










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.79 no.2 Vicente López jul./dic. 2010
ARTÍCULOS ORIGINALES
Analysis of methodologies for the study of composition and biochemical carbohydrate changes in harvest and postharvest onion bulbs
Análisis de metodologías para el estudio de la composición y cambios bioquímicos de carbohidratos en bulbos de cebolla a la cosecha y postcosecha
Abrameto1 MA, CM Pozzo Ardizzi2, MI Gil2, LM Molina1
1 Universidad Nacional de Río Negro. Garrone Nº 181, 8500 Viedma.
2 Universidad Nacional del Comahue. Ayacucho y Esandi, 8500 Viedma, Río Negro, Argentina.
Address Correspondence to: Dra. Mariza Abrameto, e-mail: marizaabrameto@hotmail.com, phone: 02920-428969.
Recibido - Received 27.V.2008.
Aceptado - Accepted 7.II.2010.
Abstract. Costly investments in storage and transport of onion bulbs makes it increasingly important to identify cultivars with the best chance of long-term storage. This paper discusses our own and other researchers' results and laboratory procedures on onion's pungency, soluble solids, dry matter, respiration rate, carbohydrates and catabolism-related enzymes under different storage conditions. The variability in parameters such as carbohydrate composition, pungency, soluble solids and soluble uronic acid concentration in water is growth dependent, and defines the quality of onion bulbs at harvest time. During storage, these parameters can be modified by environmental conditions, presence of pathogens or termination of dormancy. Under postharvest conditions, indicators of dormancy are usually measured as percentage of either root emergence or sprouting. Biochemical and physiological parameters such as decreases in sucrose levels and increases in (1) respiration rates, (2) exo hydrolase activity, and (3) bulb-softening enzyme rates (polygalacturonase poly methyl esterase) signal the end of dormancy. However, more research is needed on environmental and chemical factors that reduce the rate of bulb sprouting.
Keywords: Allium cepa L.; Respiration rate; Sprouting; Ethylene; Non-structural carbohydrates; Enzyme activity.
Resumen. Costosas inversiones en almacenamiento y transporte de bulbos de cebolla destacan la importancia de identificar cultivares con las mejores posibilidades de almacenamiento a largo plazo. En este trabajo se discuten procedimientos de laboratorio y resultados propios u obtenidos por otros autores que incluyen la cuantificación de pungencia, sólidos solubles, materia seca, tasa de respiración, carbohidratos, y enzimas relacionadas con el catabolismo en bulbos de cebolla expuestos a diferentes condiciones de almacenamiento. La variabilidad en parámetros como la composición de carbohidratos, pungencia, sólidos solubles y concentración de ácido urónico soluble en agua es cultivar dependiente, y define la calidad de los bulbos de cebolla en el momento de la cosecha. Dicha calidad puede ser modificada por las condiciones ambientales, presencia de patógenos o fin de la dormancia durante el almacenamiento. En post-cosecha, los indicadores del estado de dormancia son usualmente medidos como porcentaje de emergencia de raíces o de brotación. Señales bioquímicas y fisiológicas tales como disminuciones en los niveles de sacarosa, o incrementos en (1) la tasa de respiración, (2) la actividad exohidrolasa, o (3) las tasas de ablandamiento del bulbo por las enzimas polimetilesterasa y poligalacturonasa señalan el fin de la dormancia. Sin embargo, es necesario continuar investigando acerca de los factores ambientales y químicos que disminuyen el porcentaje de brotación.
Palabras clave: Allium cepa L.; Tasa de respiración; Brotación; Etileno; Carbohidratos no estructurales; Actividad enzimática.
Introduction
Many commercial onions (Allium cepa L.) are stored before marketed, but cultivars differ in their storage capability. The aim of onion bulb storage is to cover consumer demands and extend the availability of onions in time, keeping quality of product at the same time. Main factors which produce deterioration of onion bulbs during storage are (1) the pre and postharvest environmental conditions (O'Connor, 1979), and (2) biological factors (like respiration, resumption of growth and pathogen attack). Class "I" onions must not show any signs of external sprouting (Regulation Commission 1508/2001/EEC). Early signs of external shoot growth are allowed in Class "II" onions provided that either the number or weight does not exceed 10% per unit of presentation (Chope, 2006). Bulbs with any scale of water and/or bacterial or fungal rots are considered inadequate for marketing (Pozzo Ardizzi et al., 2008).
Many biochemical changes take place during storage, and some of them are linked to bulb's respiration. These changes can be in water (Corgan & Kedar, 1990) and carbohydrate content, flavor compound concentration, and plant growth regulators (Chope et al., 2006).
It is important to maintain the air relative humidity (RH) low enough to avoid or control pathogen's development (roughly < 80% RH).
Cultivar specific weight losses between 2-5% of fresh weight/ month have been recorded during the first month of storage (Koopsell & Randle, 1997; Gubb & MacTavish, 2002).
The relatively low initial rate of weight loss represents loss of water through the skin, and low-level respiration rates of dormant bulbs. This is followed by a change to steeper slopes, indicating more rapid weight losses, associated with sprout growth initiation and the senescence of older, fleshy scales (Gubb & MacTavish, 2002).
Sugar metabolism is linked to bulb dormancy and the state of sprouting (Kato, 1966). Variations in monosaccharide and disaccharide levels have been previously reported in onion bulbs during storage (Suzuki & Cutcliffe, 1989; Benkeblia et al., 2002; Benkeblia et al., 2004). However, the relationship between variation in sugar levels and physiological factors is not clearly understood.
Fructans are accumulated during the bulb developmental stage. Then, they are metabolized during regrowth and sprout of the bulbs. However, the enzymatic processes involved, or the mechanisms by which they contribute to long bulb maintenance are still poorly understood (Shiomi et al., 2005).
