










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.81 no.2 Vicente López jul./dic. 2012
ARTÍCULO ORIGINAL
Variación de la composición y cobertura específicas del pastizal bajo diferentes presiones de pastoreo en el Monte Rionegrino
Variation in specific composition and cover in grassland exposed to various grazing pressures in the Monte Rionegrino
Peter G1,2, FA Funk3, A Loydi4, AI Casalini5, CV Leder4
1 CURZA, Universidad Nacional del Comahue, Esandi y Ayacucho, 8500 Viedma, Argentina.
2 Universidad Nacional de Río Negro, Av. Don Bosco y Leloir, 8500 Viedma, Argentina.
3 CERZOS, CCT CONICET Bahía Blanca, Camino La Carrindanga Km 7, 8000 Bahía Blanca, Argentina.
4 Departamento de Biología, Bioquímica y Farmacia, Universidad Nacional del Sur, San Juan 670, 8000 Bahía Blanca, Argentina.
5 CENPAT, CONICET, Boulevard Brown 2915, 9120, Puerto Madryn, Argentina.
Address Correspondence to: Guadalupe Peter. Tel.: 054-02920-423772 int 146, e-mail: gpeter@unrn.edu.ar
Recibido / Received 25.VI.2011.
Aceptado / Accepted 23.VIII.2011.
Resumen. En el Monte Oriental Rionegrino la vegetación se distribuye formando islas arbustivas dispersas en una matriz de suelo desnudo (espacios interislas). El objetivo de este trabajo fue evaluar la variación en la composición y cobertura específicas del sistema en estudio bajo diferentes presiones de pastoreo. En cada uno de tres sitios se seleccionaron áreas con intensidad de pastoreo alta (IPA), media (IPM) y áreas excluidas al pastoreo (IPB). En cada área se establecieron cinco transectas lineales en las que se midió la cobertura basal y se registraron las especies interceptadas. Los sitios se ordenaron a través de Análisis de Componentes Principales utilizando el programa Infostat. Se analizó la similitud entre sitios considerando la cobertura y las especies presentes en islas e interislas por separado y en las transectas en forma conjunta, sin discriminar islas de interislas. Respecto a la composición específica se diferenciaron las islas de los interislas en todos los sitios, esencialmente por la presencia de especies arbustivas; todas las áreas IPB se diferenciaron de sus respectivas áreas IPA. En cuanto a la cobertura, las áreas IPB fueron definidas por la mayor cobertura de Poa ligularis, broza y microfitas, mientras las áreas IPA presentaron mayor cobertura de suelo desnudo. El pastoreo por sí mismo y a diferentes intensidades modifica la composición específica tanto a nivel de las islas de vegetación como de los interislas. La mayor cobertura de microfitas en las áreas IPB ayudaría a la recuperación de los sistemas.
Palabras clave: Desertificación; Islas de vegetación; Intensidad de pastoreo; Suelo desnudo; Zonas áridas.
Abstract. In the western Monte Phytogeographical Province of Rio Negro, vegetation is a shrubby-grassland steppe, where plants are interspersed within a bare soil matrix forming "phytogenic islands". The objective of this study was to evaluate the specific composition and cover of the study ecological system under diferent grazing pressures. We selected areas with high, moderate and low grazing intensity at three sites. In each area we established five linear transects, where basal cover and all intercepted species were registered. Sites were arranged through a Principal Component Analysis using the Infostat software. We analyzed similarity between sites, considering basal cover and species present in islands and inter-islands, and in both situations together. Islands and inter-islands were separated especially by the presence of shrubby species. Exclosure areas showed a greater cover of Poa ligularis, litter and biological crusts, whereas intense grazed areas presented a greater cover of bare soil. Grazing by itself and diferent grazing pressures can modify the specific composition at an island and inter-island scale. Greater cover of biological crusts could help recovery of the study system.
Keywords: Desertification; Vegetation islands; Grazing intensity; Bare soil; Arid zones.
INTRODUCCIÓN
En las zonas áridas y semiáridas de distintas partes del mundo la vegetación frecuentemente se halla distribuida formando islas arbustivo-graminosas, dispersas en una matriz de suelo desnudo o con escasa cobertura vegetal (Aguiar y Sala, 1999). Las islas de vegetación forman montículos fitogénicos dominados por arbustos (Coronato y Bertiller, 1997; Bisigato y Bertiller, 1999; Cecchi et. al., 2001), debajo de los cuales crecen otros grupos funcionales tales como gramíneas, hierbas y micrófitas.
