










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkPhyton (Buenos Aires)
versão On-line ISSN 1851-5657
Phyton (B. Aires) vol.81 no.2 Vicente López jul./dez. 2012
ARTÍCULO ORIGINAL
Anatomía y micromorfología de hojas y tallos de dos cultivares de Rosa hybrida L. para flor de corte
Leaf and stem anatomy and micromorphology in two cut flower cultivars of Rosa hybrida L.
Cohen1 G, L Mascarini1, CC Xifreda2
1 Cátedra de Floricultura, Facultad de Agronomía, Universidad de Buenos Aires. Av. San Martín 4453. C.P. 1417 C.A.B.A. Argentina.
2 Laboratorio de Etnobotánica y Botánica Aplicada. FCN y Museo. UNLP. Calle 64 N° 3. 1900 La Plata. Argentina. Asociación Internacional para el Estudio de la Dinámica de Sistemas Socio-Económicos, e-mail: cxifreda@dyses.org.ar
Address Correspondence to: Graciela Silvia Cohen, e-mail: cohen@agro.uba.ar
Recibido / Received 22.IV.2011.
Aceptado / Accepted 21.VII.2011.
Resumen. En Rosa hybrida 'Lovely Red' (LR) y 'Rouge Baiser' (RB), de un cultivo comercial de Buenos Aires, Argentina, se investigaron las diferencias micromorfológicas y anatómicas de la epidermis foliar y el xilema del tallo, de la parte media inferior del folíolo terminal de hojas 5-foliadas y de la parte basal del tallo, respectivamente, en imágenes digitales obtenidas con microscopio electrónico de barrido y microscopio óptico de contraste de fase. Se observó que la cera epicuticular fue más abundante en LR que en RB y se encontró en ambas caras de la hoja. Los estomas se ubicaron sólo en la cara abaxial (111-129 estomas/mm2), y hubo entre 822 y 1025 células epidérmicas/mm2 para LR y RB, respectivamente, sin diferencias significativas entre cultivares. Se midieron 267 y 308 vasos/mm2 con una superficie total de 0,87 y 0,64 mm²/ mm² de tallo, y un área de vaso de 0,33 y 0,21 mm2 para LR y RB, respectivamente, con diferencias significativas entre cultivares para áreas de vaso y total (p=0,05). Es decir, LR presentó menor cantidad de vasos, pero más grandes y con mayor área total. Estos resultados indican que este cultivar tendría más aptitud que RB para el transporte hídrico, pero sería más susceptible al marchitamiento por falta de agua. Los resultados obtenidos podrán complementar estudios sobre cultivares de rosa de corte en relación al rol que podrían tener la epidermis foliar y los vasos xilemáticos en las diferencias en el consumo y transporte de agua, durante la vida en vaso de esos tallos. Se podrían así diagramar técnicas de manejo poscosecha que optimicen la calidad de las flores.
Palabras clave: Estomas; Vasos xilemáticos; Rosa; 'Lovely Red'; 'Rouge Baiser'.
Abstract. Micromorphological and anatomical diferences in leaf epidermis and stem xylem of Rosa hybrida 'Lovely Red ' (LR) and 'Rouge Baiser' (RB), commercially grown in Buenos Aires, Argentina, were examined in the lower half of the terminal leafet of 5-foliate leaves and in the basal part of stem, respectively. Digital images obtained from SEM and a phase contrast microscope were used with this purpose. Epicuticular wax, which was present on both sides of the leaves, was more abundant in LR than in RB. Stomata were only present on the abaxial side; they were 111 and 129 per mm2 in LR and RB, respectively; the number of epidermal cells was 1025 and 822 mm2 in LR and RB, respectively. No significant diferences were observed between cultivars. Lovely Red and RB showed 267 and 308 vessels/mm2 stem, respectively, with a total area of 0.87 and 0.64 mm²/mm² stem, respectively, and a vessel area of 0.33 and 0.21 mm2, respectively, with significant diferences between cultivars for vessel and total area (p=0.05). As a result, LR showed a smaller vessel number, but they were larger and with a greater total surface area. Terefore, this cultivar might be better than RB for water transport, although more prone to wilting under water stress. Our results complement other studies on cut rose cultivars in terms of the role of the leaf epidermis and xylem vessels in determining diferences in water consumption and transport during the stem life in glasses. Tese results might also contribute to designing post-harvest techniques for optimizing flower quality.