Another change in the bulb biochemistry that modifies its quality in storage is the softening of tissues. This is produced by the hydrolysis of the pectin portion of the bulb cell walls through pectolitic enzymes.
This paper summarizes some methods developed to quantify mono and disaccharides, fructooligosaccharides and fructans, respiration rates, and enzymes such as polygalacturonase or fructo-hydrolases. This allows to get information about compositional changes produced during storage that modify onion quality.
Pungency and soluble solid content
Onions give off a very strong smell when they are cut. This is because the cellular disruption releases the enzyme allinase from the vacuoles, and disruption of sulfur-amino acids [known as S-alk(en)yl cysteine sulfoxides (ACSOs)] take place in the cytoplasm (Block, 1992). This reaction produces three natural compounds: 1-propenyl cysteine sulfoxide (1-PrenCSO) that is produced in high concentrations and dominates the flavour profile, Methyl cysteine sulfoxide (MCSO), and Propyl cysteine sulfoxide (PCSO). Two measurable intermediates leading to ACSO synthesis are S-2-carboxypropyl glutathione (2carb) and γ-L-glutamyl-S-(1-propenyl)-L-cysteine sulfoxide (GPECSO). Pyruvate and ammonium are non-flavour products of this enzymatic reaction (Block, 1992).
Pyruvic acid level, as an index of onion pungency, has become a routine procedure to ensure the quality of onions for both shippers and consumers.
Pyruvate produced by hydrolysis of the ACSOs can be estimated using dinitro phenyl hydrazine (DNPH) (Schwimmer & Weston, 1961). Absorbance of pyruvate phenilhydrazone is measured spectrophotometrically at 420 nm after mixing onions with water (1+1), filtering, centrifugation and dilution before analysis. Randle & Bussard (1993 b) developed a fast juice extraction method using a pneumatic press, eliminating blending bulb tissues with water. Several drops of fresh juice can be used to measure soluble solid content with an optical refractometer (Kopsell & Randle, 1997).
Yoo & Pike (1999) used the same principle as the original Schwimmer and Weston (1961) method. Included in the system, there are two high performance liquid chromatography (HPLC) pumps, an autosampler, a column heater, a spectrophotometer detector and an integrator. This automated system showed highly significant correlations with HPLC and the spectrophotometric method.
Recent technologies have developed meldolas blue electrodes. These have been considered the best sensors for detection of pyruvate with amperometers in untreated onion juice (Abayomi et al., 2006). There was a strong correlation between the sensor response and pyruvate concentration from onion cultivars (2-12 μmol/g FW), measured with the modified Schwimmer & Weston (1961) spectrophotometric assay.
After alliinase is deactivated using a microwave, the background level of pyruvic acid in onion can be measured. Recent works have confirmed that these background levels were minimal (Allium species are similar and do not normally exceed 1 μmol/ml), and can be disregarded in routine pungency analysis (Yoo & Pike, 2000; Randle & Bussard, 1993a).
Pungency values [from enzymatically developed pyruvate (EPY)] have ranged between 3-17 μmol/g of fresh tissue (Randle, 2000). However, values above 9 μmol/g are considered extremely pungent.
Pungency can differ among cultivars before and during storage. Among new cultivars, EPY either decreased or increased linearly with storage duration. In intermediate and long-day cultivars, grown and stored under the same environmental conditions (bulbs harvested from greenhouse-grown plants, cured and stored for 2 to 6 month at 5 °C), EPY decreased linearly or quadratically during storage while soluble solids decreased linearly over time. In storage at low O2 (2.0 kPa) and high CO2 (8.08 kPa) concentration, the activity of alliinase might decrease (Uddin & Mac Tavish, 2003).
In the Valle Inferior del Rio Negro, Patagonia, Argentina, onion bulbs are accumulated in the field until their sale. However, good quality of onion bulbs during the storage period cannot be assured. Losses caused by post-harvest onion diseases ranged between 20 to 90% depending, among other things, of the general bulb state when they get into the piles (maturity, neck closure, impurities); they also depend upon the care put into the construction of the structures (orientation, ventilation, Pellejero et al., 2005).
There were significant differences in pungency between onion cultivars (p<0.001, d.f.: 4.60) and storage period (p<0.001, d.f.: 3.60) treatments when five cultivars with a storage period of 90 days were compared in 2007 (Fig. 1). The interaction test was significant (p<0.001, d.f.: 12.60). This indicates a different effect of the concentration of pyruvate in each cultivar at each storage time. After 30 days of storage, all cultivars increased the pyruvate concentration. Antartica Inta (A. Inta) classified as highly pungent, while the remaining cultivars maintained moderate pungency until the end of the storage period (Fig. 1).
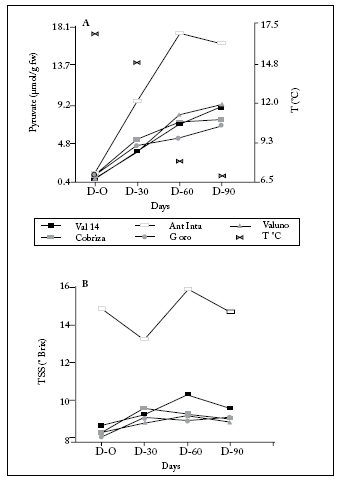
Fig. 1. (A) Changes in Pungency and mean monthly temperatures, and (B) Total Soluble Solids (TSS) in onion bulb cultivars stored during 90 days under field conditions (n=4, 10 bulbs for each sample).
Fig. 1. (A) Cambios en Pungencia y temperaturas medias mensuales, y (B) Sólidos Solubles Totales (TSS) en bulbos de cultivares de cebolla almacenados durante 90 días en condiciones de campo (n=4, 10 bulbos por cada muestra).
