










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.79 no.2 Vicente López jul./dic. 2010
ARTÍCULOS ORIGINALES
Cuticle and cuticular wax development in the sunflower (Helianthus annuus L.) pericarp grown at the field under a moderate water deficit
Desarrollo de la cutícula y ceras epicuticulares en el pericarpo de girasol (Helianthus annuus L.) bajo déficit hídrico moderado en condiciones de campo
Franchini1 MC, LF Hernández1,2, LI Lindström1
1 Lab. de Morfología Vegetal. Depto. de Agronomía, UNSur. Bahía Blanca, 8000, Argentina, e-mail: franchini@uns.edu.ar ; ivlind@criba.edu.ar
2 CICPBA, La Plata, 1900, Argentina, e-mail: lhernan@criba.edu.ar
Address Correspondence to: M. Clara Franchini, Fax: +54 - 0291 - 4595127, e-mail: franchini@uns.edu.ar
Recibido - Received 5.III.2010.
Aceptado - Accepted 8.III.2010.
Abstract. Wax in the sunflower (Helianthus annuus L.) pericarp is an important morphological feature that interferes with oil quality and varies with crop management and environmental conditions. We studied the effect of a moderate water deficit (MWD) generated from early to late anthesis on quantitative development of the cuticle, and qualitative and quantitative development of the cuticular waxes (CW) of the pericarp in two hybrids grown under field conditions. The experiment was repeated during two consecutive seasons (Exp-I and Exp-II). At harvest maturity (HM), plants grown under a MWD showed higher CW content (31 to 47%) and thicker cuticles (13%) in both experiments compared to controls. Epicuticular wax (ECW) crystals showed a granular morphology. Also a gradient of CW grain accumulation was observed along the pericarp surface, being higher in the exposed upper region. We determined a reduction of CW of 29% (mg CW/g pericarp), during the development of the fruit, from reproductive stage R6 (stage when ray flowers have lost their turgidity) to HM (stage when water content of the fruit was 11%) in Exp-I. This CW reduction reached 11% from R6 to R9 (physiological maturity) in Exp-II. This was very likely caused by an erosive action on the surface of the pericap by particulate solids carried by wind or rain. These results show how internal mechanisms and external variables regulate pericarp wax content, and that fruit dehydration affects both the quantity and quality of wax formation from the time of fertilization to maturity.
Keywords: Cuticular waxes; Oil quality; Crop management; Hull; Anthesis; Harvest Maturity; Hybrid; Helianthus annuus L.
Resumen. La presencia de ceras en el pericarpo de girasol (Helianthus annuus L.) es un carácter morfológico que interfiere en la calidad del aceite y varía con las condiciones de manejo del cultivo. Se estudió el efecto de un déficit hídrico moderado (MWD) sobre el desarrollo del espesor de la cutícula y el contenido de las ceras cuticulares en el pericarpo de girasol de dos híbridos, desde la antesis temprana hasta la antesis tardía. El ensayo fue repetido durante dos años consecutivos (Experimentos I y II, respectivamente). En ambos experimentos, a la madurez de cosecha (HM), las plantas que crecieron bajo MWD presentaron mayor contenido de ceras cuticulares (31-47%) y mayor espesor de cutícula (13%) respecto de las plantas control. Los cristales de ceras epicuticulares presentaron morfología granular. Asimismo se observó sobre la superficie del pericarpo, un gradiente decreciente en la acumulación de ceras epicuticulares en sentido basípeto del fruto. Se determinó una reducción de ceras cuticulares (CW) del 29% (mg CW/g pericarpo) en el fruto desde el estadio reproductivo R6 (flores liguladas no turgentes) hasta la HM (11% de contenido de humedad en el fruto) en el Experimento I, y del 11% desde R6 a R9 (madurez fisiológica) en el Experimento II. Esta reducción fue atribuida al efecto erosivo ocasionado por las partículas transportadas por el viento y la lluvia durante la maduración del fruto. Estos resultados muestran de qué forma los mecanismos internos y las variables externas pueden regular el contenido de ceras en el pericarpo, y cómo la deshidratación del fruto afecta la calidad y cantidad de ceras desde la fertilización hasta la madurez.
Palabras clave: Ceras cuticulares; Calidad del aceite; Manejo del cultivo; Cáscara; Antesis; Madurez de cosecha; Helianthus annuus L.
INTRODUCTION
Sunflower (Helianthus annuus L.) is one of the most widely cultivated oil crops in the world (Putt, 1997) and it is considered highly adapted to a wide range of environmental conditions (Connor & Hall, 1997). However, oil quantity and quality in sunflower can change when climate variables such as temperature, solar radiation and soil water availability are limiting (Connor & Hall, 1997; Aguirrezábal et al., 2003; Lindström et al., 2007; Rondanini et al., 2007).
Crude sunflower oil can have a range of wax content from 200 to 3500 ppm (Morrison, 1983; Brevedan et al., 2000). In cold oil, waxes crystallize generating turbidity, a physical condition that affects its stability and diminishes its marketing properties (Rivarola et al., 1988; Givon & Tirtiaux, 2000). As a result, waxes have to be removed from the oil adding an extra financial cost to the overall industrial oil extraction process (Givon & Tirtiaux, 2000).
