










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.78 no.2 Vicente López jul./dic. 2009
ARTÍCULOS ORIGINALES
Control biológico de la marchitez del chile (Capsicum annuum L.) por Bacillus thuringiensis
Biological control of chili pepper root rot (Capsicum annuum L.) by Bacillus thuringiensis
Mojica-Marín1 V, HA Luna-Olvera2, CF Sandoval-Coronado2, B Pereyra-Alférez2, LH Morales-Ramos2, NA González-Aguilar2, CE Hernández-Luna2, OG Alvarado-Gomez3
1 Maestro-Investigador de La Facultad de Ciencias Químicas. UJED. Av. Veterinaria s/n, Circuito Universitario Durango, Dgo. México, CP. 34120.
Address Correspondence to: Virgilio Mojica Marín, Maestro-Investigador de La Facultad de Ciencias Químicas. UJED. Av. Veterinaria s/n, Circuito Universitario Durango, Dgo. México, CP. 34120 Tel. Y Fax: (618) 1301111. e-mail: vmojica@citologica.com; vmojicamx@yahoo.com.mx
2 Maestro-Investigador del Instituto de Biotecnología, Facultad de Ciencias Biológicas, UANL. Apdo. Postal 38F, Pedro de Alba, s/n, Cd. Universitaria San Nicolás de los
Garza, N.L. México, CP. 66450.
3 Maestro-Investigador de la Facultad de Agronomía. UANL. Carretera Zuazua -Marín Km 17.5, Marín, N.L. México, CP. 66700.
Recibido-Received 24.01.2008.
Aceptado-Accepted 2.06.2009.
Resumen. La expansión de la marchitez del chile en nuevas plantaciones y diferentes zonas productoras de México, y la dificultad en el control de la misma, obligan a desarrollar nuevas medidas de control. Se espera que las mismas auspicien mejores expectativas para el control de la enfermedad en el contexto de una estrategia de manejo integrado. El presente estudio se realizó con el objetivo de evaluar y determinar la capacidad antagónica de 64 cepas de Bacillus thuringiensis sobre Rhizoctonia solani, Phytophthora capsici y Fusarium oxysporum mediante estudios de cultivos duales en placas Petri. Las cepas de Bacillus thuringiensis provinieron de la Colección Internacional de Bacilos Entomopatógenos de la FCB-UANL. Fueron 16 cepas de B. thuringiensis que redujeron significativamente el crecimiento radial micelial de R. solani, 19 para P. capsici y ocho para F. oxysporum, después de tres días de incubación a 25°C. Por otra parte, las mejores cepas bacterianas se evaluaron en semillas de chile variedad Anaheim mediante pruebas de germinación bajo condiciones in vivo: se observó un incremento en el porcentaje de germinación en el caso del tratamiento de inoculación con B. thuringiensis sobre R. solani.
Palabras clave: Bacillus; Control biológico; Antagonismo; Germinación; Fitopatógenos con origen en el suelo.
Abstract. Wide spreading of pepper blight on new plantations and on different production areas in Mexico, and the difficulty in controlling this disease, makes it compulsory to develop new control measures. It is expected that these measures provide new tools for controlling this disease in the context of integrated management strategies. The aim of this investigation was to evaluate and determine the antagonistic potential of 64 strains of Bacillus thuringiensis against Rhizoctonia solani, Phytophthora capsici and Fusarium oxysporum by dual culture assays. Strains of B. thuringiensis came from the International Collection of Entomopatogenic Bacillus from FCBUANL. Results showed that 16 strains of B. thuringiensis reduced significantly the radial mycelial growth of R. solani, while 19 strains did the same for P. capsici and 8 strains for F. oxysporum after three days of incubation at 25°C. In addition, the best bacterial strains were evaluated on Anaheim variety pepper seeds by in vivo germination trials: results showed an increase on germination percentage in the case of seed inoculation with B. thuringiensis over R. solani.
Key words: Bacillus; Biological control; Antagonism; Germination; Soil-borne plant pathogens.