Total soluble solid (TSS) concentration changed significantly over time. Two-ways-ANOVA showed a main effect of cultivar (p<0.001, d.f.: 4.60) (Fig. 1). The highest TSS concentration was observed in A. Inta, and the lowest in Grano Oro (G Oro) (Tukey test, p<0.05, d.f.: 72). There was no evidence of interaction between (1) onion line and (2) storage time on bulb. G Oro and Cobriza showed a maximum TSS concentration after 30 days of storage. Valuno and Valcatorce showed highest values after 60 days, although these changes were not significantly different. There was only an increase in bulbs of A Inta between 30 and 60 days of storage (Tukey test p=0.05, d.f.: 12) (Fig. 1).
Respiration and sprout suppression
During the post harvest period, producers try to conserve the bulbs under controlled environmental conditions to avoid or minimize chemical and physical changes. Management of temperature and relative humidity can decrease the general metabolism rate; especially, the respiration rate under air conditions.
Respiration rates can be measured using whole onions, without any cold treatment. This can be achieved either by (1) the glass jar technique, using gas chromatography to measure the carbon dioxide produced in the closed system, or (2) oxygen removal from the storage environment (Benkeblia& Shiomi, 2004). At harvesting time, and at several post-harvest intervals, bulbs can be incubated in glass vessels during 2 to 5 hours, under air conditions at 5 °C. After this incubation period a gas sample is removed and run by the gas chromatography (Peiris et al., 1997). The GC can be calibrated with 10% CO2, 2% O2, 88% N2, employing the hot wire detector (Chope et al., 2007).
The respiration rate is expressed as mmol CO2/kg/h. Thus, if we know the rate of dry matter loss (expressed in glucose g/day/kg fresh weight), respiration rate can be estimated, considering that for each 180 g of glucose oxidized, 264 g of carbon dioxide are produced. Then, the potential storage life of the product can be estimated. An increase in respiration rate corresponds to the break of dormancy, and the start of sprouting even though the sprouts have not emerged from the necks. Some measures of mean respiration rates were in the order of 0.1655, 0.0999 and 0.1045 mmol CO2/kg/h on onion cultivars SS1, Carlos, and Renate, respectively (Chope et al., 2007).
Pegasus cultivar had 60% sprouting at harvesting time while other varieties required 60 days of storage at 6.6 °C and 82% air relative humidity to reach similar levels (Coolong et al., 2008). The time taken for 50% bulb sprouting at a storage temperature of 4 °C was 30 days in cultivars SS1 and 107 days in cultivar Hysam; an increase in storage temperature to 20 °C reduced this time between 7 to 65 days, respectively (Chope et al., 2007). Benkeblia & Shiomi (2004) reported that the respiration rate of sprouted onions was 52% higher than that in freshly harvested and dormant bulbs. Respiration rates of Rouge Amposta bulbs were 0.08 mmol CO2/kg/h, 60 days after transfer from 0 °C (during 4 weeks) to 20 °C in darkness.
Use of controlled atmospheres (CA) (e.g., 5 kPa CO2 and 3 kPa O2) in the storage environment can inhibit sprouting of the bulbs and reduces weight loss (Yoo & Pike, 1996). Onions removed from CA after a 20-day-treatment had greater respiration rates (0.1822 mmol CO2/kg/h) than those removed from air storage (0.0703 mmol CO2/kg/h) (Chope et al., 2007).
Abdel-Rahman & Isenberg (1974) reported that endogenous ethylene increased at the end of dormancy suggesting that ethylene may have a role in sprouting. 1-methylcyclopropene (1-MCP) binds to ethylene-binding proteins, then preventing ethylene effects. Sprouting length, expressed as a percentage of bulb height, was smaller in onions SS1 treated with 1-MCP than in untreated onions stored at 4-12 °C, but not when they were stored at 20 °C (Chope et al., 2007; Chope et al., 2008). Continuous exogenous ethylene treatment in bulbs of cv "Copra" inhibited sprouting in onions stored at 18 °C by inhibiting leaf blade elongation. This effect disappeared when ethylene was removed. In contrast, treatment of dormant bulbs with 1-MCP caused breaking of dormancy 2 weeks before natural dormancy (Bufler, 2009).
Non-structural carbohydrates
Non-structural carbohydrates (NSC) in Allium spp include sucrose, fructose, glucose and various oligosaccharides, such as the fructans. Starch and raffinose are absent in this species (Benkeblia et al., 2002).
In cultivars with low, medium and high dry weight percentages, total non-structural carbohydrates represent a fairly constant percentage (83 to 88%) of dry weight (Darbyshire& Henry, 1979).
Fructans are the reserve polysaccharides in several alliance species, including onion. These are a series of homologous oligo and polysaccharides of fructose, which are formed by the successive additions of fructosyl groups, bounded by β linkage at the position of sucrose. The degree of polymerization (DP) can vary among Allium's species. In onion, the fructans DP is between 3 and 15 (GFn, G: glucosyl, F: fructosyl, n>2). The DP is correlated with dry matter (DM) content; low-DP of DM content are <10%, while maximum DP of DM% are 15- 20 (Ernst et al., 1998).
