










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkPhyton (Buenos Aires)
On-line version ISSN 1851-5657
Phyton (B. Aires) vol.79 no.2 Vicente López July/Dec. 2010
ARTÍCULOS ORIGINALES
Osmotic adjustment in Leymus cinereus cv. "Trailhead" under field conditions
Ajuste osmótico en Leymus cinereus cv. "Trailhead" bajo condiciones de campo
Torres YA1, CA Busso1, OA Montenegro2, HD Giorgetti2, HD Rodríguez2, D Bentivegna3
1 Departamento de Agronomía, Universidad Nacional del Sur (UNS),CERZOS (CONICET), 8000 Bahía Blanca, Argentina.
2 Chacra Experimental Patagones, Ministerio de Asuntos Agrarios, 8504 Carmen de Patagones, Argentina.
3 CERZOS (CONICET), CCT Bahía Blanca, Camino La Carrindanga, Km. 8, 8000, Bahía Blanca, Argentina.
Address Correspondence to: Dr. Carlos A. Busso, e-mail: cebusso@criba.edu.ar
Recibido - Received 15.IX.2010.
Aceptado - Accepted 27.IX.2010.
Abstract. Osmotic adjustment has been recognized to be an important adaptive response to water stress in higher plants. Although Leymus cinereus cv. "Trailhead" is reputedly a very drought-tolerant rangeland plant, its ability to adjust osmotically in the field has not previously been reported. This study examined the occurrence or not of osmotic adjustment in field-grown plants of Leymus cinereus cv. "Trailhead" during late spring in an arid zone of Patagonia. Plants of this species had been established in the field three years before. With this purpose, leaf laminae of this species were collected and pressure-volume curves made. Results showed turgor maintenance in the face of declining leaf water potentials (i.e., osmotic adjustment) in the study species. Even more, maximum pressure at maximum leaf water potential was increased during a 3-week-period. This response is typical of plants with a previous history of osmotic adjustment as a result of water stress.
Keywords: Osmotic adjustment; Leymus cinereus cv. "Trailhead"; Water stress; Patagonia.
Resumen. El ajuste osmótico se ha reconocido como una respuesta importante de adaptación a condiciones de estrés hídrico en las plantas superiores. Aunque Leymus cinereus cv. "Trailhead" tiene la reputación de ser una especie componente de los pastizales naturales con una alta tolerancia a la sequía, su capacidad para ajustarse osmóticamente no se ha demostrado bajo condiciones de campo. Este estudio examinó la ocurrencia o no de ajuste osmótico durante la primavera en plantas de Leymus cinereus cv. "Trailhead" que crecieron bajo condiciones de campo en una zona árida de Patagonia. Las plantas fueron examinadas por esta característica adaptativa luego de su implantación en el campo tres años antes. Se recolectaron láminas foliares de esta especie y se construyeron curvas presión-volumen. Los resultados mostraron que Leymus cinereus cv. "Trailhead" mantuvo la presión de turgencia aún cuando declinó el potencial hídrico, indicando la existencia de ajuste osmótico. Más aún, la presión de turgencia máxima se incrementó cuando el potencial hídrico fue máximo luego de un período de tres semanas. Esta respuesta es típica en plantas con una historia previa de ajuste osmótico como resultado de condiciones de estrés hídrico.
Palabras clave: Ajuste osmótico; Leymus cinereus cv. "Trailhead"; Estrés hídrico; Patagonia.
INTRODUCTION
Osmotic adjustment has been recognized to be an important adaptive response to water stress in higher plants (Hsiao et al., 1976; Turner, 1979). An increase in cell solute content causing a decreased solute or osmotic potential (ΨS) is one means by which turgor pressure (P) is maintained despite a reduced total water potential (Ψ). Physiological processes (e.g., growth) can continue by maintaining turgor at otherwise inhibitory water potentials (Jones, 1992).
Active osmotic adjustment (as opposed to passive decreases in ΨS resulting from dehydration) has been demonstrated to occur in response to water stress in crop and rangeland plants (Hsiao et al., 1976; Johnson & Assay, 1993). The ability of Leymus cinereus cv. "Trailhead" to adjust osmotically in the field has not been previously reported, although this species is reputedly a very drought-tolerant rangeland plant (Granite Seed Company, 2004). In the present study we examined osmotic adjustment in field-grown Leymus cinereus cv. "Trailhead" during late spring in an arid zone of Patagonia.
