










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.81 no.2 Vicente López jul./dic. 2012
ARTÍCULO ORIGINAL
Hábito de crecimiento y anatomía de la raíz en Macroptilium erythroloma y Macroptilium bracteatum (Fabaceae)
Growth habit and root anatomy in Macroptilium erythroloma and Macroptilium bracteatum (Fabaceae)
Martín Montiel DC1, SM Pérez de Bianchi1, A Etcheverry1, C Camardelli2, M Alemán1, T Figueroa1
1 Cátedra Botánica General. Facultad de Ciencias Naturales. Consejo de Investigación. Universidad Nacional de Salta.
2 Cátedra Zootecnia Especial. Facultad de Ciencias Naturales. Consejo de Investigación. Universidad Nacional de Salta. Avenida Bolivia 5150. 4400 Salta, Argentina.
Address Correspondence to: Dra. Dinca Cristina Martín Montiel, e-mail: dincamartin@gmail.com; Tel.: +54 387 4255434, Fax: +54 387 4360163. Av. Bolivia 5150. 4400 Salta, Argentina. Cátedra Botánica General.
Recibido / Received 13.IX.2011.
Aceptado / Accepted 14.I.2012.
Resumen. Se estudió el hábito de crecimiento y la anatomía de la raíz en dos especies del género Macroptilium, Fabaceae, nativas de las provincias del Noroeste argentino, y potencialmente forrajeras. El objetivo del trabajo fue determinar la capacidad de supervivencia de las especies en estudio, relacionando el hábito de crecimiento y la persistencia con aspectos morfológicos y anatómicos. Se colectaron muestras de raíz y se practicaron cortes con micrótomo y a mano alzada a diferentes distancias del ápice. Los cortes se examinaron con microscopios óptico y electrónico de barrido usando diversas técnicas de tinción. Macroptilium bracteatum y M. erythroloma son plantas perennes con raíces suculentas reservantes, donde la parte aérea muere en invierno y el rebrote ocurre en la primavera desde el área de innovación a través de yemas sub-superficiales. Además, M. bracteatum presenta raíces gemíferas con yemas adicionales, constituyendo una importante estrategia para el rebrote. Estos resultados indican que ambas especies presentan potencialidad forrajera.
Palabras clave: Macroptilium; Anatomía de la raíz; Raíces gemíferas; Rebrote de raíces; Especies forrajeras.
Abstract. We studied the growth habit and root anatomy on two species of the Macroptilium genera, Fabaceae. These species, potential forages, are native to the Northwest Provinces in Argentina. The aims of this paper were to determine the survival capacity of the study species relating its growth habit and persistence with morphological and anatomical features. Root samples were collected and cut using either a microtome or by hand, at varying distances from the root tip. Cuts were examined under a light microscope, and with scanning electron microscopy (SEM) using staining techniques. Macroptilium bracteatum and M. erythroloma are perennial herbs. Aerial parts die during winter, and sprouting occurs in spring from the innovation area, throughout sub-superficial buds. In addition, M. bracteatum and M. erythroloma develop root succulence as a strategy of persistence. Besides, M. bracteatum presents endogenous root buds arising from one-year-old roots with secondary growth. These results indicate that both Macroptilium bracteatum and M. erythroloma have good potential as forage species.
Keywords: Macroptilium; Root anatomy; Root buds; Root sprouts; Forage species.
INTRODUCCIÓN
Macroptilium (Fabaceae, Papilionoideae) es un género con dieciocho especies americanas, de las cuales nueve crecen en Argentina (Drewes, 1997). Entre ellas Macroptilium bracteatum (Nees y C. Mart.) Maréchal y Baudet y Macroptilium erythroloma (Mart. Ex Benth.) Urban, abundan en la región del noroeste argentino. Crecen en climas con una marcada estación seca (invierno-primavera) y veranos lluviosos con un rango de precipitación entre 500 y 1900 mm por año e inviernos con temperaturas bajo cero. Las condiciones hídricas preferidas por estas especies varían desde climas semiáridos a secos y sub-húmedos a húmedos, habitando en lugares con precipitaciones anuales de 500 mm. Macroptilium bracteatum se encuentra en suelos con textura media y disturbados por actividad antrópica, y M. erythroloma se desarrolla en suelos arenosos a lo largo de ambientes riparios (Pérez et al., 1999a).
