relación tallo/ raíz se observa el efecto del BAP, que favorece (p<0,001) el crecimiento de la parte aérea (Tabla 6), según lo sugerido por Davies (1995). El crecimiento de las raíces con BAP, sin embargo, es mayor (p<0,001) en las plantas provenientes de celdas más grandes. No ocurre lo mismo en las plantas sin tratar en las que la relación es similar, si bien los pesos de la parte aérea son bien diferentes. La relación tallo/ raíz comienza a establecerse en los momentos posteriores a la germinación, cuando el crecimiento de las raíces es máximo. Cuando se trasplantan plántulas en las cuales el crecimiento de las raíces estuvo limitado, no suele compensarse la evapotranspiración de los días iniciales, aún cuando estén bien regadas (Aloni et al., 1991). Por lo tanto, esta restricción puede simular el efecto del estrés hídrico aún con suficiente humedad en el sustrato para el normal crecimiento de la planta (Krizek et al., 1985). Muchas respuestas morfológicas y/o fisiológicas de las plantas han sido investigadas respecto al volumen de los contenedores y condiciones limitadas para el crecimiento de las raíces (Brown, 1995). Sin embargo, lo que más interesa a los productores es el desempeño post-transplante de los plantines, minimizando el tiempo de recría posterior (Hall, 1989). Puede ser interesante establecer tiempos definidos de permanencia en bandejas alveoladas para cada tamaño de celda.
Tabla 5. Peso fresco final (g) y Tasa de Crecimiento (TCC) [mg/día; entre el momento del trasplante (5ª semana) y final del ensayo (14ª semana)] de la Parte aérea y las Raíces, con y sin BAP en plántulas de petunia provenientes de celdas de diferente volumen.
Table 5. Final fresh weight (g) and Growth Rate (GR) [mg/day; between transplanting time (5th week) and the study end (14th week) of shoots and roots, with and without BA, in petunia plants grown in different cell sizes.
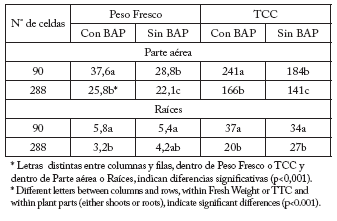
Tabla 6. Relación final Tallo/raíz, en plantas de petunia, con y sin BAP, provenientes de celdas de diferente tamaño.
Table 6. Final Shoot/root ratios in petunia plants, with and without BA, coming from different cell sizes.

Tabla 7. Número de ramificaciones, media de los promedios entre la altura y diámetros mayor y menor de cada planta [media Ø(cm)], y coeficientes de dispersión (σ n-1) de cada planta de petunia, con y sin BAP, provenientes de celdas de diferente volumen.
Table 7. Number of shoots, average of the means between both plant height and smallest and greatest diameters, and dispersion coefficients (σ n-1) of petunia plants, with and without BA, coming from different cell sizes.
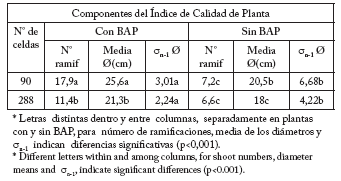
Tabla 8. Índice de Calidad de Planta (ICP) de plantas de petunia, con y sin BAP, provenientes de celdas de diferente volumen.
Table 8. Plant Quality Index (ICP) of petunia plants with and without BA, coming from different cell sizes.

Asimismo, las plantas tratadas con BAP, estaban listas para su comercialización a las 12 semanas desde la siembra, mientras que el resto no: el ensayo se finalizó a las 14 semanas, cuando el 30% de las plantas sin tratar había alcanzado la floración. Si bien en las diferentes especies ensayadas por este grupo de trabajo, siempre la suplementación con BAP retrasa levemente la floración, luego ésta se activa e incrementa y, como consecuencia, se obtienen plantas más grandes con mayor número de flores. En el momento de finalización del ensayo el número de flores abiertas en las plantas tratadas era significativamente mayor (p<0,001) (6,7 vs 3,3). Además, las diferencias en el número de ramificaciones existentes (Tabla 7) indicarían un mayor número de flores potenciales.