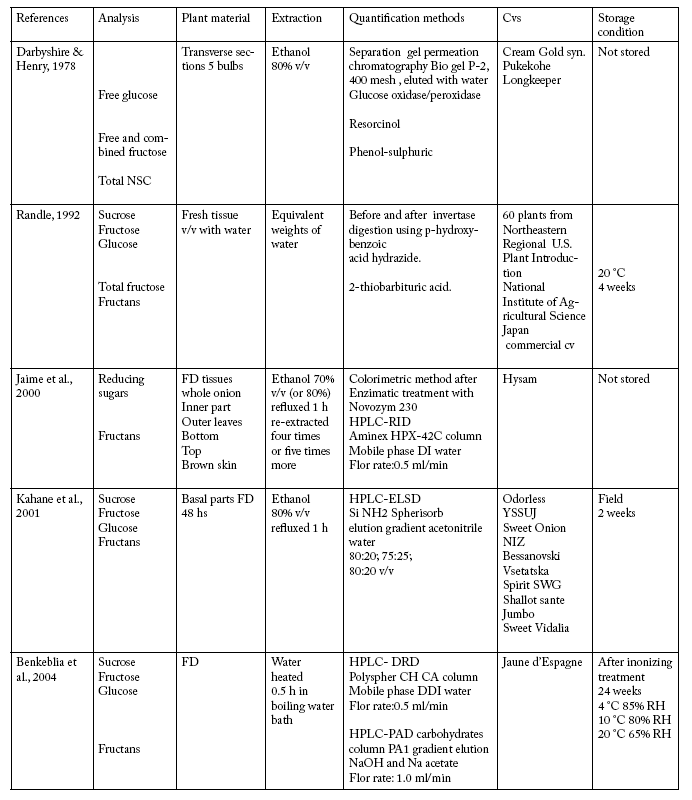
Table 1 lists papers published on the analysis of nonstructural carbohydrates from onion, indicating the applied methods. Total NSC ranged from 20.1 to 3.08 g/100 g fresh weight (FW) (Randle, 1992). These concentrations are representative of genotypic differences previously reported for dry matter accumulation in onions (Darbyshire & Henry, 1979). Results from total NSC measured colorimetrically on marketable cultivars developed in Argentina were from 1.39% to 2.94% (FW) at harvest. After 3 months of storage under field conditions, all cultivars had increased significantly in this content: 4.9 % to 6.8% (FW) (Pozzo Ardizzi et al., 2005).
Table 1. Methods developed for the analysis of non structural carbohydrates from onion in the last thirty years.
Tabla 1. Métodos desarrollados en los últimos 30 años, para el análisis de carbohidratos no estructurales de cebolla.

Glucose, fructose and sucrose have been separated either by using High-performance liquid chromatography (HPLC) and a differential refractometer detector (Benkeblia et al., 2004, Benkeblia & Shiomi, 2004; Jaime et al., 2002), or determined by HPLC and monitored by light-scattering detectors (LSD) (Kahane et al., 2001, Terry et al., 2005).
In the United Kingdom, cultivars SS1, ESS1, and Buffalo were classified as LDM, low pungency onions, monosaccharides- dominated. At the same time, Domenica, Element, Radar, and Shakespeare were HDM, medium to high pungency, disaccharide-dominated (Terry et al., 2005, Davis et al., 2007).
In 2007, we used the modified procedure of capillary GC analysis of major sugars from onion extracts following Chapman & Horvath (1989). Cultivars Valcatorce Inta and Grano de Oro had similar characteristics and showed differences in fructose and glucose levels with respect to Antártica INTA. At the same time, Antártica INTA showed evidence of significant differences in high soluble solids and low NSC with respect to the previous group (Table 2) (Gil et al., 2009).
Table 2. Mean concentration of reducing sugars, sucrose, soluble solids and pungency in onion bulbs under pre-storage conditions in the Low Valley of Rio Negro.
Tabla 2. Concentraciones medias de azúcares reductores, sacarosa, sólidos solubles y pungencia en bulbos de cebolla antes del almacenamiento en el Valle Inferior de Río Negro.

The fleshy part of the onion bulb is the richest tissue in NSC. In some cultivars, fructans are the main carbohydrates, increasing from the outer to the inner fleshy leaves (the youngest leaves). The reducing sugar/sucrose ratio is highest in the older, outer leaves when comparison is made with the inner, fleshy leaves (Jaime et al., 2000; Darbyshire & Henry, 1978).
There are different results regarding the relation between NSC storage patterns and DM. Kahane et al. (2001) showed in low and high DM accessions that cultivars expressed different profiles of NSC for similar DM. They speculated that this was probably based on a genetic regulation for fructan accumulation, complementary to osmoregulation of the cell (Derbyshire & Henry, 1979). High DM cultivars showed highest accumulation of fructans over whole bulbing period, whereas low DM cultivars quickly reached a plateau (Kahane et al., 2001). As DM and NSC distributions before bulbing were similar for all accessions, they suggested that it would be better to distinguish differences between fructan accumulator cultivars versus non-accumulators. However, Sinclair et al. (1995) obtained different results. Matrix correlations had shown negative correlations between DM and FRU/GLU, but positive correlations between DM and fructans (Kahane et al., 2001; Randle, 1992).
At present, new techniques such as (1) HPLC with refractive index or pulse amperometric detection, (2) mass spectrometry (Jaime et al., 2000), or (3) fructan assay kit (Megazime, Co. Wicklow, Ireland) (O'Donoghue et al., 2004) have been used to quantify fructans and fructoologosaccharides (FOS).
After 6 months of storage in a refrigerated chamber at 15 °C and 45% relative humidity, total both FOS and carbohydrates showed similar variation. Total FOS decreased progressively at an average rate of 1072 μg/g fresh weight/week. Total carbohydrates increased during the first 6 weeks, but after that they decreased progressively at an average rate of 1386 μg/g fresh weight/week (Benkeblia et al., 2005).
The total concentration of simple sugars can be calculated by adding the concentrations of fructose, glucose and sucrose. Chope et al. (2006) reported a total sugar (sucrose+glucose+fructose) decrease during storage at (1) temperatures between 4°C to 20 °C, and (2) two fold greater concentrations in 1-MCP (1-methylcyclopropene) -treated onion stored at 12 °C than on untreated bulbs in cultivar SS1.
Fructan hydrolysis during storage is catalyzed by FEH (fructan exohydrolases) that remove fructosyl residues from the non-reducing end of fructan molecules, and occurs with increasing activities from internal to external leaves. Exohydrolases are β-(2-6) linkage specific, and the distribution of fructan obtained by hydrolysis has occurred for 1-ketose (GF2), nystose (GF3) and 1-β-D-fructofuranosylnistose (GF4) (Kaack et al., 2004).
