










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.81 no.2 Vicente López jul./dic. 2012
ARTÍCULO ORIGINAL
Mnesithea selloana: morfología de la sinflorescencia, madurez y calidad de las cariopsis
Mnesithea selloana: morphology of sinflorescence, maturity and caryopsis quality
Galussi AA1,2, ME Moya
1 PID UNER 2132 - LICVEVC.
2 Botánica Morfológica. Facultad de Ciencias Agropecuarias. Universidad Nacional de Entre Ríos. Ruta Prov. 11 km 10,5 Oro Verde, Depto. Paraná, Entre Ríos, República Argentina.
Address Correspondence to: Dr. Alberto A. Galussi. Tel.: +54 343 4975075 - Int. 141; fax +54 343 4975096; e-mail: cultivar@fca.uner.edu.ar
Recibido / Received 30.V.2011.
Aceptado / Accepted 12.XII.2011.
Resumen. Mnesithea selloana (cola de largarto, teyú ruguay) es una especie de ciclo primavero-estivo-otoñal que florece y fructifica durante dicho período, es de buena calidad forrajera, y se halla en vías de desaparecer en los campos naturales de la región. El objetivo de este trabajo fue investigar la morfología de la sinflorescencia, la madurez y calidad de las cariopsis en dicha especie, con la finalidad de conocer aspectos reproductivos en función de la diseminación de la especie en áreas de pastoreo. La sinflorescencia se estudió en dos matas durante un período de 45 días, entre octubre y diciembre de 2010, en un predio de la FCA UNER. Se observaron el crecimiento de los racimos y sus disemínulos. Se determinaron momento de desarticulación, la humedad, el peso y el poder germinativo de las cariopsis. La sinflorescencia estuvo constituida por un eje principal que terminó en un racimo espiciflorme (unidad de floración), y por ejes laterales (paracladios del trofotagma) que repitieron la estructura del eje principal. La sinflorescencia resultante tuvo la apariencia de una panícula. Ambas matas alcanzaron a producir entre 25 y 39 sinflorescencias con un promedio de 9 a 10 racimos, lo que resultó en un total de 254 a 272 racimos por mata, respectivamente. Se produjeron 30-35 disemínulos por racimo, con dos espiguillas, desarticulables a medida que maduraron. La maduración de los disemínulos en los racimos fue basípeta, encontrándose 40% - 50% de disemínulos con cariopsis presentando 32% de humedad una vez desarticulados. La germinación dio hasta 50% de plántulas normales. Los estudios realizados posibilitan establecer estrategias para favorecer la dispersión de la especie a nivel de semillas en áreas de pastoreo.
Palabras clave: Disemínulo; Trofotagma; Paracladio; Germinación; Dispersión.
Abstract. Mnesithea selloana (known as cola de largarto, teyú ruguay) is a spring-summer-fall species that flowers and fructifes in that period; it has good forage quality, and it is in danger of extinction in the natural grasslands at the study region. The objectives of this work were to study the sinflorescence morphology, and the maturity and quality of the caryopsis to know its reproductive aspects in relation to the dissemination of this especies in grazing areas. The sinflorescence was studied in two plants during a period of 45 days, between October and December 2010, within an experimental field of the UNER FCA. Growth of clusters and their disseminules were observed. Time of dehiscence, humidity, weight and germination of caryopsis were determined. Sinflorescence was constituted by a main axis ending in a spiciform raceme (flowering unit), and by lateral axis (paraclades of the trophotagma) which repeated the main axis structure. Te resulting sinflorescence has a panicle appearance. Both plants produced 25 to 39 inflorescences with an average of 9 to 10 racemes, resulting in a total of 254 to 272 racemes per plant, respectively. Thirty to 35 disseminules were produced per raceme with two disarticulated spikelets as they matured. Maturity of disseminules in the racemes was at the base. We found 40% to 50% of disseminules with caryopsis presenting 32% humidity once disarticulated. Germination produced up to 50% of normal seedlings. Our studies made it possible to establish strategies to favor species dispersion through seeds in grazing areas.