MATERIALS AND METHODS
Study site. The study was conducted during November and December 2008 within an exclosure to domestic livestock in the Chacra Experimental de Patagones, Buenos Aires, Argentina (40°39'49.7" S, 62°53'6.4" W; 40 m.a.s.l.), within the Phytogeographical Province of the Monte (Cabrera, 1976). This study is part of a greater study program conducted from 2006 to 2008. Climate is temperate semiarid, with higher precipitations during the spring and fall seasons (Giorgetti et al., 2000). Several climatic parameters measured at the study site during the whole study program are shown in Figure 1. Soil is a typical Haplocalcid. Average pH is 7, and depth is not a constraint in the soil profile. The plant community is characterized by an open, shrubby stratum which includes differentquality, herbaceous species for cattle production (Giorgetti et al., 1997). Dominance of a particular grass or shrubby species in the study region is partially dependent on grazing history and fire frequency and intensity (Distel & Bóo, 1996).
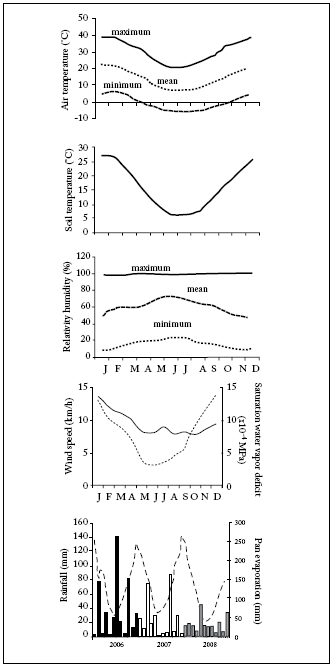
Fig. 1. Absolute monthly maximum and minimum, and mean monthly air temperatures; mean monthly soil temperatures at 0-20 cm soil depth; absolute monthly maximum and minimum, and mean monthly air relative humidity, mean monthly wind speed and saturation water vapor deficit, and mean monthly pan evaporation and monthly rainfall during 2006, 2007 and 2008 at a meteorological station located at the study site.
Fig. 1. Temperaturas del aire máximas y mínimas absolutas mensuales, y temperaturas del aire mensuales promedio; temperaturas del suelo mensuales promedio a 0-20 cm de profundidad del suelo; humedades relativas del aire máximas y mínimas absolutas mensuales, y humedades relativas del aire mensuales promedio, velocidad del viento mensual promedio y déficit de vapor de agua de saturación, y evaporación de tanque mensual promedio y lluvia mensual durante 2006, 2007 y 2008 en una estación meteorológica ubicada en el sitio de estudio.
Experimental design and defoliation treatments. At the end of 2006, 48 intraspecific plots [3 genotypes x 2 defoliation treatments x 8 plots (replicates) / treatment x 12 plants/plot = 576 plants] were established at the field. We used transplants, obtained from seeds, of P. vaginatum, and L. cinereus cvs "Magnar" and "Trailhead". Plant distance among plants in horizontal and vertical lines was 30 cm (from center to center in each plant base). In this way, competitive relationships among plants were most likely similar. Similar approaches for transplant disposition in plots have been reported in other studies (Flemmer et al., 2002). During the two study years, plants were exposed to natural rainfall (Fig. 1). Randomly chosen plants of all three genotypes were defoliated to 5 cm stubble height during the winter, plant dormancy period. In this way, only aerial plant growth produced during the following growing season was used for sampling. During mid- (22 November) to late-spring (19 December) 2006, and mid-spring 2007 (5 and 11 November), half of the plants was defoliated twice (during the vegetative and immediately after differentiation of the growth apex from vegetative to reproductive) leaving 5 cm stubble height. The other half of the plants remained non-defoliated (controls) under natural, rainfed conditions. Tillers of all three genotypes were periodically dissected and observed under a binocular microscope to determine both the developmental stage and height of the apical meristem. As a result, actively growing meristems (intercalary and leaf primordia in the growth apex) remained on the plants after the defoliation events. Despite literature reports an early internode elongation in L. cinereus (Ogle et al., 2002), this did not occur in our study. Height of the apical meristem from the tiller basal area was 5.8 ± 0.9 mm (n=15) in 2006, and 2.7 ± 0.4 mm (n=15) in 2007 on non-defoliated tillers of all three genotypes at the timing of defoliation.