Oil waxes in sunflower are derived mainly from the fruit's hull (pericarp). Wax development occurs within and outside the pericarp's epidermis cuticle (intracuticular or ICW and epicuticular wax or ECW, respectively). From the total amount of cuticular waxes (CW), 83% derives from the pericarp (Martin & Juniper, 1970; Morrison, 1983), 16% from the teguments and 1% from the embryo (Morrison et al., 1984; Franchini, 2008).
Wax content can vary between hybrids, locations and with environmental conditions (Morrison, 1983; Morrison et al., 1984). Thermal or water stress can trigger and enhance cuticular wax synthesis in several plant organs (Martin & Juniper, 1970; Premachandra et al., 1992). The level of response is phenotypically sensitive and genetically controlled (Koornneef et al., 1989; Kerstiens, 1996, 2006; Jenks, 2002).
Although ECW morphology is consistent with the organ and botanical family (Barthloot et al., 1998), ECW amount can increase in plants subjected to mild water or heat stress conditions (Patumi et al., 2002), and the magnitude of this increment can be affected by the level of the stress applied. Even though significant water stress can diminish wax development due to the negative effect on the synthesis and storage of assimilates (Vidaver et al., 1989; Moon et al., 2003), a moderate water stress can enhance cuticle thickness and wax development (Bengston et al., 1978; Weete et al., 1978; Samdur et al., 2003).
There are no comparative studies on cuticle development, and content and morphology of ECW in dry fruits of angiosperms grown under different soil water contents (Patumi et al., 2002; Moon et al., 2003; Kerstiens, 2006). Exposure of sunflower crops to short periods of water stress, especially during the fruit filling period, reduce fruit oil content and increase pericarp proportion (Hall et al., 1985). Nevertheless, there is no information on the variability in cuticle thickness or CW content among sunflower hybrids or the effects that different environmental and agronomical practices could have on CW genesis. A more complete understanding of the ontogeny and morphology of sunflower fruit waxes would contribute to increase effectiveness of future breeding efforts aimed to produce higher quality sunflower oil.
In this paper, we have analyzed the cuticle thickness and CW content of the pericarp through various developmental stages of two sunflower hybrids grown under a transitory moderate water deficit (MWD). This MWD was generated from a reproductive stage close to first anthesis to late anthesis. Possible mechanisms of fruit dehydration from the time of fertilization to maturity are discussed.
MATERIALS AND METHODS
Plant material. Two commercial sunflower hybrids, Dekalb Dekasol (DK) 3900 and DK4030 (Monsanto, Argentina), were sown on 14 November 2003 (Exp-I), and on 8 November 2004 (Exp-II) at the experimental field of the Department of Agronomy, UNS, Bahía Blanca, Argentina (38°43'21.06" S; 62°15'45.84" W). The crop was grown under drip irrigation and managed according to recommended conventional agronomical practices (Díaz-Zorita & Duarte, 2002). Plant density was adjusted to 5.6 plants/m2. The phenology referred to here corresponds to that defined by Schneiter & Miller (1981) (Table 1).
Table 1. Major features in the development of the sunflower inflorescence in both experiments (Exp-I and Exp-II) at the reproductive stages defined by Schneiter & Miller (1981).
Tabla 1. Características principales del desarrollo de la inflorescencia de girasol en los dos experimentos (Exp-I y Exp-II) según los estadios reproductivos definidos por Schneiter y Miller (1981).
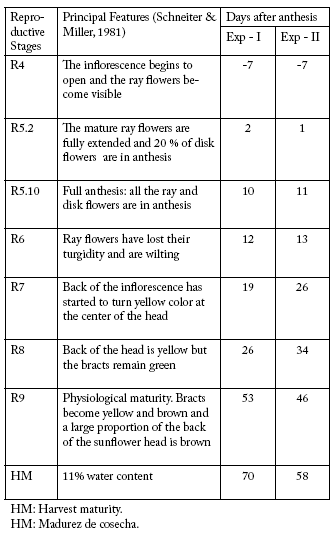
Treatments. They consisted of two water regimes: a moderate water deficit (MWD) generated by interrupting irrigation from reproductive stage R4, where the ovary wall is still developing, to R6, where the pericarp has completed its development (Lindström et al., 2007) (Table 1). Then, irrigation was restored. In the control treatment, plant water status was maintained at optimum levels during the whole growth cycle by keeping soil water content at field capacity with daily irrigation.
Plant water status in both treatments was monitored by measuring leaf relative water content (RWCleaf) (Barrs & Weatherley, 1962). With this purpose, two middle leaves from 3 plants per plot were measured at midday at different crop developmental stages. Leaf samples were placed in polyethylene bags and transported to the laboratory. Measurements were taken no longer than 60 minutes after sampling to minimize water loss due to evaporation. Moderate water deficit was determined a posteriori on the basis of the reduction of RWCleaf compared to the control (C) treatment (Bissuel-Belaygue et al., 2002).
Both experiments consisted of completely randomized split plots, with water treatment assigned to main plots and hybrids to subplots, with three replicates per treatment. Each subplot was 6.0 m long and had four rows 0.70 m apart for both experiments.
Sampling and analysis
Cuticle thickness. In both experiments, samples from 3 ovaries and/or fruits from the peripheral position of the capitulum of 3 plants per plot were taken at the reproductive stages R5.2, R6, R7, R8, R9 and Harvest Maturity (HM; fruit water content = 11%) (Table 1).