Keywords: Disseminules; Trophotagma; Paraclades; Germination; Dispersion.
INTRODUCCIÓN
El género Mnesithea Kunth comprende aproximadamente 30 especies distribuidas en las regiones cálidas del mundo. Pertenece a la familia Poaceae, subfamilia Panicoideae, tribu Andropogoneae, subtribu: Rottboelliinae. En la tribu, las espiguillas se disponen generalmente en pares formando los denominados racimos de espiguillas (Clayton, 1972; Allred, 1982). En la Argentina el género Mnesithea se encuentra representado por 6 especies de las cuales Burkart (1969) cita dos presentes en Entre Ríos: M. selloana (= Coelorhachis selloana) y M. balansae (= C. balansae). Mnesithea selloana (Hack.) de Koning & Sosef, "cola de largarto, teyú ruguay", es una especie de ciclo primavera-estivo-otoñal (megatérmica) que florece y fructifica durante dicho periodo. En Burkart (1969) se menciona que la sinflorescencia es un racimo espiciforme cilíndrico, glabro, recto, erguido, desarticulable, de 7-10 cm de longitud. Además, las espiguillas son múticas por pares en los nudos engrosados de raquis frágiles; una espiguilla es sésil oval, dorsalmente aplanada, adosada al raquis, uniflora, fértil, y la otra espiguilla es pedicelada estéril, muy reducida. Quarín (1979) y Antón (1975) coinciden con la descripción antes mencionada. Además, Antón (1975) sostiene que la sinflorescencia no es ramificada en los nudos superiores, de modo que las cañas principales llevan generalmente racimos solitarios. Otras descripciones de toda la sinflorescencia de esta especie se han encontrado en Vegetti (1997). Clayton (1969) definió a los disemínulos como la unidad formada por la espiguilla sésil, el artejo del raquis, el pedicelo y la espiguilla pedicelada. El objetivo de este trabajo fue investigar la morfología de la sinflorescencia y la madurez y calidad de las cariopsis en Mnesithea selloana, con la finalidad de conocer aspectos reproductivos relacionados con la diseminación de la especie en áreas de pastoreo.
MATERIALES Y MÉTODOS
Se extrajeron 20 matas de M. selloana de un campo con un suelo muy empobrecido (31° 49' 18.62" S; 60° 32' 57.33" O) (Fig. 1a). Dichas matas fueron trasladadas y cultivadas en un predio de la Facultad de Ciencias Agropecuarias en Oro Verde, Entre Ríos, Argentina (31° 49' 59.51" S; 60° 31' 21.84" O) desde noviembre de 2009 (Fig. 1b). Las observaciones se efectuaron entre el 25 de octubre y el 15 de diciembre de 2010, en dos matas de 0,16 m2 (0,40 m x 0,40 m) cada una. Los aspectos estudiados fueron: descripción de la sinflorescencia; número de racimos por unidad de superficie; número de racimos por sinflorescencia, número de disemínulos por racimo; estados de madurez de la sinflorescencia, el racimo y los disemínulos en el tiempo; peso del disemínulo y de la cariopsis: cantidad de disemínulos con o sin cariopsis; humedad de la cariopsis al momento de la desarticulación y posterior a la cosecha, y germinación de las cariopsis.

Fig. 1. Mnesithea selloana a) Área natural de extracción de las plantas, b) Mata cultivada en el predio de la FCA UNER.
Fig. 1. Mnesithea selloana a) Natural area of plant extraction, b) plants cultivated at the UNER FCA experimental field.
El número total de racimos por unidad de superficie en las dos matas se registró durante un periodo de 45 días, con un intervalo de 2 días entre mediciones. Cuando se observó el primer disemínulo emergido de la vaina de la hoja, se consideró el inicio del estado de emergencia de los racimos. Los racimos dejaron de ser contados cuando su disemínulo basal tuvo una coloración amarillento-amarronada.