Estudios previos sobre el valor nutricional de algunas especies de Macroptilium (Camardelli et al., 1996; Pérez de Bianchi et al., 1999b) revelaron que las mismas tienen un alto contenido de proteínas, constituyendo una buena alternativa forrajera para el área. Por otra parte, observaciones personales a campo, permitieron registrar el consumo por parte del ganado.
En las especies forrajeras, la persistencia a lo largo de los años es un valor que las califica positivamente. El objetivo del presente trabajo fue determinar la capacidad de perpetuación de estas especies, relacionando el hábito de crecimiento y la persistencia con aspectos morfológicos y anatómicos.
MATERIALES Y MÉTODOS
Este estudio se llevó a cabo en la provincia de Salta, noroeste de Argentina. Para cada especie de Macroptilium se marcaron 20 plantas en poblaciones naturales. Por otra parte, se trasplantaron 20-25 plantas de cada especie a condiciones de invernadero y campos experimentales de la Estación Experimental INTA - Salta y de la Universidad Nacional de Salta, respectivamente. Ambos grupos fueron monitoreados cada dos semanas durante un lapso de cuatro años para registrar la dinámica de crecimiento y el ciclo de vida. En cada observación se registró presencia y actividad de yemas y desarrollo de ramificaciones.
Para el análisis anatómico comparativo de raíz en crecimiento primario, se coleccionaron 100 semillas de cada especie de poblaciones naturales, las que se sembraron en cajas de Petri en laboratorio bajo condiciones de luz y temperatura naturales. Dos semanas posteriores a la germinación fueron seleccionadas al azar 20 plántulas de cada especie y se practicaron cortes transversales a mano alzada a distintas distancias del ápice. Para comparar la anatomía de raíces en crecimiento secundario de un año de M. bracteatum y M. erythroloma, se colectaron muestras de raíces de plantas que crecieron en invernadero, se fijaron en FAA y el material fue procesado con técnicas histológicas convencionales (D'Ambrogio, 1986). Se realizaron cortes transversales en raíces principales, y se efectuó la tinción utilizando safranina-fast green para confeccionar preparados permanentes. Las raíces de almacenamiento de M. bracteatum y M. erythroloma fueron analizadas en secciones transversales de 1,5 a 2 mm, y de 4 a 6 mm. La presencia de amiloplastos fue determinada por medio de microscopio con luz polarizada.
Para el estudio de raíces gemíferas de M. bracteatum, las muestras fueron lavadas y examinadas bajo lupa para analizar la presencia de protuberancias que indican la posición de las yemas. Se obtuvieron fotografías con microscopio Zeiss, usando película BanW Professional Kodak 400 CN.
Cuando se usó el microscopio electrónico de barrido (SEM), se realizaron secciones transversales que fueron deshidratadas en series graduales de alcoholes y llevadas a punto crítico. Se montaron en platinas y se metalizaron con oro. Los preparados fueron observados usando microscopio electrónico de barrido JEOL Modelo JSM 6480 LV de LASEM (Laboratory of Scanning Electron Microscopy) - ANPCyT - UNSa - CONICET - Salta, Argentina. Los ejemplares fueron depositados en el Herbario del Museo de Ciencias Naturales de Salta, de la Universidad Nacional de Salta (MCNS).
RESULTADOS
Morfología del rebrote. Macroptilium bracteatum y M. erythroloma son hierbas perennes que en el primer año formaron una corona a partir de las tríades de yemas del nudo cotiledonar y de las hojas primordiales; dichas tríades se organizaron con una yema axilar y dos yemas profilares (Debouck, 1984). A partir de las yemas de estos nudos, brotaron ramas de primer y segundo orden con entrenudos basales cortos, que a su vez portaron yemas dispuestas en tríades otorgándole a estas plantas una marcada basitonía, mientras se produjo la acumulación de reservas en las raíces. En invierno, la parte aérea murió y en la primavera siguiente los rebrotes se produjeron a partir del complejo axilar, tanto de los nudos basales de las ramas de primer orden como de las de segundo orden de la corona, utilizando las reservas almacenadas en las raíces. Estas características le brindan la característica de plurianual.