El pastoreo de grandes herbívoros puede reducir o incrementar la diversidad vegetal en los pastizales. Esto depende, al menos en parte, de la intensidad del pastoreo, de la disponibilidad de recursos, y de la historia evolutiva del pastizal (Milchunas et al., 1988; Mack, 1989; Hobbs y Huenneke, 1992; Olf y Ritchie, 1998; Proulx y Mazumder, 1998). En pastizales poco productivos y que evolucionaron en ausencia de pastoreo intenso se espera que la riqueza específica disminuya en forma lineal con aumentos en la intensidad de pastoreo, mientras que en pastizales productivos y que evolucionaron con pastoreo de grandes herbívoros cabe esperar un efecto positivo (en la riqueza específica) del pastoreo a intensidades intermedias de disturbio.
En el noreste de la Región Patagónica, la actividad ganadera trae asociados problemas de degradación de recursos naturales, tales como la reducción de la cobertura vegetal, la erosión hídrica y eólica de los suelos, y el deterioro físico-químico de los mismos (compactación, encostramiento superficial, pérdida de materia orgánica, etc.). Estos procesos afectan a la mayor parte de los ambientes de planicies aluviales con suelos franco-limosos de la provincia de Río Negro (Kröpf, 1999).
El objetivo del presente trabajo fue evaluar la cobertura y la composición específicas en el Monte Oriental Rionegrino bajo diferentes presiones de pastoreo. Tratándose de un pastizal poco productivo y que habría evolucionado sin presión de pastoreo intenso de grandes herbívoros, cabría esperar que a lo largo de un gradiente decreciente de pastoreo (1) las especies palatables y micrófitas aumenten en su cobertura, (2) disminuya la cobertura de suelo desnudo, (3) varíe la composición específica de las áreas.
MATERIALES Y MÉTODOS
El trabajo se llevó a cabo en campos ubicados en los departamentos de Adolfo Alsina y San Antonio Oeste de la provincia de Río Negro (40° 40' S, 64° 10' O). El clima de la región es subtemplado seco de transición, con veranos cálidos e inviernos moderados, y sin ningún exceso estacional de agua. Las precipitaciones promedian los 230 a 270 mm anuales y presentan una alta variabilidad intra e interanual (Bran et al., 2000). Las estaciones con mayores precipitaciones son otoño y primavera, aunque en invierno puede llover ocasionalmente, en forma de lluvias pequeñas. El clima es ventoso, especialmente en primavera y verano, y ello contribuye a hacer más marcado el déficit de humedad del suelo.
La vegetación que caracteriza el área es una estepa arbustivo-graminosa baja, cuyo estrato herbáceo está formado por un pastizal de crecimiento predominantemente invernal, y corresponde a la Provincia Fitogeográfica del Monte, distrito Sur, Subdistrito Patagonia Norte (Roig et al., 2009). La comunidad está ampliamente extendida en el este de la provincia de Río Negro, y está dominada por Chuquiraga erinacea y Condalia microphylla en el estrato arbustivo y Nassella tenuis en el estrato herbáceo. Entre los arbustos también se encuentran Prosopis alpataco, Senna aphylla, Lycium spp., Monttea aphylla, Schinus johnstonii y Ephedra ochreata. En el estrato herbáceo están presentes Poa ligularis, Piptochaetium napostaense, Jarava plumosa, Pappostipa speciosa, Nassella clarazii, y las anuales Bromus mollis, Schismus barbatus y Erodium cicutarium (Cabrera, 1971; Soreng et al., 2003).