The ovaries were fixed in FAA solution (formaldehydeacetic acid-ethanol; Ruzin, 1999) and processed according to conventional histological techniques. Samples were embedded in paraffin, cut (10 μm thick), stained (saffranine + fast green) and mounted on glass slides for microscopic observation (Ruzin, 1999). Due to the high sclerification level of the pericarp, additional free-hand sections of fruits were made at the stages R6, R7, R8, R9 and HM (Table 1). These cuts were mounted in glycerin:water (1:1). Cuticle thickness was measured using an eyepiece micrometric scale.
Epicuticular wax crystal morphology. Fruits from control plants were taken from the R6 to the HM stages, and slowly dried in air using a drying chamber. Thereafter, they were mounted for scanning electron microscope (SEM) observations.
All samples were viewed on a SEM JEOL JSM-40 (JEOL, Tokyo, Japan) at 5 kV, and at a working distance of 8 mm.
Pericarp cuticular wax content. Previous work (Franchini, 2008) demonstrated that CW content in fruits of different capitulum positions is homogenous. Therefore, determination of CW content was done on fruits from the peripheral region of the capitulum. CW content was analyzed at R6, R9 and HM in both experiments (Table 1). Twenty fruits from 3 plants per plot were submerged in a previously weighed Petri dish for 2 min, using 10 ml of carbon tetrachloride (Cl4C). The solvent was then evaporated in hot plate at 40 °C. The total CW content was calculated by subtracting the initial weight of the Petri dish from its final weight and expressed by the mass of pericarp dry weight (Franchini, 2008).
Statistical analysis. Experimental results were processed by ANOVA to determine differences between treatments and hybrids, and differences between means were evaluated with the LSD test. The statistical analysis was performed with SYSTAT statistical software (SPSS, 1997). In the text, means are given in ± S.E.
RESULTS
Plant water status. At the end of the water interruption period, the RWC of MWD leaves was lower than on controls (39.64 ± 2.40 vs 42.24 ± 1.69 in Exp-I, and 51.63 ± 3.61 vs 55.48 ± 5.04 in Exp-II). The mentioned RWCleaf magnitudes showed that, in both experiments, the procedure of irrigation shortage was enough to generate a suboptimal water status during the time of fruit development. This is based on the fact that the RWC threshold to define a moderate water deficit is 12% (Bissuel-Belaygue et al., 2002).
In both experiments, plants from the MWD treatments showed a transitory wilting status. However, it was back to normal turgidity once irrigation was reestablished.
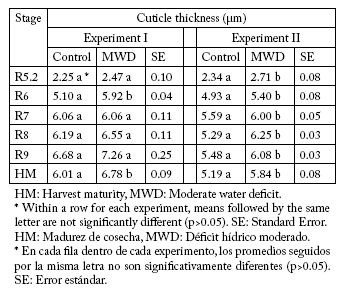
Cuticle thickness. In both experiments, the hybrid x water treatment interaction was not significant (p>0.05) for cuticle thickness at any of the developmental stages. Therefore, the average results for both hybrids were used for mean comparison between water treatments in each sampling date. There were no differences (p>0.05) between hybrids in any of the reproductive stages studied.
In Exp-I, there were significant differences (p<0.05) between water treatments in two out of the six stages analyzed (Table 2). Pericarp cuticle thickness from MWD treatment increased (p<0.05) 16% compared to the control at R6. At the same time, the observed increase (p<0.05) at HM was 13% (Table 2). In Exp-II, the MWD treatment increased (p<0.05) the thickness of the pericarp in all developmental stages by 7 to 18% (Table 2).
Table 2. Average values of cuticle thickness (μm) of sunflower pericarp (hybrids DK3900 and DK4030) at different reproductive stages (Schneiter & Miller, 1981) between both water treatments (Control and MWD) in both experiments.
Tabla 2. Valores promedio del espesor de la cutícula (μm) del pericarpo de girasol (híbridos DK3900 y DK4030) en diferentes estadios reproductivos (Schneiter & Miller, 1981) para ambos tratamientos hídricos (Control y MWD) y los dos experimentos.
Epicuticular wax crystal morphology. Since pericarp ECW crystal morphology was similar between hybrids, treatments and experiments, only the observations made on hybrid DK4030 are shown. At stage R6 (Fig. 1A) and HM (Fig. 1B-D), SEM revealed the presence of ECW crystals with granular morphology. Also, at HM there was an accumulation gradient of ECW crystals in a basipetal direction on the fruit surface (Fig. 1B-D). Thus, the presence and size of the granules was higher in the upper (Fig. 1B) than in the mid (Fig. 1C) and lower regions of the fruit (Fig. 1D). At HM, pericarps from control and MWD treatments maintained the same crystalloid granular structure and distribution of ECW crystals (Fig. 1E and F).

Fig. 1. Scanning Electron Microscopy images of the pericarp's surface of sunflower fruits (hybrid DK4030) under different treatments. (A) Epicuticular wax (ECW) crystals of granular morphology at reproductive stage R6. Lines indicate the fruit's longitudinal axis. ECW distribution of granular crystals at the upper (B), intermediate (C) and lower (D) regions of a pericarp at harvest maturity (HM). Arrows point to ECW crystals. Comparison of the accumulation and morphology of ECW crystals of the control (E) and MWD treatment (F) fruits at HM. Arrows show the contact zone between epidermal cells. (G) Pericarp's surface of a fruit at HM after removing cuticular waxes with carbon tetrachloride. Note the clean cuticle and the contact zone between epidermal cells (black arrows). A detail of the basal cell of an epidermal hair is also shown (*; Seiler, 1997). Scale bar: A-D, E, G = 20 μm. E-F = 10 μm. cut: cuticle, ECW: epicuticular wax.