INTRODUCCIÓN
En México, el chile (Capsicum annuum L.) es el cultivo hortícola más importante considerando la superficie que se siembra (FOASTAT, 2005). En la actualidad, entre los factores más importantes que limitan su producción se encuentran los fitopatógenos provenientes del suelo (Velásquez et al., 2001). Entre ellos se encuentran los géneros Pythium, Rhizoctonia, Fusarium y Phytophthora spp. Éstos causan una enfermedad conocida comúnmente con el nombre de "tristeza o marchitez del chile". Su presencia se ha reportado en todos los estados productores de chile en México (Redondo, 1974; Guigon y González, 2001). Las pérdidas estimadas a nivel nacional oscilan entre un 10 y un 60%, mientras que en áreas especificas del Bajío y Puebla pueden alcanzar el 100% (Pérez et al., 2003). Se han utilizado diversos métodos para controlar esta enfermedad, siendo las prácticas culturales (Ristaino, 1991) y el control químico los más empleados (Parra y Ristaino, 2001). Sin embargo, el control biológico ha tomado una gran relevancia en los últimos años.
El género Bacillus, independientemente de su ubicación taxonómica, se encuentra entre los agentes más adecuados para el control biológico debido a cualidades tanto morfológicas como fisiológicas que permiten su ubicuidad en la naturaleza (Martin y Travers, 1989; Smith y Couche, 1991; Meadows et al., 1992). Este género además ha tenido mucho éxito en la prevención de patologías vegetales causadas por hongos (Morgan, 1963; Reedy y Rahe, 1989; Rytter et al., 1989; Turner y Backman, 1991; Smith et al., 1993; He et al., 1994; Osburn et al., 1995). El B. thuringiensis es el organismo más exitoso en cuanto a la comercialización (Valadares-Inglis et al., 1998). Éste produce un cuerpo paraesporal, denominado δ-endotoxina [formada por una o varias proteínas cristal insecticida (PsCI)], que es tóxico a lepidópteros, dípteros, coleópteros, nemátodos y protozoarios (Vankova, 1957; Smith y Couche, 1991). Sin embargo, su potencial antifúngico es desconocido, a pesar de su especificidad, virulencia, seguridad y potencia contra organismos patógenos (Pendleton, 1968; De Barjac y Lajudie, 1974; Rowe y Margaritis, 1987; Gelernter y Scwab, 1992). Además, produce una gran diversidad de metabolitos, entre los que destacan: bacteriocinas, antibióticos y enzimas extracelulares (como proteasas y quitinasas), elementos clave en el fenómeno de supresión de patógenos por agentes biológicos.
El objetivo de este trabajo fue seleccionar cepas de B. thuringiensis con potencial antagonista para controlar R. solani, P. capsici y F. oxisporum en condiciones in vitro e in vivo. Es decir, se propuso obtener una alternativa biológica para el control de la marchitez del cultivo del chile.
MATERIALES Y MÉTODOS
Selección de cepas de B. thuringiensis. Sesenta y cuatro cepas de B. thuringiensis se obtuvieron de la Colección Internacional de Bacilos Entomopatógenos de la FCB-UANL. Éstas permanecieron liofilizadas hasta que se activaron para su uso en tubos de ensayo conteniendo 2,5 ml de caldo nutritivo a pH 7. Éstos tubos se incubaron a una temperatura de 30°C durante 24 h en agitación rotatoria a 150 rpm. Posteriormente, las cepas fueron resembradas en agar nutritivo inclinado a pH 7, y mantenidas en incubación a 28°C durante 48 h.
Aislamiento de los hongos fitopatógenos. Rhizoctonia solani, P. capsici y F. oxysporum fueron obtenidos a partir de raíces de plantas de chile con síntomas típicos de la enfermedad, y conservados en cultivos puros en glicerol.
Determinación de antagonismo in vitro. Un disco o plug de 10 mm de cada hongo fue colocado en el centro de una caja Petri con papa dextrosa agar (PDA). Posteriormente, se trazó un círculo de 6 cm de diámetro rodeando el inóculo fúngico, que recibió una suspensión de B. thuringiensis conteniendo 5x109 ufc/mL. Las cajas fueron incubadas durante 72 horas a 22±2°C, y se evaluó el crecimiento radial, tasa de crecimiento y el porcentaje de inhibición; la información obtenida se comparó con el control, donde la suspensión bacteriana fue sustituida por agua destilada estéril. Los resultados se expresan como el porcentaje de inhibición correspondiente a cada una de las cepas bacterianas. El porcentaje de inhibición fue calculado usando la fórmula siguiente: % de inhibición = [1 - (crecimiento fúngico en agar que recibió la suspensión bacteriana/ crecimiento fúngico en el control) x 100].