A lo largo de todo el ensayo las plantas de petunia en macetas y tratadas con BAP mostraron ser superiores a las no tratadas, al igual que otras especies ensayadas (Pelicano et al., 2007). Por lo tanto, se diseñó un ICP que incluyera en la cuantificación otros atributos o componentes de calidad, además de las variables evaluadas. El número de ramificaciones durante el primer mes desde el trasplante se mantuvo constante para todos los tratamientos (media: 6,5). Al final del ensayo las diferencias en el número de ramificaciones en las plantas tratadas con BAP, provenientes de celdas de mayor volumen fueron altamente significativas (p<0,001) con respecto a las provenientes de menor volumen y a los testigos sin BAP. Similar tendencia se observó en los σn-1. Esta tendencia se relaciona directamente con una planta con mayor uniformidad y, por lo tanto, mejor arquitectura. Divo de Sesar (2005) observó menor heterogeneidad en las diferentes variables evaluadas e interacción entre auxinas, fertilizantes y BAP en estacas enraizadas de olivo, arándano y jazmín amarillo. Sin embargo, en este trabajo con plantas de petunia no se encontraron interacciones entre los factores analizados.
Como ya se mencionó, este tipo de regulador de crecimiento ha demostrado ser muy importante en varias fases del desarrollo de las plantas, incluyendo la desinhibición y diferenciación de yemas, alargamiento celular, y flujo de asimilados y nutrientes a través de la planta. Las relaciones entre citoquininas, giberelinas y ácido abscísico controlan la dormición en semillas y tienen influencia en un gran número de procesos fisiológicos relacionados con el crecimiento y desarrollo de las plantas (Hartmann et al., 2002). Por lo tanto, las mismas podrían ser empleadas, con diferentes objetivos, en distintas etapas del proceso productivo de los cultivos ornamentales.
CONCLUSIONES
En este trabajo se demuestra que el tamaño de celda, utilizada en los períodos tempranos de crecimiento, condiciona el tamaño y la calidad final de plantas de petunia.
Asimismo, la suplementación con BAP permite lograr plantas de mayor tamaño con muy buena estructura de planta.
Reconocimiento: Este trabajo ha sido realizado con fondos provenientes del Subsidio UBACyT G025.
REFERENCIAS
1. Aloni, B., L. Daie y L. Karni (1991). Water relations, photosynthesis and assimilate partitioning in leaves of pepper (Capsicum annuum) transplants: Effect of water stress after transplanting. Journal of Horticultural Science 66: 75-80. [ Links ]
2. Brown, R.W. (1995). The water relations of range plants: Adaptations to water deficits. p. 291-413. En: Bedunah, D.J. y Sosebee R.E. (eds). Wildland Plants: Physiological Ecology and Developmental Morphology. Society for Range Management, Denver, CO. 710 p. [ Links ]
3. Bryant, J., J. Clausen, P. Reichardt, M. McCarthy y R. Werner (1987). Effect of nitrogen fertilization upon the secondary chemistry and nutritional value of quaking aspen (Populus tremuloides Michx.) leaves for the large aspen tortrix [(Choristoneura conflictana (Walker)]. Oecología 73: 513-517. [ Links ]
4. Carey, D., B. Whipker, I. McCall y W. Buhler (2007). Cytokinin based PGR affects growth of vegetative petunia SNA Research Conference, Vol. 52 Growth Regulators Section 102. 101-109 pp. [ Links ]
5. Chen, L. y L. Cheng (2003). Carbon assimilation and carbohydrate metabolism of Concord grape (Vitis labrusca L.) leaves in response to Nitrogen supply. Journal of American Society of Horticultural Science 128: 754-760. [ Links ]