Anatomía de raíz de Macroptilium bracteatum y M. erythroloma en crecimiento primario. En las secciones transversales de la raíz principal de las plántulas analizadas, los tejidos estuvieron totalmente diferenciados. En ambas especies la epidermis fue uniestratificada con pelos absorbentes y la exodermis presentó dos estratos indiferenciados de células; la corteza estuvo constituida por 5 - 7 estratos de células parenquimáticas con pequeños espacios intercelulares y endodermis con Bandas de Caspary. El periciclo presentó un estrato de células y la disposición de los tejidos de conducción indicó que la raíz es tetrarca.
Anatomía de raíz en crecimiento secundario en Macroptilium bracteatum y M. erythroloma. El análisis de los transcortes practicados sobre raíces secundarias de 1,5 a 2 mm de diámetro mostró la organización típica de la raíz en crecimiento secundario: presentó un estrato de células del súber, felógeno activo, felodermis columnar y lenticelas tipo III. El foema tuvo abundante parénquima axial, grupos aislados de fibras con paredes de espesor mediano a grueso y células tánicas en arreglo solitario o en racimos. El xilema presentó radios parenquimáticos con series de dos a siete células de espesor; los vasos se mostraron solitarios o en arreglo de series radiales cortas de dos a cuatro elementos, en series tangenciales y en series diagonales; vasos punteados de contorno redondeado, oval o angular, con lámina de perforación simple. Parénquima paratraqueal, vasicéntrico completo o incompleto. Fibras con paredes delgadas a gruesas en M. bracteatum y gruesas en M. erythroloma. No se observaron amiloplastos en M. erythroloma, en tanto que su presencia fue abundante en el parénquima axial y radial de xilema y floema de M. bracteatum.
El estudio de las raíces en crecimiento secundario de M. erythroloma y M. bracteatum con diámetros de entre 4 y 6 mm mostró una importante suculencia del eje principal debido a la acumulación de sustancias de reserva (Fig.1, Fig. 2). Las divisiones anticlinales y periclinales del cambium vascular permitieron, en ambas especies, el incremento de las células parenquimáticas del sistema axial y radial del xilema y floema secundarios que presentaron una importante cantidad de amiloplastos, simples, de hilo céntrico en forma de cruz. En la peridermis se observó súber en varios estratos celulares.

Fig. 1. Microfotografía (MOC) de Macroptilium erythroloma en corte transversal: raíz en crecimiento secundario de 4 - 6 mm de diámetro. A: aspecto general de la raíz en crecimiento secundario. B: detalle de vasos del xilema, fibras y parénquima (axial y radial) con amiloplastos. Escala: A = 100 µm; B = 25 µm.
Fig. 1. Microphotography of Macroptilium erythroloma in cross-section: root on secondary growth with 4 - 6 mm diameter. A: general aspect of root on secondary growth. B: detail of the xylem vessels, fibres and parenchyma with starch grains. Scale bar: A = 100 µm; B = 25 µm.
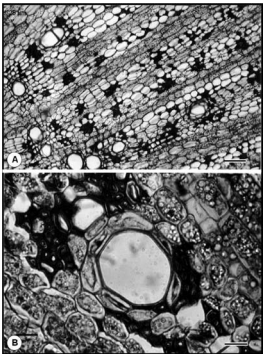
Fig. 2. Microfotografía (MOC) de Macroptilium bracteatum en corte transversal: raíz en crecimiento secundario de 4 - 6 mm de diámetro. A: xilema secundario con vasos solitarios, en series radiales múltiples cortas. B: detalle de fibras, parénquima axial paratraqueal incompleto con amiloplastos. Escala: A = 100 µm; B = 25 µm.