Measurements
Pressure-volume curves: We only used non-defoliated plants of L. cinereous cv. "Trailhead" in this study. Youngest, completely expanded leaves (n=6; one leaf from each of different plants) were randomly selected for this study on 20 November and 12 December 2008. A pressure chamber, made to measure up to 10 MPa, was used to determine Ψ. Under the high evaporative conditions in the field it was essential to minimize post-excision evaporative losses (Brown, 1995). Each leaf was therefore wrapped in film just prior to excision, and the film was left in position during the subsequent Ψ measurements. Excised leaves were maintained within a cooler containing ice to minimize water loss and metabolic activity from the field to the laboratory. In the laboratory, pressure-volume curves were obtained as reported by Kirkam (2005) using the pressure chamber. Leaves were immersed in water overnight. Thereafter, leaves were taken out from the containers having water; water on their surface areas was dried using tissue paper, and these leaves were immediately weighted to obtain the saturated weights. The first balancing pressure was annotated at the saturating point. Then, pressure was applied incrementally to each leaf lamina. After each increase in pressure, fresh weight of the leaf lamina wrapped in film was obtained. All leaves were oven-dried at 70 °C during 72 h to obtain their individual dry weight. Relative water contents (RWC) were calculated following Kirkham (2005) for each set of saturated, dry weight and fresh weights values at each Ψ. A curve of the reciprocal of Ψ versus RWC was obtained for each leaf lamina. For each Ψ measured, pairs of Ψ and P were obtained from the pressure-volume curves as indicated by Kirkham (2005).
Leaf length measurements: Tiller leaf length (as a measure of growth) was determined monthly during the 2008 growing season. Eight randomly chosen plants were marked with this purpose. On each one of these plants, one parent tiller was permanently marked with wire cables. In these plants were determined total, dry and green leaf lengths [of blades + sheaths (green + dry)]. These measurements followed Busso and Richards (1995).
RESULTS AND DISCUSSION
There has been considerable interest in osmotic adjustment as an adaptive response to water stress in higher plants (Hsiao et al., 1976; Turner, 1979). An active increase in the concentration of cell solutes allows a positive turgor potential to be maintained at lower values of water potential than would be possible if the solute concentration were to increase solely as a result of tissue water loss.
In the present study there was evidence for active osmotic adjustment by L. cinereus cv. "Trailhead" in response to scarce spring rainfalls. This is not surprising for a cultivar of this species that can withstand 150 mm annual precipitation (Granite Seed Company, 2004). Turgor pressure was maintained when Ψ declined from the November to the December samplings (Fig. 2). This way of determining the existence of osmotic adjustment is similar to that reported by Brown (1995). Decline in Ψ was most likely the result of soil water stress. This is because of the small amount of rainfall fallen during spring 2008 [September: 4.5 mm; October: 19 mm; November: 6.5 mm and December (until the 12th): 6.5 mm].
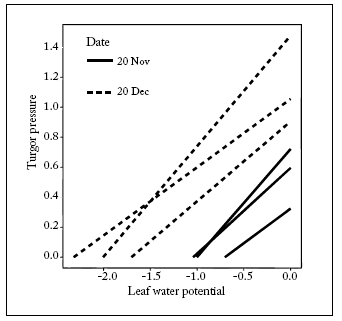
Fig. 2. Relationship between turgor pressure (MPa) and leaf water potential (MPa) in L. cinereus cv. ''Trailhead''. Each line comes from an individual leaf lamina. Values obtained from three leaf lamina had to be discarded in November and December because they broke during pressurization.
Fig. 2. Relación entre la presión de turgencia (MPa) y el potencial hídrico foliar (MPa) en L. cinereus cv. ''Trailhead''. Cada línea proviene de una lámina foliar individual. Los valores obtenidos para 3 láminas foliares en Noviembre y Diciembre tuvieron que ser descartados debido a la rotura de las láminas durante la aplicación de presión
Turgor maintenance in the face of declining Ψ most likely allowed continuation of physiological activity, and the small leaf length increases during spring of the study year (Table 1). Other studies have also reported the benefits of turgor maintenance in increasing shoot growth of various perennial rangeland grasses in the face of declining Ψ (Busso et al., 2003).
Table 1. Length (cm) of leaf lamina and sheaths (green + dry) in L. cinereus cv. ''Trailhead'' from October to December 2008. Values are the mean±1 S.E. of n=8.