Efecto de B. thuringiensis sobre la germinación de semillas de chile. Se emplearon semillas de chile de la variedad Anaheim. Éstas fueron desinfectadas en una solución de hipoclorito de sodio al 2% durante cinco minutos, seguida de tres lavados de tres minutos en agua destilada estéril. Las semillas se sumergieron durante una hora en suspensiones individuales de cada una de las cepas de B. thuringiensis. Las suspensiones contuvieron agua sacarosa al 3%, y 1 x 108 ufc/mL. Las semillas fueron posteriormente sembradas en recipientes plásticos conteniendo 300 g de una mezcla estéril de suelo, peat-moss, vermiculita y perlita (1:1:1). Los recipientes habían sido previamente inoculados con los fitopatógenos y mantenidos bajo condiciones de laboratorio a una temperatura de 22-25°C. Para el inóculo de R. solani, la suspensión se ajustó hasta alcanzar 106-7 propágulos o fragmentos por ml, para P. capsici 105-6 zoosporas por ml, y para F. oxisporum 106-7 conidios por ml. Se realizaron dos controles en donde las semillas se trataron solo con la solución estéril de agua destilada-sacarosa al 3%, y se sembraron con y sin inóculo del fitopatógeno. Cada tratamiento consistió de cuatro repeticiones, y cada una contuvo cuatro plantas. La eficacia de las cepas de B. thuringiensis fue calculada en base al porcentaje de germinación a los 20 días posteriores a la siembra.
Diseño del experimento. En cada experimento se utilizó un Diseño Completamente al Azar con Arreglo Factorial Ax- BxC considerando cada fitopatógeno por triplicado, y repetido por lo menos tres veces. Para la comparación de medias se utilizó la prueba de Tukey (p<0,05). Se utilizó la versión 6.12 de SAS (SAS Institute, Cary, NC, USA).
RESULTADOS
Determinación de antagonismo in vitro. Entre las 64 cepas de B. thuringiensis probadas (Tabla 1), 43 resultaron ser buenas antagonistas contra R. solani, P. capsici y F. oxysporum en ensayos de cultivos duales. En éstos la inhibición fue claramente observada por un menor crecimiento o la ausencia completa del micelio fúngico en la zona de inhibición que rodeaba la cepa bacteriana.
Tabla 1. Cepas de B. thuringiensis usadas en este estudio.
Table 1. Bacillus thuringiensis strains used in this study.

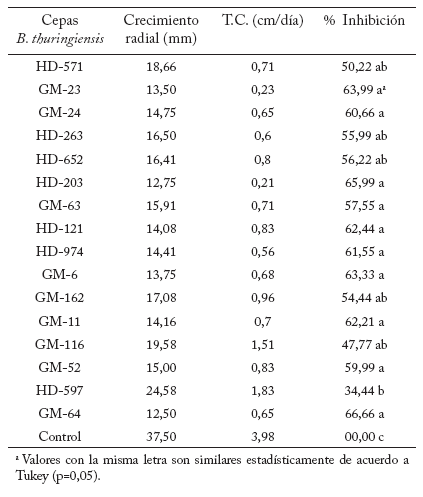
Dieciséis cepas de B. thuringiensis tuvieron efectos antagonistas en el crecimiento micelial de R. solani (Tabla 2). Las cepas GM-64 (66,66%) y HD-203 (65,99%) produjeron la mayor inhibición de crecimiento (p=0,05), mientras que la cepa HD-597 presentó el menor porcentaje de inhibición (34,4%). También se observó una reducción en la tasa de crecimiento micelial de 3,98 a 0,6 cm/día.
Tabla 2. Efecto antagonista de cepas de B. thuringiensis en el crecimiento radial, tasa de crecimiento y porcentaje de Inhibición micelial de R. solani.
Table 2. Antagonistic effect of B. thuringiensis strains in radial growth, growth rate and percentage mycelial growth inhibition of R. solani.
Para P. capsici, 19 cepas de B. thuringiensis se comportaron de una manera muy similar, produciendo porcentajes de inhibición del 15,14 al 27,80% (para las cepas HD-193 y GM- 66, respectivamente), y una reducción en la tasa de crecimiento de entre 2,86 a 1,26 cm/día (Tabla 3).