Keywords: Stomata; Xylem vessels; Rose; 'Lovely Red'; 'Rouge Baiser'.
INTRODUCCIÓN
La rosa (Rosa hybrida L.) es cultivada en la mayoría de los países del mundo (Lavid et al., 2002) y ocupa el primer lugar en la comercialización de flores de corte en el mercado mundial (VBN, 2009). El género Rosa L. incluye 200 especies y más de 18.000 cultivares (Haring, 1986; Gudin, 2000). En el mercado de Buenos Aires, Argentina, se citan 20 cultivares entre los más comercializados (Fernández, 2000), en una superficie de más de 30 ha (CHFBA, 2005), y se estima que estos valores son aún mayores.
Determinar el cultivar a producir es uno de los factores más importantes desde el punto de vista de su calidad poscosecha, comercialización y rentabilidad del cultivo.
Las diferencias entre cultivares son notables, tanto las morfológicas como las de comportamiento productivo, como fue demostrado por Mascarini et al. (2006) en las condiciones climáticas de Buenos Aires. Entre las primeras están el largo del tallo y la cantidad de pétalos (Cohen et al., 2010a) y entre las segundas, la apertura foral y las características de poscosecha, las que han sido descriptas en numerosos estudios (Kumar et al., 2008; Cohen et al., 2010b). Las diferencias anatómicas en varas de rosa fueron estudiadas sólo en algunos cultivares y las referencias bibliográficas son escasas. Según Put et al. (2001), la estructura de los vasos del xilema de rosa de corte cvs. 'Kardinal' y 'Sonia' no mostró diferencias esenciales. Sin embargo, en observaciones hechas en microscopio electrónico de barrido, la epidermis y la corteza del cultivar 'Kardinal' fueron ligeramente más gruesas que en cv 'Sonia'. Con relación a la anatomía de las hojas, se observó mayor índice estomático en cv 'Grand Gala', generando mayor pérdida de peso fresco en las hojas. Los tallos forales del cv 'Vega' fueron más susceptibles al marchitamiento provocado por falta de agua, lo que se relacionó con el mayor diámetro de los vasos del xilema (Hernández-Hernández et al., 2008). En rosa cv 'First Red' y en los cultivares autóctonos 'Arjun', 'Raktima', 'Raktagandha' y 'Pusa Pitamber', se encontró que en plantas formadas bajo alta temperatura, con enriquecimiento continuo de CO2, el índice estomático y la densidad de estomas y células epidérmicas aumentaron en todos los cultivares en comparación con hojas formadas bajo condiciones de temperatura óptima. Sin embargo, los cambios en la apertura de los estomas (longitud y ancho de las células oclusivas y el ostiolo) no mostraron un patrón claro en respuesta al aumento de temperatura y enriquecimiento de CO2, lo que sugiere que éste sería un carácter genotípico (Pandey et al., 2007).
El objetivo del presente trabajo fue investigar las diferencias micromorfológicas y anatómicas cualitativas y cuantitativas de la epidermis foliar y el xilema del tallo en Rosa hybrida cvs. 'Lovely Red'® y 'Rouge Baiser'®. Los resultados obtenidos complementarán estudios sobre poscosecha de cultivares de rosa de corte en relación al rol de la epidermis foliar y los vasos xilemáticos en determinar las diferencias en el consumo y transporte de agua durante la vida en vaso de tallos de rosa.
MATERIALES Y MÉTODOS
Varas florales de Rosa hybrida cvs. 'Lovely Red' (LR) y 'Rouge Baiser' (RB), provenientes de un cultivo comercial (Derqui, Buenos Aires, Argentina), fueron cosechadas en mayo 2009 sobre la segunda hoja de 5-folíolos contando desde la base, correspondientes a ramificaciones axilares caulinares con 60 días de crecimiento.