Fig. 2. Microphotography of Macroptilium bracteatum in cross section: root on secondary growth with 4 - 6 mm diameter. A: secondary xylem with solitary vessels, in short radial multiple series. B: detail of fibres, incomplete axial paratracheal parenchyma with starch grains. Scale bar: A = 100 µm; B = 25 µm.
En el floema, las células parenquimáticas de los radios incrementaron su espesor; en el sistema axial las células tánicas se presentaron en mayor cantidad en tanto que las fibras fueron escasas (Fig. 3). En el xilema, el aumento del espesor de los radios fue coincidente con el del floema pues las activas divisiones de las células del cambium provocaron el incremento de las células parenquimáticas tanto del sistema radial como axial, separando los vasos de conducción y ocasionando el aumento del diámetro de las raíces. De esta manera, los vasos se dispusieron casi exclusivamente en arreglos solitarios y en series múltiples cortas. Las fibras del xilema presentaron paredes secundarias gruesas y lignificadas dispuestas en grupos compactos a distancias regulares.
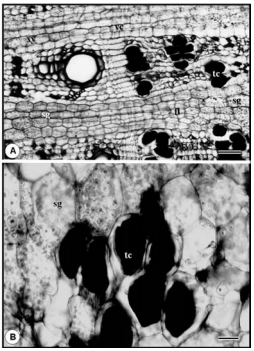
Fig. 3. Microfotografía en corte transversal de raíz de Macroptilium bracteatum con microscopio óptico. A: floema secundario con células tánicas, cambium vascular, xilema secundario con vasos en arreglo de series radiales cortas y parénquima de reserva. B: detalle de células tánicas del floema y parénquima con abundantes amiloplastos. Escala: A = 100 µm; B = 25 µm. f, floema; sg, amiloplastos; tc, células tánicas; vc, cambium vascular; xy, xilema.
Fig. 3. Microphotography in cross section of Macroptilium bracteatum root with optic microscope. A: secondary phloem with tannic cells, vascular cambium, secondary xylem with vessels arranged as short radial series and starch grains. B: detail of tannic cells and parenchyma with abundant starch grains. Scale bar: A = 100 µm; B = 25 µm. f, phloem; sg, starch grains; tc, tannic cells; vc, vascular cambium; xy, xylem.
Raíces gemíferas en Macroptilium bracteatum. La morfología externa de la raíz de M. bracteatum permitió detectar la presencia de yemas en la peridermis. Se visualizaron protuberancias de forma elipsoidal ubicadas en la superficie de la raíz.
En corte transversal se observó que dichas yemas están relacionadas con un grupo de células parenquimáticas que en su base presentaron forma de cuña y se encontraron incluidas en el xilema secundario: estas estructuras en forma de cuña fueron las trazas de las yemas. La formación de las yemas comenzó a partir de las células parenquimáticas radiales del xilema secundario temprano, próximas al xilema primario, a través de divisiones periclinales y anticlinales (Fig. 4 A, B). A partir de dichas células se diferenciaron vasos del xilema con espesamientos helicoidales y helicoidales densos; elementos de tubos cribosos del floema, ubicados longitudinalmente en relación con las células conductoras del xilema (Fig. 4 C, C'). Estas células conductoras correspondieron típicamente al xilema y floema primarios, y limitaron un área central con células parenquimáticas, que constituirán la futura médula del tallo primario.

Fig. 4. Microfotografía en microscopio electrónico de barrido (SEM) de Macroptilium bracteatum en corte transversal: raíz en crecimiento secundario de un año. A: inicio de brote lateral desde células contiguas a los polos del xilema primario (fecha). B: grupo inicial de células parenquimáticas radialmente alargadas (fecha). C: vasos helicoidales y helicoidales densos del xilema primario vistos longitudinalmente en corte transversal de raíz en crecimiento secundario. C': detalle de C. D: yema con sus partes: cono vegetativo y primordios foliares. Escala: A, B, D = 100 µm; C = 50 µm; C' = 25 µm. lp, primordios foliares; vg, cono vegetativo.