The fructose content can be quantified by GC using the carbohydrate Chapman and Horvath's modified procedure as was described previously (Chapman & Horvath, 1989).
The extraction of 1-fructan exohydrolase can be measured from fresh bulb tissue with Citrate/Phosphate buffer (pH 5.0). The slurry must be filtered and centrifuged in ultra centrifuge. The supernatant must be filtered again through Microcon centrifugal device (10,000 Molecular Weight cut off). After washing the filter with Citrate/Phosphate buffer, the protein can be suspended in Citrate/Phosphate buffer, pH 5.0 (Coolong, personal communication).
To assay the enzyme activity, it needs to be incubated at 30 °C for 2 hours in Inulin in a citrate/phosphate buffer. To stop the reaction after this time, heat in a boiling water bath. Standard Phenyl β-D phenyl glycoside and samples must be boiled, and continue with the three steps to carbohydrates analysis.
Few investigations are focused on FEH in onion bulbs. Benkeblia et al. (2005) characterized enzyme activities during postharvest time by a peak. The peaks of degrading enzymes activities (FEH) and 1-kestose hydrolyzing enzyme may be a signal of the release of bulb dormancy, after long-term storage and the start of sprouting stage (Benkeblia et al., 2005). These authors (Benkeblia et al., 2004), showed that onion bulbs with ionizing treatment, after 6 months at 20 °C, had DP 5-12 fructans that were strongly hydrolyzed in cultivar Jaune d'Espagne. Between 4°and 10 °C, slight degradation of these polymers occurred, and there was a slight accumulation of tri and tetra saccharides.
Fructan concentrations in bulb samples of cultivars Renate, Ailsa Craig and SS1, with long, medium and short storing, stored under CA conditions (2 °C, 3.03 kPa CO2 and 5.05 kPa O2) were significantly different. Cultivars with long storage had the highest proportion of fructans/g dry matter (DM) (Jaime et al., 2000; Chope et al., 2006), and these significantly changed over time. Two other cultivars were not significantly different over time (Chope et al., 2006).
Pectolytic enzymes
Textural changes may occur because insoluble polyuronide, which contributes substantially to cell wall integrity and firmness, is converted to more soluble forms (Gross, 1982). The onion cell wall at harvest is usually about 2% of fresh weight. After ten weeks of storage, 40% of the cell wall has been degraded.
That significant softening occurs in onion tissue during refrigerated storage. This transformation is facilitated by the pectolytic enzymes [e.g., pectin methyl esterase (PME, EC 3.1.1.11)]. These enzymes produce the hydrolysis of carboylix esters of polygalacturonic acid; then, the galacturonic acid chains may be hydrolized by polygalacturonase (PG, EC 3.2.1.15).
Polygalacturonase assay can be done by 2-cyanoacetamide method (Coolong et al., 2008; Gross, 1982). Polygalacturonase (PG) activity assay of onion is based on the hydrolytic release of reducing groups (D-galacturonic acid residues) from polygalacturonic acid producing tissue softening (Coolong et al., 2008).
Enzymatic extraction from fresh tissue is blended with NaCl. The slurry has to be filtered through cheesecloth, ultra centrifuged and transferred to a Millipore filtering vial for centrifugation to low temperature. The pellet has to be washed and centrifuged with Na-Acetate pH 4.4. After washing and micro centrifugation without low temperature, the proteins fall into the vial. The samples must be boiled to stop the enzymatic activity. The reaction mixtures containing enzyme extracts or blanks, (in buffer Na acetate, pH 4.4) plus 0.2% polygalacturonic acid, are incubated at 30 °C.
The amount of reducing groups formed may be calculated using a standard curve of mono D-galacturonic acid assuming that the concentration of this acid is proportional to the concentration of reducing groups. To estimate the released reducing groups, reactions are finished adding borate buffer (pH 9) and 2-cyanoacetamide in the entire vials (samples, blanks, standard) and immersing it in a boiling water bath. After equilibration to 25 °C, the absorbance is measured at 276 nm using an UV-spectrophotometer.
In tomatoes, PME has been shown to catalyze the deesterification of pectin that subsequently is a substrate for PG, catalyzing the hydrolytic cleavage of the α-1,4-glycosidic bonds. In other way, controlled activity of PME with selective PG inactivation can result in some benefits like texture improvement (Rodrigo et al., 2006).
Coolong (2008) reported that the firmest onion bulbs at harvest had the thickest cell wall, highest levels of both dry matter and total uronic acid. The poor-storing cultivars had the softest bulbs at harvest and lowest levels of uronic acid.
Remarks
Confidence in the accuracy of pyruvate measurements is becoming more important, as the popularity of low pungency onion increases (Yoo & Pike 2001). The automated pyruvic acid analysis system (Yoo & Pike, 1999) can save time and eliminate human errors because there is no need of pipetting for adding chemicals. Estimating about 17% more pyruvic acid than spectrophotometrics methods, it may overestimate pyruvic acid content by about 0.3 μmol/ml in onions with a high sugar or quercetin concentration.
The Abayomi's study (2006) has demonstrated the possibility of replacing the standard colorimetric assay used by the world onion industry (to determine pyruvate concentration) for the biosensor. The advantages are quick analysis and elimination of the use of DNPH (harmful to health). Future works should elucidate possible variations in response of interfering compounds, or cultivar interference.
Pungency differences have been attributed to metabolic dissimilarities in the S-alk(en)yl cysteine sulfoxides, associated with individual phenotypes (Kopsell & Randle, 1997). Changes in pyruvate concentrations during storage are likely different in the availability of ACSOs. ACSOs may be metabolized to supply carbon and nitrogen to growing sprouting (Uddin & Mac Tavish, 2003).