Mediciones de la micromorfología de hoja y tallo. Cortes transversales y longitudinales tangenciales y radiales de secciones de 0,5 cm de la parte basal del tallo, y secciones de la parte media inferior del folíolo terminal de hojas 5-foliadas de varas de ambos cultivares fueron observados, cuantificados y medidos sobre imágenes digitales obtenidas con microscopio electrónico de barrido (MEB) Jeol JSM-6360 LV (Facultad de Ciencias Naturales y Museo de La Plata, Argentina). Las hojas y los tallos se montaron en bases para MEB usando cinta doble faz, y fueron recubiertas con oro usando un ionizador Ion Sputter Jeol JFC-1100, para analizar la anatomía y micromorfología de tallos y hojas.
Mediciones de la anatomía foliar y caulinar. Los preparados foliares se obtuvieron rasurando en material fresco la cara abaxial del folíolo terminal de una hoja 5-foliada. Se contaron estomas y células epidérmicas de cada cultivar en imágenes digitales obtenidas con una cámara digital Sony DSC-S70 en microscopio Axioskop 2 Zeiss con óptica de contraste de fase. Se cortaron trozos de 0,5 cm de la base de las varas forales y se fjaron en 3% glutaraldehído en bufer fosfato 0,25 (pH 6,8) por 24 h, deshidratados en una serie etanol-acetona por dos horas en cada paso y embebidos en resina Spurr. Se obtuvieron secciones transversales, longitudinales tangenciales y longitudinales radiales de 1-2 µm de espesor con ultramicrótomo Sorvall Porter Blum MT2 B. Las mismas fueron teñidas con azul de toluidina (O›Brien y Mc Cully, 1981) y rojo de Rutenio (Luft, 1971) y montadas en Bálsamo de Canadá sintético marca Sigma.
Las mediciones del área de los vasos y del número de estomas se realizaron con el programa Image Tool 3.0 (Wilcox et al., 1996), y luego se calculó el área promedio de cada vaso y el área de vasos por mm2 de tallo. La densidad estomática se calculó como el número de estomas por mm2 de hoja y el índice estomático (proporción de células epidérmicas que tienen estomas) como el número de estomas/número de células epidérmicas.
El diseño estadístico fue completamente aleatorizado, con 5 repeticiones por cultivar. Los datos se analizaron con ANOVA y comparación de medias usando test de Tuckey cuando las diferencias fueron significativas con el programa InfoStat/Profesional 1.1 (Di Rienzo et al., 2002).
RESULTADOS
Evaluación de la epidermis foliar. La cera epicuticular estuvo dispuesta en partículas orientadas formando principalmente filamentos achatados ramificados; se presentó en las caras adaxial y abaxial de las hojas y fue más abundante en LR que en RB (Fig. 1A y B). La epidermis de ambos cultivares careció de tricomas y papilas, y estuvo compuesta por una sola capa de células epidérmicas. Estas células, vistas en superficie con MEB, mostraron las paredes anticlinales levemente hundidas y no fueron fácilmente visualizadas por su abundante cantidad de cera. Las células epidérmicas vistas en superficie en el microscopio óptico fueron alargadas, dispuestas irregularmente, y las paredes anticlinales fueron muy sinuosas. Los estomas tuvieron distribución hipostomática (Fig. 1C y D); sólo estuvieron sobre las áreas del mesófilo clorenquimatoso y fueron inexistentes sobre las nervaduras primarias y secundarias que sobresalieron abaxialmente.
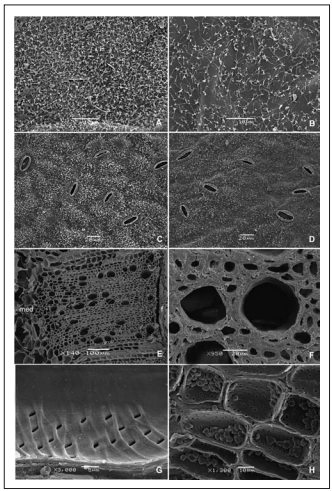
Fig. 1. A-D: Micromorfología de la epidermis del folíolo terminal de una hoja 5-foliada de rosa 'Lovely Red' (LR) y 'Rouge Baiser' (RB). A(LR)-B(RB): Cera epicuticular en la cara adaxial; C(LR)-D(RB): Vista superficial de la cara abaxial mostrando la distribución de los estomas. E-H: Micromorfología del xilema del tallo floral de rosa 'Lovely Red' (LR) y 'Rouge Baiser' (RB). E(RB): Corte transversal mostrando la distribución de los vasos, Tr; F(LR): Detalle del xilema Tr; G(LR): Engrosamientos espiralados en elementos de vasos, Lg Tg; H(LR): Parénquima radial con granos de almidón compuestos, Tr.