Fig. 4. SEM Micrograph of Macroptilium bracteatum in cross section: one-year-old root on secondary growth. A: side sprouting starts from cells contiguous of the poles of primary xylem (arrow). B: initial group of parenchyma cells radially lengthened (arrow). C: helicoidal and dense helicoidal vessels of the primary xylem seen lengthwise in cross section of root at secondary growth. C': detail of C. D: bud with their parts: vegetative cone and leaf primordia. Scale bar: A, B, D = 100 µm; C = 50 µm; C' = 25 µm. lp, leaf primordia; vg, vegetative cone.
En el extremo distal de la estructura en forma de cuña se observaron yemas completamente formadas, con presencia de cono vegetativo y primordios foliares (Fig. 4 D). Estas yemas se clasificaron como adicionales por el origen temprano de sus trazas (Bosela y Ewers, 1997), y pudieron ser seguidas centrípetamente hasta la zona del xilema primario. La mayoría de las trazas se iniciaron contiguas a los polos del protoxilema o a los vasos del xilema secundario formados tempranamente. El cono vegetativo de la yema presentó células meristemáticas típicas, isodiamétricas, con grandes núcleos y paredes primarias delgadas. Se observaron yemas hasta con cuatro primordios foliares;éstas fueron vascularizadas y ubicadas perpendicularmente en relación con el cambium vascular; cuando emergieron, atravesaron la peridermis proyectándose hacia el exterior como una protuberancia cónica.
Las yemas radicales de M. bracteatum fueron funcionales y los vástagos originados formaron nuevas plantas que se independizaron de las plantas madres.
DISCUSIÓN Y CONCLUSIONES
Volenec (2005) sostiene que hay dos factores centrales que impactan sobre el rebrote de las plantas forrajeras: la energía de reserva y los meristemos para el rebrote.
Macroptilium bracteatum y M. erythroloma concentraron la reserva en la raíz, tal como lo indicara Esau (1982) para algunas especies de Apiacea; compartieron características comunes con Macroptilium geophyllum (Quiroga y Pérez, 1996), Galactia (Fabaceae) (Basconsuelo et al., 1997), Rhynchosia (Fabaceae) y Adesmia (Fabaceae) (Basconsuelo y Kraus, 2000) en cuanto al almacenamiento de sustancias de reserva en las raíces. Esto constituye una fuente importante de carbohidratos que permitirá una rápida brotación luego de sobrevivir al estrés ambiental provocado por una estación fría y seca, y por crecer en suelos arenosos y disturbados.
En cuanto a la localización de las reservas, M. bracteatum y M. erythroloma las concentraron en el parénquima del xilema y floema secundarios. En este aspecto difirieron de Galactia marginalis (Basconsuelo, 2001), Adesmia (Kraus et al., 2003), Zornia (Fabaceae) y Poiretia (Fabaceae) (Molina, 2007) por la prevalencia del área reservante, y con M. geophyllum (Quiroga y Pérez, 1996) tanto en la localización de los tejidos de reserva como en el tipo de amiloplastos. Esto es debido a que en M. bracteatum y M. erythroloma, éstos fueron únicamente simples y no presentaron amiloplastos compuestos como en M. geophyllum.
Además, en las especies en estudio no se verificó actividad de cambia supernumerarios ni meristemos laterales como los mencionados por Kraus y Basconsuelo (2009) para Rhynchosia edulis. El cambium vascular fue el único responsable de la suculencia de la raíz, constituyendo un desarrollo del tipo normal o usual (Esau, 1982).
Los individuos estudiados de M. bracteatum y M. erythroloma presentaron complejos axilares con desarrollo completamente vegetativo (Debouck, 1984) y fueron semejantes a los descriptos por Tourn y Roitman (1996) en cuanto a la actividad de las yemas causantes del desarrollo de la corona.
Así entonces, y en acuerdo con Volenec (2005), en M. bracteatum y M. erythroloma la formación de yemas sub-superficiales a partir de una corona basal aseguran su forma de persistencia, y les otorgan perennidad. En tanto, las reservas acumuladas en las raíces brindan la energía necesaria para facilitar el rebrote una vez fnalizada la estación fría y seca.