Tabla 3. Efecto antagonista de cepas de B. thuringiensis en el crecimiento radial, tasa de crecimiento y porcentaje de inhibición micelial de P. capsici.
Table 3. Antagonistic effect of B. thuringiensis strains in radial growth, growth rate and percentage mycelial growth inhibition of P. capsici.

Sólo ocho cepas mostraron un efecto inhibitorio (p=0,05) contra F. oxisporum (Tabla 4). De éstas sólo cinco inhibieron más del 30% del crecimiento, alcanzándose la máxima inhibición con las cepas GM-23 (43,02%) y HD-121 (42,04%), y una reducción en la tasa de crecimiento de 0,52 a 0,10 cm/ día. Al realizar observaciones bajo el microscopio de luz, se observó además una disminución en el desarrollo y en la densidad del micelio en la zona de confrontación.
Tabla 4. Efecto antagonista de cepas de B. thuringiensis en el crecimiento radial, tasa de crecimiento y porcentaje de inhibición micelial de F. oxisporum.
Table 4. Antagonistic effect of B. thuringiensis strains in radial growth, growth rate and percentage of mycelial growth inhibition of F. oxisporum.

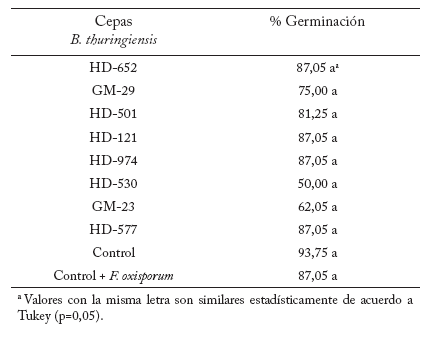
Efecto de B. thuringiensis sobre la germinación de semillas de chile. Las cepas de B. thuringiensis sólo controlaron significativamente (p=0,05) el porcentaje de germinación de las semillas infestadas con R. solani (Tabla 5). Los porcentajes de germinación para las distintas cepas de B. thuringiensis (en el control de R. solani) variaron entre 100% (GM-52) a un 25,0% (HD-597). Las siguientes cepas se destacaron por los altos porcentajes de germinación registrados: GM-6 (93,75%), HD-571, GM-24, HD-203, y GM-64 (87,5%), GM-23, HD-263, HD-652, GM-63 y HD-121 (75,0%). El resto de los tratamientos (efectos antagonistas de B. thuringiensis contra P. capsici y F. oxisporum) no fueron estadísticamente diferentes respecto al control (Tablas 6 y 7).
Tabla 5. Efecto de cepas de B. thuringiensis en la germinación de semillas de chile inoculadas con R. solani.
Table 5. Effect of B. thuringiensis strains on germination of chili pepper seeds inoculated with R. solani.

Tabla 6. Efecto de cepas B. thuringiensis en la germinación de semillas de chile inoculadas con P. capsici.
Table 6. Effect of B. thuringiensis strains on germination of chili pepper seed inoculated with P. capsici.

Tabla 7. Efecto de cepas B. thuringiensis en la germinación de semillas de chile inoculadas con F. oxisporum.
Table 7. Effect of B. thuringiensis strains on germination of chili pepper seeds inoculated with F. oxisporum.
DISCUSIÓN
La prueba de cultivo duales es extensamente usada como una de las pruebas preliminares in vitro para seleccionar agentes de control biológico (Desai et al., 2002). En el presente estudio, la prueba de cultivos duales mostró que de las 64 cepas de B. thuringiensis evaluadas, 34 presentaron diferentes capacidades para inhibir a R. solani, P. capsici y F. oxysporum; Gm-64 produjo mayor inhibición de crecimiento (66,66%) contra R. solani. Las cepas HD-263, GM-63, HD-974, GM-6 y GM- 11 fueron comunes para R. solani y P. capsici; HD-652, HD- 121 y GM-23 para R. solani y F. oxisporum, y HD-974 para R. solani, P. capsici y F. oxisporum.
Está ampliamente documentado que cepas de una misma especie, pueden exhibir diferentes capacidades para producir toxinas e inhibir el crecimiento de diferentes microorganismos (Di Pietro et al., 1991; Choi et al., 1999). Antibiosis es generalmente el modo de antagonismo observado en Bacillus spp. (Edwards et al., 1994).