Storage tends to increase pyruvate concentration, at controlled conditions (Chope et al., 2007) or field conditions as in Argentina, which generates products more pungent than fresh ones. A minimum TSS content was only observed on cultivar A INTA after 30 days of storing bulbs at 15 °C mean monthly temperature. Similar results were reported by Chope et al. (2006, 2007).
At harvesting, and 30 and 60-day-cold-treated onion bulbs, or refrigerated storage conditions, have yielded 50%-60% breakage of bulb dormancy. This effect was reported previously by other authors (De Hertogh & Le Nard, 1993). There was an interaction between cultivar and storage time regarding sprouting or rooting (Coolong et al., 2008).
A single dose of ethylene inhibitor 1-MCP reduced sprouting in short-storing onion cultivar SS1, and increased concentration of total soluble carbohydrates (sucrose, glucose and fructose), suggesting a low rate of C utilization (Chope et al., 2008). On the other hand, exogenous ethylene supply during sprouting or during dormancy inhibited sprouting of onion bulbs, and their action was on sprout leaf elongation (Bufler, 2009). It shows that suppression of sprouting and dormancy are controlled by different physiological factors, and that the role of ethylene in dormancy needs further investigation.
LSD was chosen in sugar analysis as the preferred method for detection by refractive index due to greater baseline stability and sensitivity as compared to conventional detection by refractive index (Terry et al., 2005). Among the main advantages of the method modified by Chapman & Horvath (1989) for sugars analysis are a total GC run time of 30 min. This is sufficient to elute all the components, in comparison to that by a long time HPLC elution gradient (Kahane et al., 2001). Glucose and fructose have two peaks, α and β isomers, and they do not interfere with other sugars. More reproducibility of the results and stability of the derivatives working from dried tissues is possible. The same applies to quantitative analysis of carbohydrates at different times of storage, and measurement of enzymatic activities, such as fructan-hexohydrolase.
Both methods, colorimetric and by GC (Table 1, Table 2) showed that the total carbohydrate content gave different results with marketables cultivars. Different results indicate that the extraction procedure is crucial in the determination of the concentration and ratios of non-structural carbohydrates in onion bulbs (Davis et al., 2007).
The variation parameters such as pyruvate content, soluble solids, and NSC betwen cultivars allowed to asses a classification on bulbs quality. Valcatorce, Grano de Oro and Antártica Inta are mono-saccharides-dominated. We found two groups: Grano Oro and Valcatorce, classified as low soluble solids, and moderate pungency (Fig. 1), and high soluble solids and pungency, respectively. The concentrations of fructose and glucose (mg/g DW) found in these cultivars were similar to those reported for cultivar Sheakspeare (Terry et al., 2005). There was negligible sucrose concentration as compared to Shakespeare cultivar.
Fructan content in onion bulbs tends to decrease during refrigerated atmosphere storage (e.g., Suzuki & Cutliffe, 1989, Benkeblia et al., 2000). Suzuki & Cutcliffe (1989), however, reported slight increases in fructans DP 5-8. Chope et al. (2006) reported different patterns of change in fructan concentration, which could be due to different responses of each cultivar to environmental conditions. Despite changes in fructans, slightly and highly polymerized fructan levels, and their exohydrolase activities in post-harvests, are not well documented.
The results found in cultivar Antártica Inta (high TSS, low soluble carbohydrate total content) suggest that it is a fructan acumulator.
Firmness in vegetables is a textural attribute related to freshness of products. There has been research about pectin methyl esterase activity and polygalacturonase in different vegetables as Tabasco pepper (Arancibia & Motsenbocker, 2006) and tomatoes varieties (Rodrigo et al., 2006). However, there is limited information about these pectolytic enzymes in onion. Coolong (2008) results suggest that differences in onion scale firmness at harvest may be due to differences in water soluble pectin uronic acid concentrations.
This information could be of interest to improve onion storage management, especially in countries where these problems are related to scarce use of technology during storage.
ACKNOWLEDGEMENTS
This paper is part of a postdoctoral fellowship, and research was conducted during 2006 at the Department of Horticulture, 1111 Miller Plant Sciences, University of Georgia, Athens, Georgia U.S.A. Financial support was provided by Universidad Nacional del Comahue, Neuquén. The senior author acknowledges Director Dr. William M. Randle. Technical support by Dr. Timothy W. Coolong and Dr. Pai-Tsang Chang is also acknowledged.