Se marcaron tres sinflorescencias por mata para determinar el número de racimos por sinflorescencia. Al comenzar la evaluación las sinflorecencias presentaron un único racimo emergiendo, con uno a dos disemínulos emergidos de la vaina. El registro del estado de la sinflorescencia se realizó cada 2 días hasta 45 días, registrando el número de racimos totales emergidos en cada una de ellas.
La evaluación de la madurez de los racimos y el número de disemínulos por racimo se determinó en el primer racimo lateral (paracladio del trofotagma de primer orden, segundo nudo por debajo de la unidad de foración) en dos sinflorescencias por mata. Éstos se marcaron y se observaron los cambios de estados en los disemínulos del racimo desde el ápice hacia la base del mismo. Los estados observados fueron: disemínulos emergidos; disemínulos con anteras y estigmas turgentes; disemínulos con anteras y estigmas secos; disemínulos sin anteras y disemínulos desarticulados. Se contó el número de disemínulos por estado con una periodicidad de 2 días hasta la completa desarticulación de los mismos. La determinación del peso de los disemínulos y de las cariopsis se llevó a cabo cosechando manualmente los disemínulos cada dos días, recolectándose al azar 30 disemínulos de la parte media de los racimos.
El porcentaje de presencia de cariopsis se determinó evaluando 4 repeticiones de 100 disemínulos. Para el peso de disemínulos y cariopsis a través del tiempo, se separaron los disemínulos de acuerdo al estado. Se pesaron 30 individuos para cada estado de madurez y se obtuvo el peso promedio. Para el peso de cariopsis maduros se pesaron 100 cariopsis (10 repeticiones de 10 cariopsis).
La humedad de las cariopsis se determinó por el método de alta temperatura (130-133 °C) durante una hora ± 3 minutos, sin molido ni presecado, en dos repeticiones de 0,5 g, con modificaciones en el peso (ISTA, 2010).
El análisis de germinación de las cariopsis se evaluó sembrando 3 repeticiones de 50 cariopsis entre papel húmedo con agua destilada y con una solución de AG3 (0,05%), se llevaron a estufa a 25 °C, evaluándose cada 4 días durante 17 días. En función de los análisis de semillas efectuados, se definió semilla pura y plántula normal para dicha especie, de acuerdo a las Reglas Internacionales de Análisis de Semillas (ISTA, 2010).
El análisis de los datos para los estudios realizados se efectuó con el software Infostat (Di Rienzo et al., 2010).
RESULTADOS Y DISCUSIÓN
Estructura de la sinflorescencia. La planta de Mnesithea selloana esta formada por un conjunto jerárquicamente ordenado de ejes (macollas) que presentan distintos grados de ramificación, hallándose ejes al estado vegetativo y otros al estado reproductivo. Cada uno de estos ejes al estado reproductivo puede ser considerado una sinflorescencia en el sentido de Cámara Hernández y Rúa (1991) y Vegetti (1997). La sinflorescencia en M. selloana está constituida por el trofotagma y la unidad de floración (Sell, 1976) o sinflorescencia (Fig. 2a). A nivel del trofotagma se originan un número variables de ejes laterales (paracladios del trofotagma) que rematan en una unidad de floración lateral similar a la unidad de floración terminal. La sinflorescencia resultante tiene la apariencia de una panícula (Rúa, 1999), pero dado que este término se aplica para una unidad de floración, y estas sinflorescencias presentan varias unidades de floración, ha sido caracterizada como falsa panoja (Nicora y Rúgolo de Agrasar, 1987). La unidad de floración (tanto la terminal como las laterales) carece de espiguilla terminal y está constituida por paracladios reducidos a un par de espiguillas (paracladios cortos). En conjunto cada unidad de floración ha sido considerada por practicidad como un racimo espiciforme pero dicha denominación es morfológicamente inadecuada (Vegetti, 1997).

Fig. 2. Mnesithea selloana a) Plan estructural de la planta formado por la unidad de floración (UIF) y el trofotagma (TT), Paracladios del trofotagma de primer orden (PCTT), de segundo orden (PCTT´); b) Racimo espiciforme subzona de paracladios cortos; c1) Disemínulo, c2) Espiguilla fértil (con cariopsis), y c3) Espiguilla pedicelada (estéril) (X10); d) Disemínulo con dos espiguillas fértiles con cariopsis (X10); e1) Cariopsis: vista escutelar, e2) Cariopsis: vista hilar (X30).