Abreviaturas: m: médula; f: floema; Tr: corte transversal; Lg Tg: corte longitudinal tangencial.
Fig. 1. A-D: Micromorphology of epidermis in the terminal leafet of a 5-foliate leaf in rose 'Lovely Red' (LR) and 'Rouge Baiser' (RB). A(LR)-B(RB): Epicuticular wax in the adaxial leafet side; C(LR)-D(RB): Stomata distribution in abaxial side. E-H: Xylem micromorphology in the floral stem of rose 'Lovely Red' (LR) and 'Rouge Baiser' (RB). E(RB): Vessels distribution, Tr; F(LR): Xylem detail, Tr; G(LR): Helical sculpture in vessels members, Lg Tg; H(LR): Radial parenchyma with compound starch grains, Tr.
Abbreviations: m: pith; f: phloem; Tr: transversal cut; Lg Tg: longitudinal tangential section.
El número de estomas fue de 111 y 129 por mm2, y el de células epidérmicas de 822 y 1025 por mm2 para LR y RB, respectivamente, y las diferencias no fueron significativas entre cultivares (Tabla 1).
Tabla 1. Datos cuantitativos de la epidermis foliar (densidad estomática, cantidad de células epidérmicas e índice estomático) y del xilema del tallo (área de los vasos por mm2 de tallo, área del vaso y cantidad de vasos) de rosas 'Lovely Red' (LR) y 'Rouge Baiser'(RB). Letras diferentes indican diferencias estadísticamente significativas (p=0,05).
Table 1. Quantitative data in the foliar epidermis (stomatal density, number of epidermal cells and stomatal index) and stem xylem (vessel area/mm2 of stem, vessel area and number of vessels) of roses 'Lovely Red' (LR) and 'Rouge Baiser' (RB). Different letters indicate significant differences (p=0.05).

Evaluación de la estructura del tallo. Los tallos estudiados de ambos cultivares mostraron crecimiento secundario, y en sección transversal tuvieron forma circular. Estos no mostraron presencia de exudados con materiales bloqueantes tales como látex, mucílagos y gomas, ni de taninos, suberina o materiales amorfos. Presentaron epidermis uniestratificada, corteza parenquimática (Fig. 2A), cilindro vascular completo de estructura colateral y médula parenquimática compacta (Fig. 2B). La epidermis estuvo formada por células epidérmicas propiamente dichas y células estomáticas, careció de tricomas y presentó una cutícula gruesa. La corteza estuvo formada por 12-14 capas de células parenquimáticas con engrosamientos diferenciales; las 5-6 capas externas presentaron paredes lignificadas engrosadas, mientras que las células internas de la corteza presentaron paredes delgadas. No se observó colénquima. El foema secundario presentó casquetes de fbras foemáticas asociadas (Fig. 2A).

Fig. 2. Anatomía del xilema del tallo floral de rosa 'Lovely Red' (LR) y 'Rouge Baiser' (RB). A(LR): Estructura del tallo, Tr X100. B(RB): Corteza del tallo, Tr X400. C(LR)-D(RB): Detalle del xilema, Tr X400. E(LR): Corte longitudinal tangencial mostrando los radios parenquimáticos uniseriados y multiseriados, Lg Tg X400. F(RB): Estructura del tallo, Lg Tg X100.
Abreviaturas: co: corteza; cu: cutícula; med: médula; ru: radio uniseriado; rm: radio multiseriado; Tr: corte transversal; Lg Tg: corte longitudinal tangencial.