REFERENCES
1. Abayomi, L.A., L.A. Terry, S.F. White & P.J. Warner (2006). Development of a disposable pyruvate biosensor to determinate pungency in onions (Allium cepa L). Biosensors and Bioelectronic of onion bulbs 21: 2176-2179. [ Links ]
2. Abdel-Rahman, M. & F.M.R. Isenberg (1974). The role of exogenous plant growth regulators in the dormancy. Journal of Agricultural Sciences 82: 113-116. [ Links ]
3. Arancibia, R. & A. Motsenbocker (2006). Pectin methyl esterase activity in vivo differs from activity in vitro enhances polygalacturonase mediated pectin degradation in Tabasco pepper. Journal of Plant Physiology 163: 488-496. [ Links ]
4. Benkeblia, N., K. Ueno, S. Onodera & N. Shiomi (2005).Variation of fructo-oligosaccharides and their metabolizing enzymes in onion bulb (Allium cepa L. cv Tenshin) during long term storage. Journal of Food Sciences 70: 208-214. [ Links ]
5. Benkeblia, N., S. Onodera & N. Shiomi (2004). Effect of gamma irradiation and temperature on fructans (fructo-oligosaccharides) of stored onion bulbs Allium cepa L. Food chemistry 87: 377-382. [ Links ]
6. Benkeblia, N. & N. Shiomi (2004). Chilling effect on soluble sugars, respiration rate, total phenolics, peroxidase activity and dormancy of onion bulbs. Sciences Agricultural (Piracicaba, Brazil) 61: 281-285. [ Links ]
7. Benkeblia, N., P. Varoquax, N. Shiomi & H. Sakai (2002). Storage technology of onion bulbs cv Rouge Amposta: effect of irradiation, maleic hydrazide and carbamate isopropyl, N-phenyl (CIP) on respiration rate and carbohydrates. International Journal of Food Sciences and Technology 37: 169-176. [ Links ]
8. Benkeblia, N., P. Varoquax, B. Gouble & G. Selselet Attou (2000). Respiratory parameters of onion bulbs. Effect of irradiation and temperature. Journal of the Science of Food and Agriculture 80: 1772-1778. [ Links ]
9. Block, E. (1992). The organosulfur Chemistry of the genus Allium. Implications for the organic chemistry of sulfur. Angewandte Chemie, International Education. English. 31: 1135-1178. [ Links ]
10. Bufler, G. (2009). Exogenous ethylene inhibits sprout growth in onion bulbs. Annals of Botany 103: 23-28. www.aob.oxfordjournals.org [ Links ]
11. Coolong, T., W. Randle & L. Wicker (2008). Structural and chemical differences in the cell wall regions in relation to scale firmness of three onion (Allium cepa L.) selections at harvest and during storage. Journal of the Science of Food and Agriculture 88: 1277-1286. [ Links ]
12. Corgan, J.N. & N. Kedar (1990). Onion cultivation in subtropical climates, p. 31-48. In: H.D. Rabinowitch and J.L. Brewster (eds). Vol 2. Onions and allied crops. CRC Press, Boca Raton, Fla. [ Links ]
13. Chapman, G. & R. Horvath (1989). Determination of Nonvolatile Acids and sugars from fruits and sweet potato extracts by capillary GLC and GLC/MS. Journal of Agriculture and Food Chemistry 37: 947-950. [ Links ]
14. Chope, G.A. (2006). Understanding the mechanisms behind onion bulb dormancy in relation to the potential for improved onion storage. Cranfield University, Plant Science Laboratory, U.K. Ph.D Thesis. [ Links ]
15. Chope, G.A., L. Terry & P. White (2006). Effect of controlled atmosphere storage on abscisic acid concentration and other biochemical attributes of onion bulbs. Post harvest Biology and Technology 39: 233-242. [ Links ]
16. Chope, G.A., L. Terry & P. White (2007).The effect of 1-methylcyclopropene (1-MCP) on the physical and biochemical characteristics of onion cv SS1 bulbs during storage. Post harvest Biology and Technology 44: 131-140. [ Links ]
17. Chope, G.A., L. Terry & P. White (2007). The effect of the transition between controlled atmosphere and regular atmosphere storage on bulbs of onion cultivars SS1, Carlos and Renate. Post harvest Biology and Technology 44: 228-239. [ Links ]
18. Chope, G. A. & L. Terry (2008). The role of abscisic acid and ethylene in onion bulb dormancy and sprout suprpression. Stewart Postharvest Review 2-7. [ Links ]
19. Darbyshire, B. & R.J. Henry (1978). The distribution of fructans in onions. New Phytologist 81: 29-34. [ Links ]
20. Darbyshire, B. & R.J. Henry (1979). The association of fructans with high percentage dry weight in onion cultivars suitable for dehydrating. Journal of the Science of Food and Agriculture 30: 1035-1038. [ Links ]
21. Davis, F., T.A. Leon, G.A. Chope & C.F. Faul (2007). Effect of Extraction Procedure on Measured Sugar Concentrations in Onion (Allium cepa L.) Bulbs. Journal of Agriculture and Food Chemistry 55: 4299-4306. [ Links ]
22. De Hertogh, A.A. & M. Le Nard (1993) Physiology and biochemical aspect of flower bulbs. In: De Hertogh A.A., Le Nard M. (Ed) The physiology of flower bulbs. Amsterdam: Elsevier, 1993. chap. 6. 53-69 p. [ Links ]
23. Ernst, M.K., N.J. Chatterton, P.A. Harrison & G. Matitschka (1998). Characterization of fructan oligomers from species of the genus Allium L. Journal Plant Physiology 153: 53-60. [ Links ]
24. Gil, MI., M. Abrameto, M.C. Pozzo Ardizzi, A. Van Koninenburg, G. Ashkar & G. Pellejero (2009). Composición de carbohidratos no estructurales en cultivares de cebolla (Allium cepa L.) del valle Inferior del río Negro. ASAHO. XXXII Congreso de Horticultura. Salta, Argentina. [ Links ]
25. Gross, K. (1982). A rapid and sensitive spectrophotometric method for assaying polygalacturonase using 2-cyanoacetamide. Hort-Science 17: 933-934. [ Links ]
26. Gubb, I.R. & H.S. MacTavish (2002). Allium crop science: Recent Advances, ed. by Ravinowitch, H.D. and Currah, L..10 Onion pre-and postharvest considerations, Wallingford, Oxon: CABI Publishing. p. 233-265. ref. (Text in En) (SB338.R3). [ Links ]