Fig. 1. Imágenes con Microscopia electrónica de barrido de la superficie del pericarpo de girasol (DK4030) bajo diferentes tratamientos. (A) Morfología granular de los cristales de ceras epicuticulares (ECW) en el estadio reproductivo R6. Las líneas indican el eje longitudinal del fruto. Se observa la distribución granular de cristales en la región superior (B), intermedia (C) e inferior (D) del pericarpo a la madurez de cosecha (HM). Las flechas indican los cristales de ECW. Comparación de la acumulación y morfología de los cristales de ECW en frutos de los tratamientos control (E) y MWD (F) a la madurez de cosecha. Las flechas indican la zona de contacto entre células epidérmicas. (G) Superficie del pericarpo de un fruto a la madurez de cosecha luego de la remoción de las ceras cuticulares con tetracloruro de carbono (Cl4C). Note la cutícula limpia y la zona de contacto entre células epidérmicas (flechas negras). También se muestra un detalle de la célula basal de un pelo epidérmico (*; Seiler, 1997). Escalas: A-D, E, G = 20 μm. E-F = 10 μm. cut: cutícula, ECW: ceras epicuticulares.
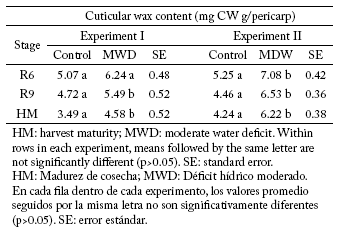
Pericarp cuticular wax content. Since the hybrid x water treatment interaction was not significant (p>0.05) for the variable CW content, only the average results for both hybrids in each experiment are presented (Table 3). Also, no differences (p>0.05) were found between hybrids in the CW content. In both experiments, pericarp CW content of MWD plants was higher than on controls (Table 3). The pericarp CW content of MWD plants increased approximately 16 to 31% in Exp-I, and 35 to 47% in Exp-II as compared to controls (Table 3).
Table 3. Mean wax cuticular content in the pericarp of two hybrids (DK3900 and DK4030) for the control and MWD treatments in both experiments at the reproductive stages R6, R9 and HM (Schneiter& Miller, 1981).
Tabla 3. Promedio del contenido de ceras cuticulares del pericarpo en los híbridos (DK3900 y DK4030) para los tratamientos control y MWD, en los estadios reproductivos R6, R9 y HM (Schneiter & Miller, 1981).
Mean water treatments values for each developmental stages (R6, R9 or HM) were compared to analyze the change in CW content per unit of pericarp dry weight (mgCW/g pericarp). Pericarp CW content decreased from R6 to HM. The observed reduction was 29% from R6 to HM in Exp-I. During Exp-II, the CW content was reduced (p<0.05) by 11% from stage R6 to R9, but not between R9 and HM (Fig. 2).
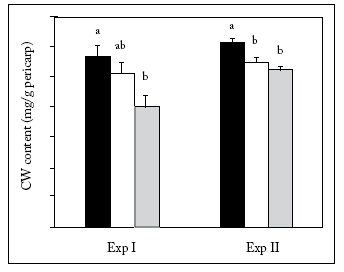
Fig. 2. Cuticular Wax (CW) content per unit of pericarp dry weight (mg CW/g pericarp), averaged over water treatments at R6 (black); R9 (white); HM (grey) in each experiment. Within each phenological stage, different letters indicate significant differences (p<0.05).
Fig. 2. Contenido de ceras cuticulares (CW) por unidad de peso seco de pericarpo (mg CW/g pericarpo) promediado de los tratamientos hídricos en los estadios R6 (negro); R9 (blanco); HM (gris) en cada experimento. Dentro de cada estadio, las barras con diferentes letras indican magnitudes significativamente diferentes (p<0.05).
DISCUSSION
It is important to relate the adaptative response of cuticle and CW development in MWD plants during fruit development to prevent unnecessary fruit dehydration that could put at risk the success of the seed development process. The epicuticular wax is the first barrier presented at the fruit-environment interface; it can protect the fruit and the physiological processes within the seed against damage produced by ultraviolet (UV) radiation (Grant et al., 2003) and microbial attack (Stephanou & Manetas, 1997). The importance of epicuticular wax in stems and leaves has led to its use in the quantification of physical attributes involved in crop improvement (Beattie & Marcell, 2002; Grant et al., 2003).
In both experiments, the MWD treatment increased cuticle thickness in comparison to the control in each of the developmental stages sampled (Table 2). We demonstrated that sunflower fruits showed an adaptive response to water deficit conditions by increasing cuticle thickness. This kind of adaptive response has been reported to occur in sugar beet leaves (Beta vulgaris L.; Luković et al., 2009) and olive fruit (O. europaea L.; Patumi et al., 2002). This indicates that water deficit applied during the ovary and fruit (stages R4-R6) developmental stages directly affects cuticle thickness.
Epicuticular wax crystal morphology. The presence of ECW crystals in the form of granules was the most prominent feature of the pericarp in the sunflower fruit from stage R6 until HM. This granular wax morphology, as defined by Barthlott et al. (1998), is characteristic of leaves from mesophytes such as Allium and Saccharum (Eglinton & Hamilton, 1967), and it has also been found in grapes (Vitis vinifera) (Casado & Heredia, 2001).