Fig. 2. Mnesithea selloana a) Plant structural plan formed by the flowering unit (UIF) and the trophotagma (TT), Paraclades of the trophotagma of frst order (PCTT), second order (PCTT´); b) Spiciform racemes, sub-zone of short paraclades; c1) Disseminule, c2) Fertile spikelet (with caryopsis), and c3) Pedicellate spikelet (sterile) (X10); d) Disseminule with two fertile spikelets with caryopsis (X10); e1) Caryopsis: scutellar view, e ) Caryopsis: hilar view (X30).
Según los estudios realizados, la sinflorescencia puede alcanzar una longitud total de 81 cm, con 6 nudos, presentando entrenudos del trofotagma de 16,5 a 11 cm los proximales y de 9 a 6 cm. los distales. Los paracladios del trofotagma encontrados fueron de primero, segundo y tercer orden en número de 4 a 6, hasta 4 y hasta 2, respectivamente. El origen de los mismos es mayormente profilar. El orden de aparición de los racimos en los paracladios del trofotagma de primer orden es basípeto, siendo la secuencia, alterna-dística, nudo 3, 2, 4, 5 y 6; (considerando como primero al nudo por debajo del que porta el racimo). La aparición de los racimos en los paracladios de segundo y tercer orden es acrópeta y sigue el patrón de alternancia dística. En los racimos espiciformes (Fig. 2b), las espiguillas se disponen en el raquis de a pares, constituyendo el disemínulo (Fig. 2c1). Generalmente una espiguilla es fértil, sésil (Fig. 2c2) y puede originar cariopsis y la otra es estéril y pedicelada (Fig. 2c3). También se hallaron disemínulos con dos espiguillas con cariopsis (Fig. 2d) y otras sin cariopsis. La antesis y maduración de las cariopsis en los racimos es basípeta. El cambio de verde claro a rojizo y amarillento amarronado de las espiguillas, expresa el grado de madurez del disemínulo.
El número de sinflorescencias hallado por unidad de superficie (0,16 m2) en el periodo observado fue de 39 y 25 en la mata 1 y 2, respectivamente.
En las sinflorescencias evaluadas, el mayor número de racimos se observó entre los 36 y 38 días, con un promedio de 7 y 10 racimos para las matas 1 y 2, respectivamente (Fig. 3). El número total de racimos hallados en las matas 1 y 2 alcanzó un máximo de 272 y 254, respectivamente, entre los 31 y 36 días desde el inicio de las observaciones (Fig. 4).

Fig. 3. Evolución del número promedio de racimos por sinflorescencia de M. selloana; n: 3 sinflorescencias por mata.
Fig. 3. Evolution of the average number of racemes per M. selloana inflorescence; n: 3 sinflorescences per plant.

Fig. 4. Evolución del número de racimos por mata.
Fig. 4. Evolution of raceme number per plant.
Características de los disemínulos a la madurez y germinación. En el periodo de 9 a 38 días (Mata 1) y de 7 a 38 días (Mata 2) de emergencia de los primeros disemínulos, suceden los estadios, estigmas y anteras turgentes, anteras secas y sin anteras, indicados con líneas verticales en la Figura 5. La desarticulación de los disemínulos en las Matas 1 y 2 se inició entre los 26 y 23 días de emergido el primer disemínulo del racimo, respectivamente, finalizando entre los 18 y 21 días posteriores. Esto significa que en el transcurso de 45 días se produce la desarticulación completa del racimo. Al momento de la desarticulación, las cariopsis contenían 32% de humedad, aunque con secado natural, la humedad disminuyó a 9,5 - 10,5% luego de 10 días.

Fig. 5. Períodos de emergencia y desarticulación de disemínulos en el racimo de la sinforescencia de M. selloana.
Fig. 5. Periods of disseminule emergency and disarticulation at the M. selloana sinforescence racemes.