La formación de yemas radicales en M. bracteatum fue coincidente con lo registrado para Convolvulus arvensis (Convolvulaceae) (Howard et al., 1966) y para Circium arvense (Asteraceae) (Hamdoun, 1970) en cuanto (1) al origen endógeno, (2) la iniciación a partir de las células localizadas en los polos del protoxilema y (3) la observación de que en el ápice de las yemas radicales se produjo un número de primordios foliares antes de la emergencia (Hamdoun, 1970). Las observaciones realizadas sobre las yemas radicales, el origen endógeno, y el estadio de la raíz en que se inició la formación de yemas (primario) y la formación de conexiones vasculares de M. bracteatum coincidieron con las informadas por Raugh (1932) para Coronilla varia (Fabaceae), Howard et al. (1966) para Convolvulus arvensis, y Polokick y Raju (1982) para Asclepias syriaca (Apocynaceae). Las yemas radicales de M. bracteatum respondieron a las características señaladas por Bosela y Ewers (1997) en cuanto a su característica de adicionales. M. bracteatum también compartió la característica de persistencia de las especies por la formación de yemas radicales que le brindan la capacidad de propagarse vegetativamente con Neottia (Orchidaceae), Listera cordata (Orchidaceae), Taeniophyllum proliferum, T. reijnvaanii (Orchidaceae), Ophioglossum (Ophioglossaceae), Anemone canadienses (Ranunculaceae), Rumex acetosella (Polygonaceae), Sium latifolium (Apiaceae) (Raugh, 1932) y Asclepias syriaca (Polokick y Raju, 1982).
M. erythroloma y M. bracteatum tuvieron las características señaladas por Volenec (2005) en cuanto a los factores centrales que impactan sobre el rebrote de las plantas forrajeras: (1) raíces que acumulan energía de reserva en el parénquima axial del xilema y floema, y (2) meristemas para el rebrote que se forman en la corona a partir del primer año de vida de la planta. En el caso de M. bracteatum, se agregó a esta estrategia la formación de yemas radicales adicionales sobre raíces de un año con crecimiento secundario.
Las características morfológicas y anatómicas descriptas, sumadas al valor nutricional registrado para M. erythroloma y M. bracteatum y al consumo por parte del ganado (Camardelli et al., 1996; Pérez de Bianchi et al., 1999b), harían aptas estas especies para ser cultivadas como forrajeras.
AGRADECIMIENTOS
Las autoras agradecen al Laboratory of Scanning Electron Microscopy (LASEM) - ANPCyT UNSa CONICET y al Consejo de Investigación de la Universidad Nacional de Salta (CIUNSa) - Argentina.
REFERENCIAS
1. Basconsuelo, S., R. Malpassi, T. Kraus, C. Bianco y F. Weberling (1997). Growth forms of species of Galactia (Leguminosae) in the southern part of the Province Córdoba, Argentina. Beitraege zur Biologie der Pfanzen 70: 107-119. [ Links ]
2. Basconsuelo, S. y T. Kraus (2000). Morfología del sistema radical de Rhynchosia edulis Gris. (Fabaceae). Phyton, International Journal of Experimental Botany 69: 119-126. [ Links ]
3. Basconsuelo, S. (2001). Morfología de los sistemas radicales en Phaseoleae (Leguminosae) en el área serrana de la Provincia de Córdoba. Tesis Doctoral, Universidad Nacional de Río Cuarto, Argentina. [ Links ]
4. Bosela, M.J. & F.W. Ewers (1997). The mode of origin of root buds and root sprouts in the clonal tree Sassafras albidum (Lauraceae). American Journal of Botany 84: 1466-1481. [ Links ]