En tres sitios representativos de la estepa arbustivo-graminosa, separados entre sí por una distancia máxima de 30 km, se seleccionaron áreas con diferente intensidad de pastoreo por ganado ovino y bovino: áreas con alta intensidad de pastoreo ubicadas a = 600 m de la aguada (IPA), áreas moderadamente pastoreadas a = 2500 m de la aguada (IPM), y áreas clausuradas o con pastoreo circunstancial de ganado doméstico en los últimos 10 años (IPB). La elección de estas tres áreas de muestreo se debe a que en las zonas áridas existe la formación de piósferas, en donde se produce un gradiente radial de intensidad de pastoreo desde las aguadas (Lange, 1969; Bisigato y Bertiller, 1997). En cada área se establecieron cinco transectas de 20 m de longitud, sobre las cuales se registraron la cobertura basal al centímetro de todas las especies herbáceas interceptadas y la cobertura aérea de las especies leñosas. Además, se registraron las especies interceptadas en los interislas y todas las especies presentes en las islas interceptadas por las transectas. Se consideró como islas a aquellas áreas dominadas por arbustos y con acumulación de sedimentos visible respecto al área que las rodeaba. Se denominaron interislas a aquellas zonas en las que no se observó acumulación de sedimentos.
Se realizó un análisis de similitud entre sitios respecto de la cobertura basal de especies en las cinco transectas de cada sitio e intensidad de pastoreo por separado (N= 45). Finalmente se realizaron análisis de similitud entre sitios respecto de la composición de especies teniendo en cuenta los tres tratamientos y los parches de vegetación (isla e interisla), para los cuales se consideraron las especies presentes en las cinco transectas conjuntamente. Los sitios se ordenaron a través de Análisis de Componentes Principales utilizando el programa InfoStat (Di Rienzo et al., 2008). Se realizaron análisis considerando la presencia de especies en islas e interislas de cada sitio (N= 18) y considerando todas las especies presentes en cada sitio, sin discriminar su presencia en isla o interisla (N= 9).
Los datos de cobertura se transformaron a arcoseno de la raíz cuadrada previo a la realización de los análisis (Zar, 1999).
RESULTADOS
Al ordenar los sitios teniendo en cuenta la cobertura basal de las especies presentes, los dos primeros componentes reunieron el 60% de la varianza acumulada (Fig. 1). Los sitios IPB se agruparon principalmente en el primer cuadrante, definidos por una mayor cobertura de broza, micrófitas y Poa ligularis. Los sitios IPA se ubicaron hacia el segundo cuadrante con una mayor cobertura de suelo desnudo y menor cobertura de los grupos mencionados. Por su parte, los sitios IPM se distribuyeron en el extremo negativo del segundo eje, sin variables importantes que los agruparan.
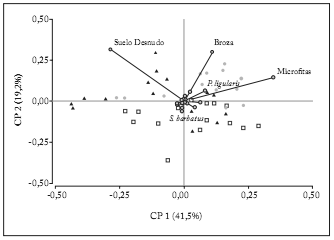
Fig. 1. Ordenamiento de tres situaciones de pastoreo. Para cada situación en cada uno de tres sitios se midió la cobertura basal de especies en cinco transectas.
Referencias: ?= Intensidad de Pastoreo Baja, ?= Intensidad de Pastoreo Media, ?= Intensidad de Pastoreo Alta.
Fig. 1. Arrangement of three different grazing pressures. For each grazing pressure in all three sites, basal cover was determined on five transects.
References: ?= Low Grazing Pressure, ?= Medium Grazing Pressure,?= High Grazing Pressure.
Al ordenar los sitios sobre la base de las especies presentes en islas e interislas, los dos primeros componentes reunieron el 52% de la varianza acumulada (Fig. 2). El primer eje diferenció las islas de los interislas de todos los sitios e intensidades de pastoreo. Las islas tendieron a asociarse con las especies arbustivas (sobre todo Chuquiraga erinacea, Senna aphylla, Schinus johnstonii, Lycium spp., Larrea divaricata, Condalia microphylla, Prosopis alpataco) y las especies herbáceas que tuvieron más importancia en este eje fueron Baccharis ulicina y Pappostipa speciosa. La única especie asociada negativamente a esta componente fue Baccharis melanopotamica.

Fig. 2. Ordenamiento de tres sitios para distintas situaciones de pastoreo. Para cada situación de todos los sitios, las especies se registraron discriminando su presencia en islas e interislas.
Referencias: IPB = Intensidad de Pastoreo Baja; IPM = Intensidad de Pastoreo Media; IPA = Intensidad de Pastoreo Alta; 1 = Sitio 1; 2 = Sitio 2; 3 = Sitio 3. La letra "i" al final de la sigla de cada situación indica que el dato pertenece a la isla, mientras que la ausencia indica interisla.