6. Davies, P.J. (1995). The Plant Hormones: Physiology, Biochemistry and Molecular Biology. 2nd Ed, Kluwer. Academic Publishers, New York, 545 p. [ Links ]
7. Di Rienzo, J., A. Guzmán y F. Casanoves (2002). A multiple comparisons method based on the distribution of the root node distance of a binary tree. Journal of Agricultural, Biological and Environment Statistics 7: 1-14. [ Links ]
8. Divo de Sesar, M. (2005). Integración de estudios fisiológicos, histológicos y bioquímicos realizados durante el enraizamiento, rustificación y crecimiento posterior de especies de interés agronómico suplementadas con citoquininas. Tesis Doctoral. 222 pp. [ Links ]
9. Divo de Sesar, M., A. D'Ambrogio, J. Boquete, F. Vilella y A. Stella (2003). Determinación del momento óptimo de aplicación de BAP en estacas semileñosas de Jazminum mensyi. Floricultura en Argentina. Facultad de Agronomía, UBA. p. 27-36. [ Links ]
10. Evans, J. (1989). Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecología 78: 1-19. [ Links ]
11. Hall, M. (1989). Cell size of seedlings containers influences early vine growth and yield of transplanted watermelon. HortScience 24: 771-773. [ Links ]
12. Hartmann, H.T., D. Kester, F.T. Davies Jr. y R.L. Geneve (2002). Plant Propagation: Principles and Practices. Seventh Edition, Prentice Hall Inc., New Jersey. 880 p. [ Links ]
13. Holcomb, E. (1994). Bedding plant . 4th Edition, Ball Publisihing, USA . 430 p. [ Links ]
14. Krizek, D., A. Carmi, F. Mirecki, F. Snayder y J. Bruce (1985). Comparative effects of soil moisture stress and restricted root zone volume on morphogenetic and physiological responses of soybean (Glicine max (L.) Merr). Journal of Experimental Botany 36: 25-38. [ Links ]
15. Mok, D.W. y M.C. Mok (1994). Cytokinins, Chemistry, Activity and Function. CRC Press. Boca Raton. 339 p. [ Links ]
16. Mugnai, S., P. Vernieri y F. Tognoni (2000). Container volume effects on morphology and physiology of tomato seedlings. Acta Horticulturae 516: 499-505. [ Links ]
17. NeSmith, D. y J. Duval (1998). The effect of container size. Hort. Technology 8: 4. [ Links ]
18. Ookawa, T., Y. Naruoka, A. Sayama y J. Hirasawa (2004). Cytokinin Effects on Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase and Nitrogen Partitioning in Rice during Ripening. Crop Science 44: 2107-2115. [ Links ]
19. Pelicano, A., M. Divo De Sesar, N. Zamuner, J. Danelon y M. Yoshida (2007). Consecuencia de la calidad del alimento sobre la productividad del gusano de seda (Bombyx mori). Revista Ciencia e Investigación Agraria 34: 81-89. [ Links ]
20. Tonutti, P. y C. Giulivo (1990). Effect of available soil volume on growth of young kiwi plants. ActaHorticulturae 282: 283-294. [ Links ]
21. Vavrina, C.S. (2002). An introduction to the production of containerized, vegetable transplants. Horticultural Sciences Department, University of Florida, Institute of Food and Agricultural Sciences (IFAS). Fact Sheet HS849. [ Links ]
22. Weston, L. y B. Zandstra (1986). Effect of roots container size and location of production on growth and yield of tomato transplants. Journal of American Society of Horticultural Science 111: 498-501. [ Links ]
23. Wien, H.C. (1997). Transplanting. En: Wien, H.C. The Physiology of Vegetable Crops. Reprinted 2002. Cabi Publishing. 662 p. [ Links ]
24. Xu, G. y U. Kafkafi (2001). Nutrient supply and container size effects on flowering, fruiting assimilate allocation and water relations on sweet pepper. Acta Horticulturae 554: 113-120. [ Links ]