Fig. 2. Xylem anatomy of the floral stem in rose 'Lovely Red' (LR) and 'Rouge Baiser' (RB). A(LR): Stem structure, Tr x100. B(RB): Stem cortex, Tr X400. C(LR)-D(RB): Xylem composition, Tr X400. E(LR): Uni- and multiseriate parenchyma rays, Lg Tg. X400. F(RB): Stem structure, Lg Tg, X100.
Abbreviations: co: cortex; med: pith; ru: uniseriate ray; rm: multiseriate ray; Tr: trasnversal cut; Lg Tg: longitudinal tangential section.
La estructura del xilema en sección transversal, mostró que los vasos fueron numerosos, redondeados a angulares, y midieron entre 25 y 52 µm (con 30 µm de diámetro promedio) en RB, y entre 30 y 55 µm (con 35 µm de diámetro promedio) en LR. Los vasos estuvieron distribuidos más o menos uniformemente en todo el xilema y fueron generalmente solitarios. En RB, mientras la mayor parte fueron solitarios, hubo escasos vasos tangenciales cortos agrupados de a dos o tres. LR presentó vasos numerosos, de mayor tamaño que RB pero en menor cantidad (Fig. 2C).
En RB los vasos también fueron numerosos pero disminuyó su cantidad hacia el interior del tallo (Fig.2D). En sección longitudinal se observó que los vasos presentaron perforaciones simples con placas de perforación oblicuas, y la disposición de las punteaduras areoladas fue alterna. Las paredes de los vasos presentaron engrosamientos espiralados delgados (Fig.1G).
El parénquima asociado a vasos fue escaso, de tipo paratraqueal y apotraqueal difuso. Se presentaron radios parenquimáticos multiseriados y escasos uniseriados (Fig. 2E). La distribución del parénquima radial formó radios uniseriados escasos y multiseriados 4-6-seriados de hasta 5 mm de alto y, en RB, se distinguieron también radios 3-seriados. Los radios uniseriados fueron predominantemente homocelulares formados por células erectas; los multiseriados fueron heterocelulares formados por células erectas, cuadradas y procumbentes (Fig. 2E) y presentaron abundante almidón compuesto (Fig. 1H). Los elementos celulares más comunes fueron las fibras, de 9 a 12 µm de ancho, de paredes muy engrosadas de 2-4 µm de espesor con punteaduras simples. Sólo en RB se observó una estructura parcialmente estratificada en la disposición de las fibras (Fig. 2F).
El número de vasos fue de 267 y 308 vasos por mm2 para LR y RB, respectivamente. La superficie total de estos vasos fue de 0,87 y 0,64 mm²/mm² de tallo, y los vasos tuvieron un área de 0,33 y 0,21 mm2 para LR y RB, respectivamente. Hubo diferencias significativas entre cultivares para áreas de vaso y total (p=0,05) (Tabla 1).
DISCUSIÓN
Las características anatómicas observadas en este estudio concuerdan con las detalladas para el género Rosa L. (Fahn et al., 1986) y la familia Rosaceae (Metcalfe y Chalk, 1979). La presencia de abundante cera en la cara abaxial de las hojas ya fue citada para la Rosa híbrida de té por Martin y Juniper (1970). La cantidad de cera epicuticular es importante en la reducción de la humectabilidad de las superficies y afecta la permeabilidad cuticular (Fahn, 1990). Además, cubre los órganos aéreos de las plantas lo que contribuye a prevenir la pérdida descontrolada de agua (Kosma y Jenks, 2007). Mientras la estructura de los depósitos de cera fue igual en los dos cultivares estudiados, una mayor cantidad de cera depositada, tal como muestra LR, podría incidir en la tasa de transpiración de las hojas y el consumo de agua. Por otro lado, el reflejo de la luz por la cera cuticular protege a la hoja del daño por calor (McClendon, 1984).