5. Camardelli, M.C., S.M. Pérez de Bianchi, E. Quero y H. Carusso (1996). Evaluación agronómica preliminar de la calidad forrajera de Macroptilium erythroloma (Benth.) Urban. Actas de las I Jornadas de Información Científico - Técnicas de la Facultad de Ciencias Agrarias de la Universidad Nacional de Jujuy y I Jornadas Regionales. San Salvador de Jujuy, Argentina. [ Links ]
6. D'Ambrogio, A. (1986). Manual de Técnicas en Histología Vegetal. Editorial Hemisferio Sur, Buenos Aires. [ Links ]
7. Debouck, D.G. (1984). Morfología de la planta de frijol común (Phaseolus vulgaris L.). Centro Internacional de Agricultura Tropical CIAT, Cali, Colombia. [ Links ]
8. Drewes, S. (1997). El Género Macroptilium (Fabaceae) en la Flora Argentina. Boletín de la Sociedad Argentina de Botánica 32: 195-216. Argentina. ISSN 373-580 X. [ Links ]
9. Esau, K. (1982). Anatomía de las Plantas con Semilla. Impreso en la República Oriental del Uruguay. Editorial Hemisferio Sur. S.R.L. (3° Reimpresión - año 1993). [ Links ]
10. Hamdoun, A.M. (1970). The anatomy of subterranean structures of Cirsium arvense (L.) Scop. Weed Research 12: 128-36. [ Links ]
11. Howard, T., J.R. Bonnet y J.G. Torrey (1966). Comparative anatomy of endogenous bud and lateral root formation in Convolvulus arvensis roots cultured in vitro. American Journal of Botany 53: 496-507. [ Links ]
12. Kraus, T.A., C.A. Bianco y F. Weberling (2003). Root system morphology of Fabaceae species from central Argentina. Wulfenia 10: 61-72. [ Links ]
13. Kraus, T. y S. Basconsuelo (2009). Secondary root growth in Rhynchosia edulis Griseb. (Leguminosae): Origin of cambia and their products. Flora 204: 635-643. [ Links ]
14. Molina, M.G. (2007). Morfología de los sistemas radicales en la Tribu Aeschynomeneae (Leguminosae) en el suroeste de la Provincia de Córdoba. Tesis Doctoral, Universidad Nacional de San Luis, Argentina. [ Links ]
15. Pérez de Bianchi, S.M., D.C. Martín Montiel de López, M.C. Camardelli y E. Quero (1996). Incorporación de las especies del Género Macroptilium al Banco Activo Regional de Germoplasma del INTA. Informe Técnico. Código: 20/D020. Consejo de Investigación - Universidad Nacional de Salta. [ Links ]
16. Pérez de Bianchi, S.M., M.C. Camardelli, F. Juárez, A.R. Bianchi y R. Neumann (1999a). Geographic distribution of Macroptilium species in Argentina. Tropical Grasslands 33: 22-33. [ Links ]
17. Pérez de Bianchi, S.M., M.C. Camardelli y H. Caruso (1999b). Producción de Macroptilium bracteatum y M. erythroloma, leguminosas nativas de la República Argentina. IIª Jornadas Regionales de Información Científico-Técnicas de las Facultades de Ciencias Agrarias. Potosí, Bolivia. [ Links ]
18. Polokick, P.L. y M.V. Raju (1982). The origin and development of root buds in Asclepias syriaca. Canadian Journal of Botany 60: 2119-2125. [ Links ]
19. Quiroga, M. y S.M. Pérez de Bianchi (1996). Anatomía de la raíz d Macroptilium geophilum (Burk) Debouck, Juárez et Pérez. Aportes botánicos de Salta. Serie Misceláneas. Volumen 1. N° 4. Herbario MCNS, Facultad de Ciencias Naturales. ISSN 0327-7771. Universidad Nacional de Salta. [ Links ]
20. Raugh, W. (1937). Die Bildung von Hypokotyl - und Wurzelsprossen und ihre Bedeutung für die Wuchsformen der Pflanzen. Nova Acta Leopoldina N. F. 4, Nr 24. [ Links ]
21. Tourn, M.G. y G.G. Roitman (1996). Morphology and growth of Gouania ulmifolia Triana et Planch (Rhamnaceae): An architectural analysis. In: L'arbre. Biologie et development, Edelin, C. ed. Naturalia Monspeliensa, A-7: 666-667. [ Links ]
22. Volenec, J. (2005). Grazing Forages: Taking Care of the Plant. Great Lakes. International Grazing Conference proceedings. Marca Líquida Agropecuaria, Córdoba, 15: 68-70. [ Links ]