En la Tabla 1 se observa la evolución del peso de disemínulos y cariopsis hasta la desarticulación (cosecha). En el primer estadio el peso reflejó que aún no hay desarrollo de la cariopsis en el disemínulo. En los tres estadios siguientes se mostró un incremento sensible y variable del peso promedio de los disemínulos debido al desarrollo incipiente de la cariopsis. En el último estadio, los disemínulos con cariopsis presentaron un peso promedio de 10,4 ± 1,6 mg, representando las cariopsis un 21% del peso total del mismo. Los disemínulos vanos al desarticularse presentaron un peso similar (6,7 mg) al peso de los disemínulos del primer estadio. A la cosecha, la diferencia en el peso entre disemínulos se dio por la presencia o no de cariopsis, lo cual podría indicar una posible separación a través de aire y zaranda y mesa de gravedad.
Tabla 1. Disemínulos y cariopsis de Mnesithea selloana en diferentes estadios de madurez.
Table 1. Mnesithea selloana disseminules and caryopsis at different maturity stages.

El número promedio de disemínulos por racimo fue entre 30-35, de los cuales el 40-50% presentaron cariopsis. Cuando el disemínulo presentó cariopsis, generalmente tuvo solo una (Fig. 2c2), aunque también se hallaron disemínulos con dos cariopsis (Fig. 2d). La cariopsis fue pequeña, amarillenta a castaño claro, aovadas y de 1,5 - 3 mm de largo por 1-2 mm de ancho, con embrión basal lateral (Martin, 1946) (Fig. 2e). El peso promedio de la cariopsis madura (base húmeda) fue de 2,20 mg.
A los fines del análisis de semillas, se define como 'semilla pura' a: disemínulo con cariopsis; cariopsis y trozo de cariopsis, mayor que la mitad del tamaño original. Dado que el disemínulo puede contener o no cariopsis, es aconsejable detectar la presencia de cariopsis para un ensayo de germinación. Dicha determinación se debe realizar en forma manual en cada disemínulo, ya que la observación con diafanoscopio, no permite su detección.
Las cariopsis sembradas con un contenido de agua de 9,5% dieron 39% de plántulas normales. En la figura 6 se observa un grupo de semillas de rápida germinación a los 4 días; luego, el número de plántulas normales aumenta gradualmente hasta los 10 días, y permanece constante hasta los 17 días. Al finalizar el ensayo, se hallaron 47% de semillas frescas y 14% de semillas muertas. Similar comportamiento tuvieron las semillas tratadas con AG3, pero con mayor cantidad de plántulas normales (50%), 35% de semillas frescas y 15% de semillas muertas. Dado el comportamiento mencionado, se propone el pretratamiento con AG3, un primer conteo a los 4 días y un segundo a los 10 días, pudiendo prolongarse el ensayo hasta 7 días. Los patógenos necrotróficos hallados externamente en las semillas muertas del ensayo de germinación fueron Alternaria sp. y Curvularia sp., los cuales se consideran que no inciden en la viabilidad del embrión.M. selloana es una Monocotiledónea gramínea con germinación hipógea, con una raíz primaria esencial. La plántula normal (Fig. 7) se asemeja a la descripta para los géneros Lolium, Brachiaria,Bromus, Chloris y otros del Tipo D1 (ISTA, 2006). La parte de la plántula que crece hacia la luz y se torna verde es la hoja primaria, que inicialmente está cubierta por el coleoptilo.
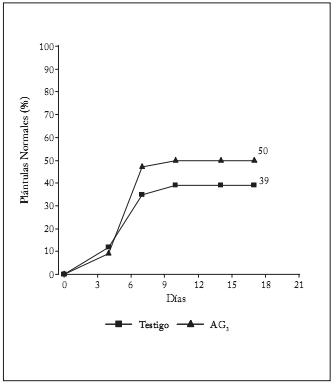
Fig. 6. Germinación de cariopsis de M. selloana con un sustrato humedecido con agua (testigo) y con una solución de AG3 (0,05%). * (% PN) Promedio de 3 repeticiones de 50 cariopsis.