27. Jaime, L., E. Molla, A. Fernández., M. Martin Cabrejas, F. Lopez Andreu & R. Esteban (2002). Structural carbohydrate differences and potencial source of dietary fiber of onion (Allium cepa L.) tissues. Journal Agriculture and Food Chemistry 50: 122-128. [ Links ]
28. Jaime, L., F. Martinez, M. Martin Cabrejas, E. Molla, F. López Andreu, K. Waldron & R. Esteban (2000). Study of total fructan and fructooligosaccharide content in different onion tissues. Journal of the Science of Food and Agriculture 81: 177-182. [ Links ]
29. Kato, T. (1966). Physiological studies on the bulbing and dormancy of onion plants. VIII: Relation between dormancy and organic constituents of bulbs. Journal of the Japanese Society for Horticultural Science 3: 57-63. [ Links ]
30. Kaack, K., L.P. Christensen, S.L. Hansen & K. Grevsen (2004). Non structural carbohydrates in processed soft fried onion (Allium cepa L.). European Food Research and Technology 1-20. [ Links ]
31. Kahane, R., E. Vialleguerin, I. Boukema, D. Tzanoudakis, C. Bellamy, C. Chamaux & C. Kik (2001). Changes in non structural carbohydrate composition during bulbing in sweet and high-solid onions in field experiments. Environmental and Experimental Botany 45: 73-83. [ Links ]
32. Kopsell, D. & W.M. Randle (1997). Onion cultivars differ in pungency and bulb quality changes during storage. HortScience 32: 1260-1263. [ Links ]
33. O'Connor, D. (1979). Grower guide no. 2; Grower Books. London. [ Links ]
34. O'Donoghue, E.M., S.D. Somerfield, M. Shaw, M. Bendall, D. Hedderly, J. Eason & I. Sims (2004). Evaluation of carbohydrates in Pukekohe Long keeper and Grano cultivar of Allium cepa. Journal of Agriculture and Food Chemistry 52: 5383-5390. [ Links ]
35. Peiris, K.H., J.L. Mallon & S.J. Kays (1997). Respiratory rates and vital heat of some speciality vegetables at various storage temperatures. Hort Technology 7: 46-49. [ Links ]
36. Pellejero G., M.C. Pozzo Ardizzi, G. Aschkar & P. Lui (2005). Effect of two modalities of preparation onions conserved in the field on the development of Aspergillus Niger in post-harvest. Revista Pilquén, Sección Agronomía 6: 1-7. [ Links ]
37. Pozzo Ardizzi, M.C., G. Pellejero, M.I. Gil, G. Aschkar, M.A. Abrameto, R. Roa & A. Van Konijnemburg (2008). Valoración de la severidad y la incidencia de Aspergillus Niger en dos cultivares de cebolla producidos en el valle inferior del rio Negro. ASAHO, XXXI Congreso Argentino de Horticultura, Mar del Plata. [ Links ]
38. Pozzo Ardizzi, M.C., M.A. Abrameto, G. Pellejero, G. Aschkar, M.I. Gil & A. Van Konijnemburg (2005). Efecto del período de conservación sobre algunas propiedades nutracéuticas y organolépticas en los bulbos de cultivares nacionales de cebollas (Allium cepa L.) en el valle Inferior del río Negro. RIA 34: 115-130. www.inta.gov.ar/ediciones/ria/34_3/08_POZZO [ Links ]
39. Randle, W.M. (1992). Sulfur nutrition affects nonstructural watersoluble carbohydrates in onion germplasm. Hortscience 27: 52-55. [ Links ]
40. Randle, W.M & M.L. Bussard (1993a). Pungency and sugar of short day onions as affected by sulfur nutrition. Journal of American Society for Horticultural Science 118: 766-770. [ Links ]
41. Randle, W.M. & M.L. Bussard (1993b). Streamlining onion pungency analysis. HortScience 28: 60. [ Links ]
42. Randle, W.M. (2000). Flavor Session Keynote Address. Proceding of the Third International Symposium on Edible Alliaceae. Athens, Georgia, USA. [ Links ]
43. Rodrigo, D., C. Cortes, E. Clynen, L. Schoofs, A. Van Loey & M. Kendrickx (2006). Thermal and high-pressure stability of purified polygalacturonase and pectinmethylesterase from four different tomato processing varieties. Food Research International 39: 440- 448. [ Links ]
44. Shiomi N., N. Benkeblia & S. Onodera (2005). The metabolism of the fructooligosacharides in Onion Bulbs: A Comprehensive Review. Journal Applied Glycoscience 52: 121-127. [ Links ]
45. Schwimmer, S.S. & W.J. Weston (1961). Enzymatic development of pyruvic acid in onion as measure of pungency. Journal of Agriculture and Food Chemistry 9: 301- 304. [ Links ]
46. Sinclair, P.J., A.B. Blackeney & E.W.R. Barlow (1995). Relationships between bulb dry matter content, soluble solids concentration and non structural carbohydrates composition in the onion (Allium cepa). Journal of Sciences of Food and Agriculture 69: 203-209. [ Links ]
47. Suzuki, M. & J.A. Cutcliffe (1989). Fructans in onions bulbs in relation to storage. Canadian Journal Plant Sciences 69: 1327-1333. [ Links ]
48. Terry, L.A., K.A. Law, K.J. Hipwood & P.H. Bellamy (2005). Nonstructural carbohydrates Profiles in Onion Bulbs Influence Taste Preference. Post Harvest Technology 33-40. [ Links ]
49. Uddin, M. & H.S. Mac Tavish (2003). Controlled atmosphere and regular storage induced changes in S-alk(en)yl-L-cysteine sulfoxides and allinase activity in onion bulbs Allium cepa L. cv Hysam. Post Harvest Biology Technology 28: 2 -245. [ Links ]
50. Yoo, K.S. & L.M. Pike (1999). Development of an automated system for pyruvic acid analysis in onion breeding. Science Horticulturae 82: 193-201. [ Links ]
51. Yoo, K.S. & L.M. Pike (1996). Controlled atmosphere storage suppresses leaf growth and flowering in onion bulbs. HortSciences 31: 875. [ Links ]
52 Yoo, K.S. & L.M. Pike (2000). Determination of background pyruvic acid concentration in onions, Allium species and other vegetables. Proceedings of the Third International Symposium on Edible Alliaceae.Athens, Georgia, USA. 97-99 p. [ Links ]
53. Yoo, K.S. & L.M. Pike (2001). Determination of background pyruvic acid concentration in onions Allium species and other vegetables. Science Horticulturae 89: 249-256. [ Links ]