La mayoría de los Bacillus spp. producen muchas clases de antibióticos como bacilomicina, fungimicina, micosubtilina y zwittermicina, que son efectivos en suprimir el crecimiento de patógenos in vitro y/o in situ (Pal y Gardener, 2006).
De hecho, Bacillus sp. actúa vía antibiosis, competencia por nutrientes, sitios de exclusión e infección, parasitismo y/o inducción de resistencia (Jacobsen y Backman, 1993; Arras y Arru, 1997; Kloepper et al., 2004). El efecto inhibidor de las cepas de B. thuringiensis en hongos fitopatógenos se puede asociar a la producción de enzimas que pueden actuar contra la pared celular. Esto es debido a que algunas bacterias antagónicas de hongos fitopatógenos producen quitinasas (Mavingi y Heulin, 1994; Asaka y Shoda, 1996). En México se han seleccionado y caracterizado las enzimas quitinoliticas de B. thuringiensis, y han llegado a la conclusión que la acción sinérgica entre las quitinasas y las proteínas Cry se puede utilizar en el control biológico de fitopatógenos (Barboza et al., 1999). Fenómenos similares fueron observados por Basha y Ulaganathan (2002). Estos autores encontraron, vía microscopio de luz, que la interacción in vitro de Bacillus sp. (cepa BC121) contra Curvularia lunata, produjo la presencia anormal de hifas, condensación, deformación y ocurrencia de malformaciones extensivas y daños al micelio. Emmert y Handelsman (1999) informaron que Bacillus cereus modificó la composición iónica del medio de cultivo en el que fue cultivado: aumentó el pH, secuestró calcio y excretó amonio. Esta combinación es altamente tóxica a las zoosporas de Oomycetes patógenos, causando un rápido ensanchamiento y expulsión de la vacuola, seguido de lisis de las zoosporas. El tratamiento de semillas de chile con B. thuringiensis fue favorable, ya que incrementó de manera significativa el porcentaje de germinación. Resultados similares han sido previamente observados en varias especies de Bacillus (Raupach y Kloepper, 1998).
Se debe destacar el comportamiento de las cepas sobre los diferentes fitopatógenos, ya que algunas resultaron ser sobresalientes por su efecto antifúngico contra alguno de los hongos, no teniendo el mismo efecto contra otros. Por ejemplo, la respuesta de la cepa GM-6 (93,75 % de germinación) a R. solani, versus su respuesta a P. capsici (87,05% de germinación).
De la misma manera, la cepa HD-974 de B. thuringiensis produjo un 62,05% de germinación en presencia de R. solani; 75% en presencia de P. capsici y 87,5% en presencia de F. oxisporum, Estos resultados sugieren la variabilidad existente entre cepas, aunque algunos autores (Schippers et al, 1987) reportan la especificidad de los antibióticos producidos por bacterias. Bettiol y Lazaretti, en 1997 probaron metabolitos de B. subtilis en semillas de frijol, mostrando una disminución en la incidencia de R. solani; sin embargo, en semillas de arroz no se observó un efecto similar. En el caso de Fusarium spp, los autores notaron una disminución de su incidencia en semillas de arroz, aunque no se observaron resultados positivos en semillas de frijol. Los resultados del presente estudio indicaron la eficacia y el efecto antifúngico en ensayos in vitro, y las potencialidades del tratamiento de semillas con cepas de B. thuringiensis.
AGRADECIMIENTOS
Los autores agradecen a la Fundación Produce Durango A.C. el financiamiento de este trabajo como parte de los proyectos 10-2005-4945 y 10-2008-0386.