Tabla 1. Longitud (cm) total de láminas y vainas (verdes + secas) en L. cinereus cv. ''Trailhead'' desde Octubre a Diciembre 2008. Los valores son el promedio±1 S.E. de n=8.

It is clear from Figure 2 that P increased at the highestΨ from the first to the second sampling date. Brown (1995) reported that an increased maximum P at maximum Ψ could be expected on the same plants during a subsequent water stress if these plants showed P maintenance because of osmotic adjustment in the previous water stress period.
REFERENCES
1. Brown, R.W. (1995). The water relations of range plants: Adaptations to water deficits. In: Bedunah, D.J., Sosebee, R.E. (Eds.), Wildland Plants: Physiological Ecology and Developmental Morphology, Soc. Range Manage., Denver, USA, pp. 291-413. [ Links ]
2. Busso, C.A., R.E. Brevedan, A.C. Flemmer & A.I. Bolletta (2003). Morphophysiological and demographic responses of perennial grasses to defoliation under water stress. In: Hemantaranjan, A. (Ed.), Plant Physiology & Plant Molecular Biology in the New Millennium. Advances in Plant Physiology, Vol. V, Scientific Publishers, Jodhpur, pp. 341-395. DOI: http://angelfire.com/ ak5/adv_pp/index.htm [ Links ]
3. Busso, C.A. & J.H. Richards (1995). Drought and clipping effects on tiller demography and growth of two bunch grasses in Utah. Journal of Arid Environments 29: 239-251. [ Links ]
4. Cabrera, A.A. (1976). Regiones fitogeográficas Argentinas. In: Ferreira Sobral, E.F. (Ed.), Enciclopedia Argentina de Agricultura y Jardinería, ACME, Buenos Aires, Argentina. (in Spanish) [ Links ]
5. .Distel, R.A. & R.M. Bóo (1996). Vegetation states and transitions in temperate semiarid rangelands of Argentina. In: West, E.N. (Ed.), Proceedings V International Rangeland Congress: Rangelands in a Sustainable Biosphere, Soc. Range Manage., Salt Lake City, USA, pp. 117-118. [ Links ]
6. Flemmer, A.C., C.A. Busso, O.A. Fernández & T. Montani (2002). Root growth, appearance and disappearance in perennial grasses: Effects of the timing of water stress with or without defoliation. Canadian Journal of Plant Science 82: 539-547. [ Links ]
7. Giorgetti, H.D., O.A. Montenegro, G.D. Rodríguez, C.A. Busso, T. Montani, M.A. Burgos, A.C. Flemmer, M.B. Toribio & S.S. Horvitz (1997). The comparative influence of past management and rainfall on range herbaceous standing crop in east-central Argentina: 14 years of observations. Journal of Arid Environments 36: 623-637. [ Links ]
8. Giorgetti, H.D., Z. Manuel, O.A. Montenegro, G.D. Rodríguez & C.A. Busso (2000). Phenology of some herbaceous and woody species in central, semiarid Argentina. Phyton, International Journal of Experimental Botany 69: 91-108. [ Links ]
9. Granite Seed Company. 2004. Granite Seed. Granite Seed Company. 89 p. [ Links ]
10. Hsiao, T.C., E. Acevedo, E. Fereres & D.W. Henderson (1976). Water stress, growth, and osmotic adjustment. Philosophical Transactions of the Royal Society, London, Series B 273: 479-500. [ Links ]
11. Johnson, D.A. & K.H. Asay (1993). Viewpoint: selection for improved drought response in cool-season grasses. Journal of Range Management 46: 194-202. [ Links ]
12. Jones, H.G. (1992). Drought and drought tolerance In Plants and microclimate. A quantitative approach to environmental plant physiology. 2nd edition. Cambridge University Press, Cambridge, U.K. pp. 272-276. [ Links ]
13. Kirkham, M.B. (2005). Principles of soil and plant water relations. Elsevier Academic Press, San Diego, California. 500 p. [ Links ]
14. Ogle, D.G., L. St. John, L. Holzworth, S.R. Winslow & T.A. Jones (2002). Basin Wildrye. NRCS Plant Guide. USDA, NRCS, Idaho State Office and the National Plant Data Center. [ Links ]
15. Turner, N.C. (1979). Drought resistance and adaptation to water deficits in crop plants. In: Stress Physiology in Crop Plants. Ed. H. Mussell and R.C. Staples, pp. 343-372. Wiley-Interscience, New York. [ Links ]