The basipetal decreasing gradient in the pericarp of the ECW crystal distribution at HM (Fig. 1B-D) can be a consequence of the way in which fruits are inserted in the receptacle. The lower region of the fruit, in close contact with its neighbors, showed less accumulation of ECW crystals and lower relief morphology than the upper region. Conversely, the upper region, which is free and exposed to the environment, and possibly more sensitive to heat and/or dehydration, showed prominent granules of ECW crystals (Fig. 1B). An irregular distribution of these granules, of a quantitative and qualitative origin, was observed in fruits from orange trees [Citrus sinensis (L.) Osbeck; El Otmani et al., 1989] and grapes (Casado & Heredia, 2001) due to different levels of solar radiation intercepted by the fruits at different positions on the plant.
Chemical composition of the waxes is influenced by climatic conditions during fruit development. In turn, chemical composition determines ECW crystal morphology (Baker, 1982; Cameron et al., 2006). SEM revealed that fruit pericarp from both water treatments showed an equally waxy appearance with similar wax morphology (Fig. 1E and F). This agrees with data reported for tobacco (Nicotiana glauca L. Graham) (Cameron et al., 2006), where the morphological shape of leaf crystals on plants exposed to periodic drying remained unchanged compared to controls. Likewise, morphology of ECW crystals was maintained in leaves and bracts of cotton (Gossypium hirsutum L.) exposed to different water conditions (Bondada et al., 1996).
Pericarp cuticular wax content. In both experiments, plants from MWD treatment increased the CW production on the pericarp when exposed to water stress (Table 3). This has been interpreted as an adaptative plant response to avoid water diffusion across the cuticle (Premachandra et al., 1992; Patumi et al., 2002). In cherry (Prunus avium L. cv. Sam; Knoche et al., 2001) and tomato (Lycopersicon esculentum L.; Vogg et al., 2004) fruits, intracuticular rather than epicuticular waxes seem to be responsible for avoiding water diffusion across the cuticle, thus acting as transpiration barriers.
Increased CW content, and reductions in cuticular transpiration rates, have been well documented in leaves of weeping lovegrass (Eragrostis curvula Schrad.; Echenique et al., 1986) and sorghum (Sorghum bicolor L.; Premachandra et al., 1992). Likewise, Cameron et al. (2006) observed a higher CW content (Nicotiana glauca L. Graham), and a subsequent reduction in epidermis conductance on the surface of tobacco leaves under water deficit. A similar response has been reported on leaves of peanut (Arachis hypogaea L.; Samdur et al., 2003) and sesame (Sesamum indicus L.; Kim et al., 2007), where water stressed plants during post-flowering had a significant increase in total wax amount.
The observed reduction in CW content and HM from R6 to R9 in both experiments (Fig. 2) could be attributed to the erosive action produced by several environmental factors, among wich rainfall and wind are particularly common (Percy& Baker, 1990). They can transport abrasive particulate material that can remove ECW crystals from the pericarp surface. The same effect has been observed on leaves of Eucalyptus sp. (Baker& Hunt, 1986) and Fragaria sp. (Neinhuis & Barthlott, 1997).
In both experiments, the highest content of CW occurred at the growth stage R6, when fruits are still young and have a high water concentration (Rondanini et al., 2007). This fact agrees with observations made by Neinhuis et al. (2001). These authors demonstrated that cuticular transpiration allows waxes attached to water molecules to move from the inner regions of the leaf to its outer surface. Thus, in young epidermis with a thin cuticle (such as that present in undeveloped fruits, with a lower resistance for the passage of waxes through it compared with mature fruits) a higher CW content can be expected.
Preliminary studies in sunflower (Franchini, unpublished), showed that the increase in thickness of the cuticle during pericarp development is accompanied by an increase in CW content. However, cuticular transpiration rates should be studied to understand how the increase in cuticle thickness and CW content affect transpiration rate. In sorghum (Jordan et al., 1984), CW contents greater than 0.067 g CW fruit/m2 provided an effective barrier to prevent water loss through the leaf cuticle under most conditions.
One might therefore make the question of how CW and the cuticle control fruit moisture loss during achene maturation. Franchini (2008) observed that a ripe fruit which had its CW removed with solvent, presented a smooth surface with no imperfections. Therefore, except for mechanical damage, there would be no other way for evaporation from the surface (Fig. 1G). The inner layers of the pericarp are known to be compressed because of seed development during fruit maturation (Lindström et al., 2007), and while dehydration is taking place (Rondanini et al., 2007). Therefore, pressure against the inner walls of the pericarp during fruit development until the vascular bundles collapse (Lindström et al., 2007) could also act as a driving force for water expulsion through the cuticle. In view of this physiological and morphological dynamics acting simultaneously during the seed filling period, and considering the important barrier to water passage across the cuticle of the pericarp, an interesting question arises: what are the most likely ways of water loss from fruit during its maturation? The sunflower pericarp can lose an average of 55 mg water in a period of 50-60 days from 10 DAA to harvest maturity (Rondanini et al., 2009). Considering an average surface of the pericarp of approximately 110 mm2 (Franchini, 2008), and a daily period of dehydration of 10 h, the fruit would lose about 0.25 mg/m.s. of water.
Extreme values of cuticular transpiration (i.e., 2.5 mg/m.s.; Jordan et al., 1984), were found in leaves of sorghum; values for the epidermis of apples varieties ranged between 0.19 and 1.84 mg H20/m.s.