REFERENCIAS
1. Arras, G. y S. Arru (1997). Mechanism of action of some microbial antagonists against fungal pathogens. Annali di Microbiologia ed Enzimologia 47: 97-120. [ Links ]
2. Asaka, O. y M. Shoda (1996). Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Applied and Environmental Microbiology 62: 4081-4085. [ Links ]
3. Barboza, C.J.E., J.C. Contreras, R.R. Velásquez, J.M. Bautista, R.M. Gómez, C.R. Cruz y J.E. Ibarra (1999). Selection of chitinolytic strains of Bacillus thuringiensis. Biotechnology Letters 21: 1125- 1129. [ Links ]
4. Basha, S. y K. Ulaganathan (2002). Antagonism of Bacillus species (strain BC121) towards Curvularia lunata. Current Science 82: 1457-1463. [ Links ]
5. Bettiol, W. y E. Lazzaretti (1997). Tratamento de sementes de arroz, trigo, feijão e soja com um produto formulado à base de células e de metabólitos de Bacillus subtilis. Scentia Agrícola 54: 89-96. [ Links ]
6. Choi, K.C., C. Young., S.H. An y B. Yook (1999). Effects of antagonistic bacteria and soil borne pathogenic fungi on growth of pasture plant seedlings. Korean Journal of Dairy Science 21: 41-48. [ Links ]
7. De Barjac, H. y J. Lajudie (1974). Mise en evidence de facteurs antagonists du type des bacteriocines chez Bacillus thuringiensis. Annals Microbiology 125: 529-537. [ Links ]
8. Desai, S., M.S. Reddy y J.W. Kloepper (2002). Comprehensive testing of biocontrol agents. En: Gnanamanickam, S.S. (ed). Biological Control of Crop Diseases. Marcel Dekker, Inc., New York·Basel, USA. pp. 387-420. [ Links ]
9. Di Pietro, D., M.R. Gut., J.P. Pachlatko y A. Schwinn (1991). Role of antibiotics produced by Chaetomium globosom in biocontrol of Pythium ultimum, a causal agent of damping-off. Phytopathology 82: 131-135. [ Links ]
10. Edwards, S.G., T. McKay y B. Seddon (1994). Interaction of Bacillus species with phytopathogenic fungi-Methods of analysis and manipulation for bicontrol purposes. En: Blakeman J.P. and Williamson B. (ed). Ecology of Plant Pathogens. CAB International, Wallingford, Oxon, UK. pp. 101-118. [ Links ]
11. Emmert, E.A.B. y J. Handelsman (1999). Biocontrol of Plant Disease: A (Gram-) positive perspective. FEMS Microbiology Letters 171: 1-9. [ Links ]
12. FAOSTAT (2005). Base de datos de estadísticas agrícolas (verificado el 29 de noviembre de 2007). http://faostat.fao.org/site/340/ DesktopDefault.aspx?PageID=340. [ Links ]
13. Gelernter, W. y G.E. Scwab (1992). Transgenic bacteria, virus, algae and other microorganisms as Bacillus thuringiensis toxins delivery systems. En: Entwhiste, P., Bailey, M.J., Cory, J. and Higgs, S. (Eds). Bacillus thuringiensis: Its Uses and Future as a Biological Insecticide. Wiley and Sons. New York, N. Y. pp. 78-105. [ Links ]
14. Guigón, L.C. y P.A. González (2001). Estudio regional de las enfermedades del chile (Capsicum annuum L.) y su comportamiento temporal en el sur de Chihuahua, México. Revista Mexicana de Fitopatología 19: 49-56. [ Links ]
15. He, H., L.A. Silo-Su., J. Handelsman y J. Clardy (1994). Zwittermicin A, an antifungal and plant protection agent from Bacillus cereus. Tetrahedron Letters 35: 2499-2502. [ Links ]
16. Jacobsen, B.J. y P.A. Backman (1993). Biological and cultural plant disease controls: Alternatives and supplements to chemicals in IPM systems. Plant Disease 77:311-315. [ Links ]
17. Kloepper, J.W., C.M. Ryn y S. Zhang (2004). Induced systemic resistance and promotion of plant growth by Bacillus sp. Phytopathology 94: 1259-1266. [ Links ]
18. Martin, P.W. y R.S. Travers (1989). Worldwide abundance and distribution of Bacillus thuringiensis isolates. Applied and Environmental Microbiology 55: 2437-2442. [ Links ]
19. Mavingui, P. y T. Heulin (1994). In vitro chitinase and antifungal activity of a soil, rhizosphere and rhizoplane population of Bacillus polymyxa. Soil Biology and Biochemistry 26: 801-803. [ Links ]