Fig. 2. Arrangement of three sites exposed to different grazing pressures. For each grazing pressure at all three sites, species were determined at islands or inter-islands.
References: IPB = Low Grazing Pressure; IPM = Medium Grazing Pressure; IPA = High Grazing pressure; 1 = Site 1; 2 = Site 2; 3 = Site 3. The "i" at the end of the reference indicates island data, while its absence reports inter-island information.
Considerando las especies presentes en cada área, sin discriminar su presencia en isla o interisla, los dos primeros componentes reunieron el 45% de la varianza acumulada (Fig. 3). En todos los casos las IPB se diferenciaron de sus respectivas áreas pastoreadas, siendo las IPB 1 y 2 definidas por la presencia de N. clarazii y P. napostaense e IPB 3 por la presencia de L. divaricata. Las especies más asociadas al eje 1 fueron Hofmannseggia trifoliata, Cyclolepsis genistioides, Bredemeyera microphylla y J. plumosa mientras que Lycium spp. se asoció a este eje en oposición a las especies anteriores. Respecto a la segunda componente, las variables más correlacionadas fueron N. clarazii, B. microphylla, Margyricarpus pinnatus y P. napostaense, con C. genistioides en el extremo opuesto.
DISCUSIÓN
Las áreas IPB tendieron a tener mayores cantidades de Poa ligularis, diferenciándose de las áreas IPM e IPA que no presentaron cantidades importantes de esta gramínea perenne palatable. Cabe destacar que algunas de las áreas IPB no se dispusieron en el análisis con las demás zonas de clausura; esto podría deberse a la entrada ocasional de ganado en una de las áreas (sitio 1). Nuestros resultados concuerdan con los obtenidos para el Monte Austral y otros sistemas de la Patagonia en los cuales se señala el efecto del pastoreo sobre el establecimiento y reducción de cobertura de gramíneas perennes (Bertiller, 1994; Bisigato y Bertiller, 1997; Perelman et al., 1997; Bertiller, 1998; Bertiller y Bisigato, 1998; Bertiller et al., 2002; Bisigato y Bertiller, 2004; Bertiller et al., 2009).
Se observó un agrupamiento de las islas en contraposición a los interislas y una variación de la composición específica de las áreas con diferente intensidad de pastoreo. Estas diferencias se deben principalmente a la presencia de especies arbustivas, aunque también están dadas por un pequeño número de especies herbáceas subordinadas (Baccharis ulicina, B. melanopotamica y Pappostipa speciosa) dentro de las islas. Esto sugiere que la presencia de arbustos modifica la composición de especies bajo su canopeo, independientemente de la presencia e intensidad del pastoreo.
En aquellos sistemas que evolutivamente no han sido expuestos a altas presiones de pastoreo, como en el caso de estudio, los mecanismos de resiliencia han tenido menor oportunidad de desarrollarse. Por lo tanto, la introducción de herbívoros en estos sistemas conduce a extinciones irreversibles de especies en las comunidades (Milchunas et al., 1988). Cuando la presión de pastoreo es liberada, la comunidad puede seguir diferentes trayectorias (Cingolani et al., 2005). Esto es evidente en el sitio de estudio, ya que en ninguno de los casos las IPB (clausuras o áreas con entrada ocasional de ganado) forman grupos, lo cual señala diferencias en la recuperación que ha seguido cada una de las áreas tras el retiro de los animales.
Los resultados obtenidos en los análisis composicionales mostraron la diferenciación entre las intensidades de pastoreo dentro de cada sitio. Por lo tanto, nuestros resultados sugieren que, en términos de composición, las modificaciones que el pastoreo produce son evidentes tanto a nivel de las islas de vegetación como de los espacios interislas. Es decir, el pastoreo no sólo modificaría los interislas donde se producen el pisoteo y la defloliación más intensa, sino también las islas de vegetación. Las explicaciones posibles de estos cambios en las islas (donde no se producen pastoreo ni pisoteo intensos) serían la reducción que el pastoreo produce en la biomasa y en el éxito reproductivo de los competidores dominantes; como así también, la oferta y el transporte de propágulos de las especies raras y las oportunidades de generación de propágulos (Olf y Ritchie, 1998). Algunos pastos nativos (P. napostaense, P. lanuginosa, P. ligularis y P. speciosa) fueron más importantes en el análisis de las interislas, lo que resultó suficiente para diferenciar los sitios con menor intensidad de pastoreo de los sitios más intensamente pastoreados. Este efecto de la intensidad de pastoreo en la composición de la vegetación a nivel de islas e interislas en conjunto, quedó expuesto al agruparse las IPB e IPM en una "composición promedio", y diferenciándose las IPA hacia uno u otro extremo (Fig. 3, eje 2).