Most of the outer fruit layers do not present structures associated with the physiological regulation of control of water loss, such as stomata (Ölmes et al., 2006). Therefore, water must find its way across the cuticle to evaporate. According to data reported for several organs of numerous species (Kerstiens, 2006; Schönherr, 2006), cuticular capacity to allow water movement across the wax matrix (permeance) is low, though variable. Schönherr (2006) showed that water molecules can easily find their way across the cuticle but only as individual molecules. When water transport across the cuticle is massive, the presence of aqueous pores is required (Schönherr, 2006). Pore opening depends on the content of environmental humidity. Franchini (2008) worked on the cuticle ultrastructure in achene pericarps from plants grown under optimal water conditions. She demonstrated the lack of structures similar to aqueous pores through which evaporation could occur (Schönherr, 2006). Moreover, loss of fruit water via xylem backflow to other capitulum tissues could make an important contribution to dehydration particularly under conditions of high evaporative demand (Lang, 1990). In this case, as well as during advanced maturation stages, water deficit may result in a reverse flow of water from the fruit to the plant through the fruit's vascular bundles (Layzell & LaRue, 1982). Laxman& Srivastava (2000) demonstrated the capitulum capacity for losing water through the epidermis and also via stomata located in the bracts and in the lower surface of the receptacle.
CONCLUSIONS
This work showed that MWD treatments on sunflower plants can generate changes in the quantitative levels of CW development at the fruit surface. A MWD treatment on plants during fruit development led to an increase of 23 and 42% in the CW content of the pericarp compared to controls in Exp-I and Exp-II, respectively. From R6 to HM, CW content decreased, possibly due to the erosive effect produced by wind and rain on the fruit surface.
ECW located at the upper region of the sunflower fruit can have a reflective effect, helping the fruit to avoid overheating and dehydration. It could also help to maintain the oil quantity and quality that is synthesized in the seed. So, a balance should exist between the amount of wax developed in the fruit (which can contribute to yield quality) and the industrial requirements that lead to its removal before the oil can be commercialized.
Our results can be used as a physiological tool to define the dynamics of wax accumulation in the sunflower fruit pericarp, a variable that can be genetically modified ( Jenks, 2002). Breeders need to be able to manipulate two characters currently antagonists in the sunflower fruit: seed oil and pericarp wax content. This is also the first step to the study of possible mechanisms of water loss through the pericarp, which remain unknown.
ACKNOWLEDGEMENTS
This work was supported by a grant of the Argentine Sunflower Association (ANPCyT - ASAGIR, PICTOS No. 13151) to L.F.H., the Secretaría Gral. de Ciencia y Tecnología (SeGCyT) UNS and the Comisión de Investigaciones Científicas (CIC, La Plata) Argentina. We acknowledge Dr. R.C. Johnson, Washington State University, for his valuable comments on the manuscript. We also thank Ms. A. Flemmer for collaborating in the field experiments and Ms. M. C. Moreno (CIC-PBA) for revising the manuscript. Sunflower genotypes, kindly provided by Monsanto of Argentina, are greatly appreciated.
REFERENCES
1. Aguirrezábal, L.A.N., Y. Lavaud, G.A.A. Dosio, N.G. Izquierdo, F.H. Andrade & L.M. González (2003). Intercepted Solar Radiation during Seed Filling Determines Sunflower Weight per Seed and Oil Concentration. Crop Science 43: 152-161. [ Links ]
2. Baker, E.A. (1982). Chemistry and morphology of plant epicuticular waxes. In: Cuttler, D.F., Alvin K.L. & Price C.E. (eds.), pp. 139- 165. The Plant Cuticle. Academic Press, London. [ Links ]
3. Baker, E.A. & G.M. Hunt (1986). Erosion of waxes from leaf surfaces by simulated rain. New Phytologyst 102: 161-173. [ Links ]
4. Barrs, H.D. & P.E. Weagherley (1962). A re-examination of the relative turgidity technique for estimating water deficit in leaves. Australian Journal of Biological Sciences 15: 413-428. [ Links ]
5. Barthlott, W., C. Neinhuis, D. Cutler, F. Ditsch, I. Meusel, I. Theisen& H. Wilhelmi (1998). Classification and terminology of plant epicuticular waxes. Botanical Journal of the Linnean Society 126: 237-260. [ Links ]
6. Beattie, G.A. & L.M. Marcell (2002). Effect of alterations in cuticular wax biosynthesis on the physicochemical properties and topography of maize leaf surface. Plant, Cell & Environment 25: 1-16. [ Links ]
7. Bengston, C., S. Larsson & C. Liljenberg (1978). Effects of water stress on cuticular transpiration rate and amount and composition of epicuticular wax in seedlings of six oat varieties. Physiologia Plantarum 44: 319-324. [ Links ]