20. Meadows, M.P., D.J. Ellis, J. Butt., P. Jarret y H.D. Burges (1992). Distribution, frequency,and diversity of Bacillus thuringiensis in animal feed mill. Applied and Environmental Microbiology 58: 1344-1350. [ Links ]
21. Morgan, F.L. (1963). Infection inhibition and germ-tube lysis of three cereal rusts by Bacillus pumilus. Journal of Bacteriology 53: 1346-1348. [ Links ]
22. Osburn, R.M., J.L. Milner., E.S. Oplinger., R.S. Smith y J. Handelsman (1995). Effect of Bacillus cereus UW85 on the yield of soybean at two field sites in Wisconsin. Plant Disease 79: 551-556. [ Links ]
23. Pa, K.K. y B. M. Gardener (2006). Biological control of plant pathogens. En: The Plant Health Instructor DOI: 10.1094/PHIA- 2006-1117-02. [ Links ]
24. Parra, G. y J. Ristaino (2001). Resistance to Mefenoxam and Metalaxyl among field isolates of Phytophthora capsici causing Phytophthora Blight of bell pepper. Plant Disease 85: 1069-1075. [ Links ]
25. Pendleton, I.R. (1968). Ecological significance of antibiotics of some varieties of Bacillus thuringiensis. Journal of Invertebrate Pathology 13: 225-240. [ Links ]
26. Pérez, M.L., O.L. Durán., M.R. Ramírez., P.J.R. Sánchez y P.V. Olalde (2003). Compatibilidad fisiológica y sensibilidad a fungicidas de aislamientos de Phytophthora capsici Leo. Revista Mexicana de Fitopatología 21: 19-25. [ Links ]
27. Raupach, G.S. y J.W. Kloepper (1998). Mixtures of plant growth promoting rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology 88: 1158-1164. [ Links ]
28. Redondo, E. (1974). Estudio Preliminar en la Obtención de Posibles Plantas Diferenciales Para Agrupar las Razas Patogénicas del Hongo Phytophthora capsici Leonian. Tesis de Maestría. Escuela Nacional de Agricultura. Colegio de Postgraduados. Chapingo, México. [ Links ]
29. Reedy, M.S. y J.E. Rahe (1989). Bacillus subtillis B-2 and selected onion rhizobacteria in onion seedling rhizospheres: effects on seedling growth and indigenous rhizosphere microflora. Soil Biology and Biochemistry 21: 379-383. [ Links ]
30. Ristaino, J.B. (1991). Influence of rainfall, drip irrigation and inoculum density on the development of Phytophthora root and crown rot epidemics and yield in bell pepper. Phytopathology 81: 922-929. [ Links ]
31. Rowe, G.E. y A. Margaritis (1987). Bioprocess developments in the production of biosecticides by Bacillus thuringiensis. Critical Reviews Biotechnology 6: 87-123. [ Links ]
32. Rytter, J.I., F.L. Lukezic., R. Craig y G.W. Moorman (1989). Biological control of geranium rust by Bacillus subtillis. Phytopathology 79: 367-370. [ Links ]
33. Schippers, B., A.W. Bakker y P.A. Bakker (1987). Interactions of deleterious and beneficial rhizosphere microorganisms and the effect of cropping practices. Annual Review of Phytophatology 25: 339-358. [ Links ]
34. Smith, R.A. y G.A. Couche (1991). The phylloplane as source of Bacillus thuringiensis variants. Applied and Environmental Microbiology 57: 311-315. [ Links ]
35. Smith, R.P., M.J. Havey y J. Handelsman (1993). Suppression of cottony leak of cucumber with Bacillus cereus strain UW85. Plant Disease 77: 139-142. [ Links ]
36. Turner, J.T. y P.A. Backman (1991). Factors relating to peanut yield increases after seed treatment with Bacillus subtillis. Plant Disease 75: 347-353. [ Links ]
37. Valadares, I.M.C., M.T. Souza y W. Shiler (1998). Engenharia genética de microorganismos agentes de controle biológico. En: Melo, I. S. de; Azevedo, J. L. de (Eds.) Controle biológico. Jaguariúna: Embrapa-CNPMA, V.1, p. 208-216. [ Links ]
38. Vankova, J. (1957). Study of the effect of Bacillus thuringiensis on insect. Folia Biologica 3:175-182. [ Links ]
39. Velásquez, V.R., A.M.M. Medina y R.J.J. Luna (2001). Sintomatología y géneros de patógenos asociados con las pudriciones de la raíz del chile (Capsicum annuum L.) en el norte centro de México. Revista Mexicana de Fitopatología 19: 175-181. [ Links ]