La micromorfología cristalina que presentaron ambas epidermis fue clasificada como hebras por Barthlott et al. (1998). Estos autores afirman que la formación del tipo cristaloide está determinada por el componente químico dominante. Las hebras estarían constituidas mayormente de flavonoides, cuyas propiedades fotoquímicas sugieren que pueden funcionar como protectores UV (Yu et al., 2006). Bruun (2005) citó la presencia de flavonoides en hojas de Rosa rugosa Murray. La hoja hipostomática, con estomas anomocíticos confinados a la cara abaxial, se corresponde con caracteres mesofíticos, propios de plantas de climas templados (Cutler, 1978). Nuestros resultados coinciden con las observaciones realizadas por Pandey et al. (2007) y da Costa Ferreira et al. (2009) en Rosa hybrida.
La presencia de una cutícula gruesa es una característica típica en las paredes externas de los tallos de rosa (Martín y Juniper 1970). Hernández-Hernández et al. (2009) lo confirmaron en los cultivares 'Gran Gala' y 'Vega'. Nuestro trabajo confirma la presencia de dicha cutícula para los cvs. 'Lovely Red' y 'Rouge Baiser'.
La presencia de vasos como únicos elementos conductores del xilema, por la carencia de traqueidas, fue corroborada como un carácter diagnóstico de valor genérico. Fahn et al. (1986) tampoco describieron traqueidas cuando estudiaron la anatomía del leño en cuatro especies de rosa. Bruun (2005) obtuvo mediciones del diámetro de vasos (35 µm), y del diámetro y espesor (11 y 2 µm, respectivamente) de los elementos celulares, concordantes con las de la presente investigación. Mientras nosotros hemos reconocido a estos elementos como fibras xilemáticas, el citado autor los denominó fibrotraqueidas. La presencia de engrosamientos espiralados en los elementos vasales tiene significado funcional pues contribuiría a aumentar la superficie de la pared, lo cual incrementaría la superficie de contacto con el agua (Carlquist, 1988).
La superficie de los cortes transversales de tallos de rosa observados microscópicamente no mostraron presencia de (1) exudados con materiales bloqueantes (látex, mucílago, gomas), (2) taninos, (3) suberina o (4) materiales amorfos. Estos resultados coinciden con lo informado por Van Door (1995). Las diferencias anatómicas del tallo que permitirían separar a los dos cultivares estudiados son principalmente cuantitativas. La distribución de los vasos en el anillo de crecimiento fue más homogénea en LR, que presentó menos vasos, de mayor diámetro; en cambio, en RB hubo más vasos, los que disminuyeron hacia el interior del leño y fueron de menor diámetro. Cuanto mayor es el diámetro de los vasos, mayor es la conductividad del agua (Carlquist, 1988). Desde que los vasos son estructural y funcionalmente similares a tubos capilares, su conductancia hidráulica se incrementa con la cuarta potencia de su radio (ver ecuación de Hagen y Poiseuille: Niklas, 1992). Sin embargo, los vasos de mayor tamaño son los que cavitan primero en almacenamiento de tallos en seco (Salleo y Lo Gullo, 1989), y la cavitación reduce considerablemente la conductividad hidráulica (Van Door, 1995); esto fue demostrado en rosas cortadas (Dixon et al., 1988). Siendo que el área total de vasos por unidad de superficie de tallo es mayor en LR, este cultivar podría tener más aptitud que RB para el transporte hídrico. Esta conclusión se ve reforzada porque los vasos del xilema son los elementos fundamentales para la conducción de agua por la falta de traqueidas. Sin embargo, los vasos de conducción en LR serían más susceptibles al marchitamiento por falta de agua; esto fue demostrado en rosa 'Vega' (Hernández-Hernández, 2009).
A la luz de los resultados obtenidos en el presente trabajo, nuevas investigaciones sobre la vida poscosecha de los cultivares analizados podrían permitir relacionar las características anatómicas y micromorfológicas descriptas con el consumo de agua y vida en vaso y, con ello, diagramar estrategias de manejo poscosecha de los mismos.
AGRADECIMIENTOS
Agradecemos a la Dra. S. B. Maldonado (FCEN, UBA), por la infraestructura otorgada en el uso del Laboratorio; al Lic. R. Urréjola (FCN y Museo, UNLP) por la asistencia técnica en las sesiones MEB; al Ing. Agr. G. Lorenzo (Cátedra de Floricultura, FAUBA) por sus aportes en metodología estadística, y al cultivo Rosas Argentinas por proveer el material vegetal para la investigación.