Fig. 3. Ordenamiento de tres sitios para distintas situaciones de pastoreo. Para cada situación de todos los sitios se registraron las especies presentes sin discriminar su ubicación en islas o interislas. Referencias: IPB = Intensidad de Pastoreo Baja; IPM = Intensidad de Pastoreo Media; IPA = Intensidad de Pastoreo Alta; 1 = Sitio 1; 2 = Sitio 2; 3 = Sitio 3.
Fig. 3. Arrangement of three sites at different grazing pressures. For each grazing pressure at all sites, species were registered without discriminating either island or inter-island location.
References: IPB = Low Grazing Pressure; IPM = Medium Grazing Pressure; IPA = High Grazing pressure; 1 = Site 1; 2 = Site 2; 3 = Site 3.
En conclusión, los cambios en la composición de la vegetación debidos a diferentes intensidades de pastoreo serían más fácilmente reversibles en sistemas con una larga historia evolutiva de pastoreo, debido a su resiliencia. Contrariamente, los sistemas con una corta historia de pastoreo, como el sitio de estudio, poseen menor resiliencia y cambiarían irreversiblemente a estados alternativos al estar sujetos a pastoreo intenso. En el caso de nuestro análisis, y como fuera propuesto en trabajos anteriores (e.g., Beeskow et al., 1995; Bisigato y Bertiller, 1997; Bertiller y Bisigato, 1998; Cingolani et al., 2005; Bertiller et al., 2009), debido a la existencia de múltiples trayectorias y estados irreversibles en respuesta a la variación en intensidad de pastoreo, el retiro del disturbio por sí solo no produciría el retorno del sistema al estado original. Por otro lado, la exclusión de pastoreo aumentaría la oferta de pastos palatables y la cobertura de micrófitas, disminuyendo además la superficie de suelo desnudo. Aunque hay evidencia que indica que las micrófitas facilitan la sucesión (Bowker, 2007), son necesarios más estudios para identificar cómo las costras biológicas (en relación con el pastoreo) podrían ayudar a la recuperación de los sistemas, al conducir la sucesión hacia una trayectoria deseable.
AGRADECIMIENTOS
Agradecemos al personal de los establecimientos donde se realizaron los muestreos. A Roberto A. Distel y Alicia I. Kröpf por sus aportes al trabajo. Este trabajo fue subsidiado con proyectos de CONICET (PIP 112-200801-01046 y ANPCyT-O UNRN 198/10). GP, FAF y AIC pertenecen al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
REFERENCIAS
1. Aguiar, M.R. y O.E. Sala (1999). Patch structure, dynamics and implications for the functioning of arid ecosystems. Tree 14: 273-277. [ Links ]
2. Beeskow, A.M., N.O. Elissalde y C.M. Rostagno (1995). Ecosystem changes associated with grazing intensity on the Punta Ninfas rangelands of Patagonia, Argentina. Journal of Range Management 48: 517-522. [ Links ]
3. Bertiller, M.B. (1994). Modelos ecológicos alternativos de la dinámica de la vegetación en ecosistemas de pastizales: un caso de estudio en la Patagonia. Revista Argentina de Producción Animal 14:15-23. [ Links ]
4. Bertiller, M.B. (1998). Spatial patterns of the germinable soil seed bank in northern Patagonia. Seed Science Research 8: 39-45. [ Links ]
5. Bertiller, M.B. y A.J. Bisigato (1998). Vegetation dynamics under grazing disturbance. The state-and- transition model for the Patagonian steppes. Ecología Austral 8: 191-199. [ Links ]
6. Bertiller, M.B., J.O. Ares y A.J. Bisigato (2002). Multiscale indicators of land degradation in the Patagonian Monte, Argentina. Environmental management 30: 704-15. [ Links ]
7. Bertiller, M.B., L. Marone, R. Baldi y J.O. Ares (2009). Biological interactions at diferent spatial scales in the Monte desert of Argentina. Journal of Arid Environments 73: 212-221. [ Links ]