8. Bissuel-belaygue, C., A.A. Cowan, A.H. Marshall & J. Wery (2002). Reproductive development of white clover (Trifolium repens L.) is not impaired by a moderate water deficit that reduces vegetative growth. I. Inflorescence, flower and ovule production. Crop Science 42: 406-414. [ Links ]
9. Bondada, B.R., D.M. Oosterhuis, J.B. Murphy & K.S. Kims (1996). Effect of water stress on the epicuticular wax composition and ultrastructure of cotton (Gossypium hirsutum L.). Environmental and Experimental Botany 36: 61-69. [ Links ]
10. Brevedan, M.I.V., A.A. Carelli & G.H. Crapiste (2000). Cambios en composición y calidad de aceites de girasol durante su extracción y desgomado. Aceites y Grasas 51: 417-423. [ Links ]
11. Cameron, K.D., M.A. Teece & L.B. Smart (2006). Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiology 140: 176-183. [ Links ]
12. Casado, C.G. & A. Heredia (2001). Ultrastructure of the cuticle during growth of the grape berry (Vitis vinifera). Physiologia Plantarum 111: 220-224. [ Links ]
13. Connor, D.J. & A. Hall (1997). Sunflower Physiology. In: Schneiter, A.A. (ed.), pp. 113-182. Sunflower Technology and Production. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI. [ Links ]
14. Díaz-zorita, M. & G.A. Duarte (2002). Manual Práctico para el Cultivo de Girasol. Hemisferio Sur, Buenos Aires. 313 p. [ Links ]
15. Echenique, C.V., M. Poverene & N.R. Curvetto (1986). Epicuticular wax and water stress in Eragrostis curvula (Schrad) Cultivars. Effects of water stress on the quantity and morphology of epicuticular wax. Microscopía Electrónica y Biología Celular 10: 91-95. [ Links ]
16. Eglinton, G. & R. Hamilton (1967). Leaf epicuticular waxes. Science 156: 1322-1335. [ Links ]
17. El Otmani, M., M.L. Arpaia, C.W. Coggings, J. Pehrson & N.V. O´Connel (1989). Developmental changes in 'Valencia' orange fruit epicuticular wax in relation to fruit position on the tree. Scientia Horticulturae 41: 69-81. [ Links ]
18. Franchini, M.C. (2008). Desarrollo de la cutícula y ceras epicuticulares del pericarpio de frutos de plantas de girasol crecidas bajo dos regímenes hídricos. MSc. Thesis. Universidad Nacional del Sur. 83 p. [In Spanish] [ Links ].
19. Givon, V. & A. Tirtiaux (2000). Revisión de los distintos métodos para la eliminación de gomas y ceras. In: Aceites y Grasas. Libro X Aniversario Recop. Art. Tec. Nº 1-41. ASAGA, pp. 101-110. [ Links ]
20. Grant, R.H., G.M. Heisler, W.Gao & M. Jenks (2003). Ultraviolet leaf reflectance of common urban trees and the prediction of reflectance from leaf surface characteristics. Agriculture and Forest Meteorology 120: 127-139. [ Links ]
21. Hall, A.J., C.A. Chimenti, F. Vilella & G. Freier (1985). Timing of water stress effects on yield components in sunflower. Proceedings of the 11th International Sunflower Conference. Mar del Plata, Argentina, p. H1-H6. [ Links ]
22. Jenks, M.A. (2002). Critical issues with the plant cuticle's function in drought tolerance. In: Wood, A.J. (ed.), pp. 97-127. Biochemical & Molecular Responses of Plants to the Environment. Research Signpost Press, Kerala, India. [ Links ]
23. Jordan, W.R., P.J. Shouse, A. Blum, F.R. Miller & R.L. Monk (1984). Environmental Physiology of Sorghum. II. Epicuticular wax load and Cuticular Transpiration. Crop Science 24: 1168-1173. [ Links ]
24. Kerstiens, G. (1996). Cuticular water permeability and its physiological significance. Journal of Experimental Botany 47: 1813-1832. [ Links ]
25. Kerstiens, G. (2006). Water transport in plant cuticle: an update. Journal of Experimental Botany 57: 2493-2499. [ Links ]
26. Kim, K.S., S.H. Park & M.A. Jenks (2007). Changes in leaf cuticular waxes of sesame (Sesamum indicum L.) plants exposed to water deficit. Journal of Plant Physiology 164: 1134-1143. [ Links ]
27. Knoche, M., S. Peschel, M. Hinz & M.J. Buckovac (2001). Studies on water transport through the sweet cherry fruit surface. II. Conductance of the cuticle in relation to fruit development. Planta 213: 927-936. [ Links ]
28. Koornneef, M., C.J. Hanhart & F.Thiel (1989). A genetic and phenotypic description of eceriferum (cer) mutants in Arabidopsis thaliana. Journal of Heredity 80: 118-122. [ Links ]
29. Lang, A.. (1990). Xylem, phloem and transpiration flows in developing apple fruits. Journal of Experimental Botany 41: 645-651. [ Links ]
30. Laxman, R.H. & G.C. Srivastava (2000). Photosynthetic and respiratory activity in sunflower (Helianthus annuus L.) bracts. Indian Journal of Plant Physiology 5: 101-104. [ Links ]
31. Layzell, D. & T. Larue (1982). Modeling C and N Transport to Developing Soybean Fruits. Plant Physiology 70: 1290-1298. [ Links ]
32. Lindström, L.I., C.N. Pellegrini & L.F. Hernández (2007). Histological development of the sunflower fruit pericarp as affected by pre- and early post-anthesis canopy shading. Field Crops Research 103: 229-238. [ Links ]
33. Luković, J., I. Maksimović, L. Zorić, N. Nagl, M. Perčić, D. Polić& M. Putnik-delić (2009). Histological characteristics of sugar beet leaves potentially linked to drought tolerance. Industrial Crops and Products 30: 281-286. [ Links ]