Fig. 6. M. selloana caryopsis germination with a water wet substrate (control) and a AG3 (0.05%) solution. * (% PN) Average of 3 replicates of 50 caryopsis.

Fig. 7. Plántula normal de M. selloana en un ensayo de germinación entre papel a 25 °C. a) 4 días, b) 5 días, c) 7 días, d) 9 días, e) 10 días.
Fig. 7. M. selloana normal seedling in a germination test in paper at 25°C. a) 4 days, b) 5 days, c) 7 days, d) 9 days, e) 10 days.
CONCLUSIÓN
La estructura de la sinflorescencia constituida por la unidad de floración con paracladios cortos y por el trofotagma con numerosos paracladios del trofotagma de primer, segundo y tercer orden, contribuye a la producción de numerosos disemínulos a través del periodo reproductivo. La característica en la sinflorescencia de desarticulación de los disemínulos, la elevada humedad de las cariopsis, así como la mediana cantidad de cariopsis con capacidad para germinar, indicarían la conveniencia de una cosecha manual continua en el período reproductivo, secado (natural) de los disemínulos, y la evaluación del poder germinativo de las cariopsis. Estos conocimientos de la especie puestos en función de la diseminación, permitirían la multiplicación de la especie en campos de pastoreo.
AGRADECIMIENTOS
Al Dr. Ing. Agr. Abelardo Vegetti por su contribución en la descripción de la sinflorescencia de Mnesithea selloana y por la lectura y sugerencias al manuscrito. Al Laboratorio de Fitopatología de la FCA UNER por la determinación de los patógenos hallados en el análisis de germinación.
REFERENCIAS
1. Allred, K.W. (1982). Describing the Grass Inflorescence. Journal of Range Management 35: 672-695. [ Links ]
2. Antón, A.M. (1975). Notas críticas sobre gramíneas de argentina. Kurtziana 8: 61-69. [ Links ]
3. Burkart, A. (1969). Flora Ilustrada de la provincia de Entre Ríos. Colección Científica del INTA. Tomo II, 551 p. [ Links ]
4. Cámara Hernández, J. y G.H. Rua (1991). The synflorescence of Poaceae. Beiträge zur Biologie Pfanzen 66: 297-311. [ Links ]
5. Clayton, W.D. (1969). A revision of the genus Hyparrhenia. Kew Bulletin Addit. Ser. 2: 1-196. [ Links ]
6. Clayton, W.D. (1972). The Awned Genera of Adropogoneae. Studies in the Gramineae: XXXI. Kew Bulletin 27: 457-474. [ Links ]
7. Di Rienzo, J.A., F. Casanoves, M.G. Balzarini, L. González, M. Tablada y C.W. Robledo (2010). InfoStat versión 2010. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
8. ISTA (2006). Handbook on Seedling Evaluation. 3rd. ed. International Seed Testing Association. Switzerland. [ Links ]
9. ISTA (2010). Rules International Seed Testing Association. Switzerland. [ Links ]
10. Martin, A.C. (1946). The comparative internal morphology of seeds. American Midland Naturalist 36: 513-660. [ Links ]
11. Nicora, E.G. y Z. Rúgolo de Agrasar (1987). Los Géneros de gramíneas en América Austral. Ed. Hemisferio Sur. Buenos Aires, Argentina, 661 p. [ Links ]
12. Quarín, C.L. (1979). Los Géneros Rhytachne y Coelorhachis (Gramineae) en Argentina. Kurtziana 12-13: 7-35. [ Links ]
13. Rua, G.H. (1999). Sinflorescencias. Bases teóricas para su análisis. Sociedad Argentina de Botánica. Buenos Aires, Argentina, 100 p. [ Links ]
14. Sell, Y. (1976). Tendances évolutives parmi les complexes inflorescentiels. Revue Générale de Botanique 83: 247-267. [ Links ]
15. Vegetti, A.C. (1997). Sobre la estructura de la sinflorescencia en especies de Rottboelliinae (Poaceae - Andropogoneae). Candollea 52: 475-495. [ Links ]