El subsidio CIC-PBA y el proyecto UBACyT G424 financiaron la realización del presente trabajo.
REFERENCIAS
1. Barthlott, W., C. Neinhuis, D. Cutler, F. Ditsch, I. Meusel, I. Teisen y H. Wilhelmi (1998). Classification and terminology of plant epicuticular wax. Botanical Journal of the Linnean Society 126: 237-260. [ Links ]
2. Bruun, H.H. (2005). Rosa rugosa Tunb. ex Murray. Journal of Ecology 93: 441-470. [ Links ]
3. Carlquist, S. (1988). Comparative Wood Anatomy. Springer-Verlag (eds), Berlin. 456 p. [ Links ]
4. CHFBA. (2005). Censo Hortiforícola Provincia de Buenos Aires. Dirección Provincial de Estadística (DPE) y Dirección de Economía Rural (DER) Gobierno de la Provincia de Buenos Aires. 115 p. [ Links ]
5. Cohen, G., L. Mascarini, G. Lorenzo, C.C. Xifreda y F. Vilella (2010a). Consumo de agua, área foliar y de pétalos de dos cultivares de rosa y su relación con la vida poscosecha. XXXIII Congreso Argentino de Horticultura, 28 Septiembre-1 Octubre, Rosario, Argentina. Actas, 69 p. [ Links ]
6. Cohen, G., L. Mascarini, G. Lorenzo y C.C. Xifreda (2010b). Vida poscosecha de dos cultivares de Rosa hybrida con el uso de germicidas en el agua del vaso. 5º Congreso Argentino de Floricultura y Plantas ornamentales- 12ª Jornadas Nacionales de Floricultura, 2-5 Noviembre, Entre Ríos, Argentina. Actas, pp. 52-55. [ Links ]
7. Cutler, D.F. (1978). Applied plant taxonomy. Longman, London. 103 p. [ Links ]
8. da Costa Ferreira, C., K.R. Sousa de Oliveira, P.L. Ferreira Borges, V.F. Mainardi y N.C. Barboza da Silva (2009). Anatomia foliar e caulinar de Rosa spp (L.), Rosaceae. XIII Encontro Latino Americano de IniciaÇão Científca e IX Encontro Latino Americano de Pós-GraduaÇão - Universidade do Vale do Paraíba, Brasil. [ Links ]
9. Di Rienzo, J., M. Balzarini y C.W. Robledo (2002). InfoStat/Profesional. Versión 1.1. Universidad Nacional de Córdoba. Estadística y Diseño. FCA, Argentina. [ Links ]
10. Dixon, M.A., J.A. Butt, B.P. Murr y M.J. Tsujita (1988). Water relation of cut greenhouse roses: the relationship between stem water potential, hydraulic conductance and cavitation. Scientia Horticulturae 36: 109-118. [ Links ]
11. Fahn, A., E. Werker y P. Baas (1986). Wood Anatomy and Identification of Trees and Shrubs from Israel and Adjacent Regions, The Israel Academy of Sciences and Humanities, Jerusalem. 221 p + 82 plates. [ Links ]
12. Fahn, A. (1990). Plant Anatomy. 4th ed., Pergamon Press. Oxford. 588 p. [ Links ]
13. Fernández, H.L. (2000). El cultivo de la rosa, para flor cortada. 2da Parte. Boletín técnico EEA INTA Escobar. http://www.inta.gov.ar/sanpedro/info/doc/ orn/hf_001.htm [ Links ]
14. Gudin, S. (2000). Rose: genetics and breeding. Plant Breeding 17: 159-189. [ Links ]
15. Haring, P.A. (1986). Modern Roses. American Rose Society, Shreveport, LA. 129 p. [ Links ]
16. Hernández-Hernández, F., M. L. Arévalo-Galarza, M. T. Colinas-León, H. A. Zavaleta- Mancera y J. Valdes Carrasco (2009). Diferencias anatómicas y uso de pulso en dos cultivares de Rosa (Rosa sp.). Revista Chapingo. Serie Horticultura. México. Vol. 15-2: 11-16. [ Links ]