34. Martin, J.T. & B.E. Juniper (1970). The cuticles of plants. Edward Arnold, London. 347 p. [ Links ]
35. Moon, D., Y. Cho & F. Mizutani (2003). Wax deposition on the fruit surface of Satsuma Mandarin as affected by water stress. Asian Journal of Plant Sciences 2: 1138-1141. [ Links ]
36. Morrison III, W.H. (1983). Variation in wax content of sunflower seeds with location and hybrids. Journal of American Oil Chemistry Society 61: 1013-1015. [ Links ]
37. Morrison III, W.H., R.E. Sojka & P. Unger (1984). Effects of planting date and irrigation on wax content of sunflower-seed oil. Journal of American Oil Chemistry Society 61: 1242-1245. [ Links ]
38. Neinhuis, C. & W. Barthlott (1997). Characterization and distribution of water-repellent, self-cleaning plant surfaces. Annals of Botany 79: 667-677. [ Links ]
39. Neinhuis, C., K. Koch & W. Barthlott (2001). Movement and regeneration of epicuticular waxes through plant cuticles. Planta 213: 427-434. [ Links ]
40. Ölmes, H.A., B.E. Ak & R. Gülcan (2006). The relationship between stomata density and fruit quality of some apricot varieties growing in different altitudes in Malatya province. XII International Symposium on Apricot Culture and Decline. Acta Horticulturae 701: 111-120. [ Links ]
41. Patumi, M., R.D. Andrea, V. Marsilio, G. Fontanazza, G. Morrelli & B. Lanza (2002). Olive and olive oil quality after intensive monocone olive growing (Olea europaea L., cv. Kalamata) in different irrigation regimes. Food Chemistry 77: 27-34. [ Links ]
42. Percy, K.E. & E.A. Baker (1990). Effect of simulated acid rain on epicuticular wax production, morphology, chemical composition and on cuticular membrane thickness in two clones of Sitka spruce (Picea sitchensis (Bong) Carr). New Phytologist 116: 79-87. [ Links ]
43. Premachandra, G.S., H. Saneoka, K. Fujita & S. Ogata (1992). Leaf water relations, osmotic adjustment, cell membrane stability, epicuticular wax load and growth as affected by increasing water deficits in sorghum. Journal of Experimental Botany 43: 1569-1576. [ Links ]
44. Putt, E.D. (1997). Early History of Sunflower. In: Schneiter, A.A. (ed.), pp. 1-19. Sunflower Technology and Production. Sunflower American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI. [ Links ]
45. Rivarola, G., M.C. Añon & A. Calvelo (1988). Crystallization of waxes during sunflower seed oil refining. Journal of American Oil Chemistry Society 62: 1508-1513. [ Links ]
46. Rondanini, D.P., R. Savin & A. Hall (2007). Estimation of physiological maturity in sunflower as a function of fruit water concentration. European Journal of Agronomy 26: 295-309. [ Links ]
47. Rondanini, D.P., A. Mantese, R. Savin & A.J. Hall (2009). Water content dynamics of achene, pericarp and embryo in sunflower: Associations with achene potential size and dry-down. European Journal of Agronomy 30: 53-62. [ Links ]
48. Ruzin, S.E. (1999). Plant Microtechnique and Microscopy. Oxford University Press. New York. 322 p. [ Links ]
49. Samdur, M.Y., P. Manivel, V.K. Jain, B.M. Chikani, H.K. Gor, S. Desai & J.B. Misra (2003). Genotypic differences and Water Deficit Induced Enhancement in Epicuticular Wax Load in Peanut. Crop Science 43: 1294-1299. [ Links ]
50. Schneiter, A.A. & J.F. Miller (1981). Description of sunflower Growth Stages. Crop Science 21: 902-904. [ Links ]
51. Schönherr, J. (2006). Characterization of aqueous pores in plant cuticles and permeation of ionic solutes. Journal of Experimental Botany 57: 2471-2491. [ Links ]
52. Seiler, G.J. (1997). Anatomy and morphology of sunflower. In: Schneiter, A.A. (ed.), pp. 67-111. Sunflower Technology and Production. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI. [ Links ]
53. Stephanou, M. & Y. Manetas (1997). The effects of seasons, exposure, enhanced UV-B radiation, and water stress on leaf epicuticular and internal UV-B absorbing capacity of Cistus creticus: a mediterranean field study. Journal of Experimental Botany 48: 1977-1985. [ Links ]
54. SPSS, 1997. SYSTAT for Windows1, Release 7.0. SPSS Inc., Chicago, IL. [ Links ]
55. Vidaver, W., W. Binder, R.C. Brooke, G.R. Lister & P.M.A. Toivonen (1989). Assessment of photosynthetic activity of nursery grown Picea glauca (Moench) Voss seedlings using an integrating fluorometer to monitor variable chlorophyll fluorescence. Canadian Journal of Forest Research 19: 1478-1482. [ Links ]
56. Vogg, G., S. Fischer, J. Leide, E. Emmanuel, R. Jetter, A.A Levy & M.. Riederer (2004). Tomato fruit cuticular waxes and their effects on transpiration barrier properties: functional characterization of a mutant deficient in a very-long chain fatty acid b-ketoacyl-CoA Synthase. Journal of Experimental Botany 55: 1401-1410. [ Links ]
57. Weete, J.D., G.L. Leek, C.M. Peterson, H.E. Currie & W.D. Brance (1978). Lipid and surface wax synthesis in water stressed cotton leaves. Plant Physiology 62: 675-677. [ Links ]