8. Bisigato, A.J. y M.B. Bertiller (1997). Grazing efects on patchy dryland vegetation in northern Patagonia. Journal of Arid Environments 36: 639-653. [ Links ]
9. Bisigato, A.J. y M.B. Bertiller (1999). Seedling emergence and survival in contrasting soil microsites in Patagonian Monte shrubland. Journal of Vegetation Science 10: 335-342. [ Links ]
10. Bisigato, A.J. y M.B. Bertiller (2004). Seedling Recruitment of Perennial Grasses in Degraded Areas of the Patagonian Monte. Journal of Range Management 57: 191-196. [ Links ]
11. Bowker, M.A. (2007). Biological Soil Crust Rehabilitation in Theory and Practice: An Underexploited Opportunity. Restoration Ecology 15: 13-23. [ Links ]
12. Bran, D., J. Ayesa y C. López (2000). Regiones Ecológicas de Río Negro. Comunicación Técnica Nº 59, Área de Investigación de Recursos Naturales, INTA EEA S. C. De Bariloche. [ Links ]
13. Cabrera, A.L. 1971. Fitogeografía de la Republica Argentina. Boletín de la Sociedad Argentina de Botánica 16: 1-42. [ Links ]
14. Cecchi, G.A., R.A. Distel y A.I. Kröpf (2001). Islas de vegetación en el monte austral: formaciones naturales o consecuencia del pastoreo? XXª Reunión Argentina de Ecología, Bariloche. [ Links ]
15. Cingolani, A.M., I. Noy-Meir y S. Díaz (2005). Grazing efects on rangeland diversity: a synthesis of contemporary models. Ecological Applications 15: 757-773. [ Links ]
16. Coronato, F.R. y M.B. Bertiller (1997). Climatic controls of soil moisture in an arid steppe of northern Patagonia (Argentina). Arid Soil Research and Rehabilitation 11: 277-288. [ Links ]
17. Di Rienzo, J.A., F. Casanoves, M.G. Balzarini, L. Gonzalez, M. Tablada, C.W. Robledo (2008). InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
18. Hobbs, R.J. y L.F. Huenneke (1992). Disturbance, diversity, and invasion: implications for conservation. Conservation Biology 6: 324-337. [ Links ]
19. Kröpf, A.I. (1999). Efecto del pastoreo y otros disturbios sobre la estructura y la dinámica de la vegetación de una estepa arbustiva semiárida. Tesis U.B.A. [ Links ]
20. Lange, R.T. (1969). The piosphere: sheep track and dung patterns. Journal of Rangeland Management 22: 396-400. [ Links ]
21. Mack, R.N. (1989). Temperate grasslands vulnerable to plant invasions: Characteristics and consequences. Biological Invasions: A Global Perspective. En: Drake, J.A., Mooney, H.A., Castri, F.D., Groves, R.H., Kruger, F.J., Rejmánek, M. and Williamson, M. (eds), pp. 155-179. [ Links ]
22. Milchunas, D.G., O.E. Sala y W.K. Lauenroth (1988). A generalized model of the efects of grazing by large herbivores on grassland community structure. American Naturalist 132: 87-106. [ Links ]
23. Perelman, S.B, R.J.C. León y J.P. Bussacca (1997). Floristic changes related to grazing intensity in a Patagonian shrub steppe. Ecography 20: 400-406. [ Links ]
24. Proulx, M. y A. Mazumder (1998). Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient- rich ecosystems. Ecology 79: 2581-2592. [ Links ]
25. Olf, H. y M.E. Ritchie (1998). Effects of herbivores on grassland plant diversity. Tree 13: 261-265. [ Links ]
26. Roig, F.A., S. Roig-Juñent y V. Corbalán (2009). Biogeography of the Monte Desert. Journal of Arid Environments 732: 164-172. [ Links ]
27. Soreng, R.J., P.M. Peterson, G. Davidse, E.J. Judziewicz, F.O. Zuloaga, T.S. Filgueiras y O. Morrone (2003). Catalogue of New World Grasses (Poaceae): IV. Subfamily Pooideae. Contributions from the United States National Herbarium 48: 1-730. [ Links ]
28. Zar, J.H. (1999). Biostatistical analisis, 4th Edition edn. Prentice Hall, Upper Saddle River, New Jersey. [ Links ]