17. Kosma, D.K. y M.A. Jenks (2007). Eco-physiological and moleculargenetic determinants of plants cuticle function in drought and salt stress tolerance. En: Advances in Molecular breeding toward drought and salt tolerant crops. M.A. Jenks, P.M. Hasegawa, y S.M. Jain (eds.). Springer, Dordrecht, The Netherlands. pp. 91-120. [ Links ]
18. Kumar, N., G.Ch. Srivastava y K. Dixit (2008). Flower bud opening and senescence in roses (Rosa hybrida L.). Plant Growth Regulation Springer 55: 81-99. [ Links ]
19. Lavid, N., J. Wang, M. Shalit, I. Guterman, E. Bar, T. Beuerle, N. Menda, S. Shafir, D. Zamir, Z. Adam, A. Vainstein, D. Weiss, E. Pichersky y E. Lewinsohn (2002). O-Methyltransferases Involved in the Biosynthesis of Volatile Phenolic Derivatives in Rose Petals. Plant Physiology Vol. 129: 1899-1907. [ Links ]
20. Luft, J. (1971). Ruthenium red and violet. II. Fine structural localization in animal tissues. Anatomical Record 171: 369- 392. [ Links ]
21. Martin, J.T. y B.E. Juniper (1970). The cuticles of plants. E. Arnold, London. 347 p. [ Links ]
22. Mascarini, L., G.A. Lorenzo y F. Vilella (2006). LAI, water content and R: FR ratio calculated by spectral refectance and its relation with plant arquitecture and cut rose production. The Journal of the American Society for Horticultural Science 131: 313-319. [ Links ]
23. McClendon, J. H. (1984). The micro-optics of leaves. I Patterns of refection from the epidermis. American Journal of Botany 71: 1391-97. [ Links ]
24. Metcalfe, C.R. y L. Chalk (1979). Anatomy of the Dicotyledons. 2nd ed.; Vol. I Clarendon Press, Oxford. 297 p. [ Links ]
25. Niklas, K.J. (1992). Plant biomechanics: an engineering approach to plant form and function. Chapter four: The ascent of water. Chicago, IL, USA: University of Chicago Press. 622 p. [ Links ]
26. O' Brien, T. y M. McCully (1981). The Study of Plant Structure, Principles and Selected Methods. 6: Formulary pp. 1-104. Termarcarphi Pty, Melbourne. [ Links ]
27. Pandey, R., M. Priya Chacko, M.L. Choudhary, K.V. Prasad y P. Madan (2007). Higher than optimum temperature under CO2 enrichment influences stomata anatomical characters in rose (Rosa hybrida). Scientia Horticulturae 113: 74-81. [ Links ]
28. Put, H. M.C. y A.C.M. Clerkx (2001). Anatomy of cut rosa xylema observed by scanning electron microscope. Proc. III IS Rose Research. Acta Horticulturae 547: 331-9. [ Links ]
29. Salleo, S. y M.A. Lo Gullo (1989). Xylem cavitation in nodes and internodes of Vitis vinifera L. plants subjected to water stress: limits of restoration of water conduction in cavitated xylem conduits. En: Structural and functional responses to environmental stresses: Water Storage. K.H. Kreeb, H. Richter y T.M. Hinckley (eds.). SPB Academic Publishing, The Hague, The Netherlands. pp. 33-42. [ Links ]
30. Van Doorn, W.G. (1995). Vascular occlusion in cut rose flowers: a survey. Acta Horticulturae 405: 58-66. [ Links ]
31. VBN. Federation of Dutch Flower Auctions. (2009). http://www.vbn.de/ [ Links ]
32. Wilcox, C., S. Dove, W. McDavid y D. Greer (1996). Image Tool, Version 1.0. Department of Dental Diagnostics at the University of Texas Health Science Center, San Antonio, TX. [ Links ]
33. Yu, O., M. Matsuno y S. Subramanian (2006). Flavonoid Compounds in Flowers: Genetics and Biochemistry. In: Floriculture, Ornamental and Plant Biotechnology Volume I. Global Science Books, UK. pp. 283-292. [ Links ]

