










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.82 no.2 Vicente López dic. 2013
ARTÍCULOS ORIGINALES
Floristic diversity of a shrubland in northeastern Mexico
Diversidad florística de un área dominada por arbustos en el Noreste de México
Foroughbakhch1 R, MA Alvarado-Vázquez1, A Carrillo Parra2, JL Hernández-Piñero1, MA Guzmán Lucio1
1 Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas, Depto. de Botánica San Nicolás de los Garza, N.L. México.
2 Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, A.P. 41, 67700 Linares, N.L., México.
Address Correspondence to: R. Foroughbakhch, Universidad Autónoma de Nuevo León, UANL, Facultad de Ciencias Biológicas. Ave. Universidad S/N Ciudad Universitaria San Nicolás de los Garza, Nuevo León. C.P. 66451 México. e-mail: rahimforo@hotmail.com rahim.forough@gmail.com
Recibido / Received 25.IX.2012.
Aceptado / Accepted 2.XI.2012.
Abstract. With the objective of ecologically characterize the woody plant communities, twelve sampling sites were randomly selected based on cartographic material and digital ortophotos from the Tamaulipan matorral located in Northeastern Mexico. In each site, the minimum sampling area was determined. Thereafter quadrats of 10 x 10 m were used in variable numbers for the evaluation of the tree and shrub layers, and of 1 m² for the herbaceous layer. In each quadrat, all individuals of any species were registered, measuring its height and crown projection for the determination of density, canopy cover, frequency and importance value for each species. In order to evaluate the overall community a series of ecological indices were used: richness (Margalef), diversity (Shannon-Weiner), equitativity (Shannon), dominance (Simpson) and similarity (Sorensen and Jaccard). A total of 160 taxa pertaining to 46 families were found. The families with a greater number of taxa were the Poaceae, Asteraceae and Cactaceae with 19, 18 and 14 taxa, respectively, followed by the Fabaceae with 9 taxa, and the Euphorbiaceae and Acanthaceae with 8 taxa. The rest of the families mostly presented less than 3 taxa, and 23 of them were represented by a single taxon. The estimation of the diversity according to the Index of Shannon-Weiner (H') showed that in the herbaceous and shrub strata the communities with greater H' were the same than those of greater richness. The greater values of H' varied between 1.23-1.56. The equitativity index showed a large variation in the tree stratum of the study communities, reaching values between 0.20-0.90.
Keywords: Tamaulipan thornscrub; Diversity; Ecological index; Scrubland; Plant communities.
Resumen. Con el objetivo de determinar las características ecológicas de comunidades vegetales maderables, doce sitios fueron aleatoriamente seleccionados en base a material cartográfico y ortofotos digitales correspondientes al matorral tamaulipeco localizado en el noreste de México. En cada sitio, se determinó el área mínima de muestreo sobre la base de cuadrantes de 100 m2 (10 x 10 m) donde se evaluaron los estratos arbóreos y arbustivos. Para la evaluación del estrato herbáceo se utilizaron cuadrantes de 1 m2 (1 x 1 m). En cada cuadrante todas las plantas fueron identificadas, registrando la altura, proyección de la corona, y la densidad, frecuencia, abundancia y valor de importancia de cada una de las especies. Para poder evaluar el índice de similitud de comunidades vegetales, se usaron una serie de índices ecológicos: índice de riqueza de Malgalef, índice de diversidad de Shannon-Wiener, índice de equitatividad, índice de dominancia de Simpson e índice de similitud de Sörensen-Jaccard. Se registraron un total de 160 taxa perteneciendo a 46 familias. Las familias con un número mayor de taxa fueron Poaceae, Asteraceae y Cactaceae con 19, 18 y 14 taxa, respectivamente, seguido por Fabaceae con 9 taxa, Euphorbiaceae y Acanthaceae con 8 taxa cada uno; el resto de las familias presentaron menos de 3 taxa, y 23 de ellas presentaron solo un taxón. En relación a la diversidad florística, los estratos herbáceos y arbustivos mostraron mayor índice de Shannon-Weiner (H') en comparación al estrato arbóreo, y por ende mayor riqueza en especies. El valor de H' varió entre 1,23-1,56; el índice de equitatividad mostró una mayor variación en el estrato arbóreo alcanzando valores entre 0,20-0,90.
Palabras clave: Matorral tamaulipeco; Diversidad; Índice ecológico; Arbustos; Comunidad vegetal.
INTRODUCTION
Our study was conducted in the coastal plains of the Gulf of Mexico, where vegetation consists of small trees and shrubs, referred to as the "thorny and subinermal matorral", "Tamaulipan thorny scrub" or "Tamaulipan shrubland", in most of the territory (Ludwig et al., 1975; Rzedowski, 1978; Forough-bakhch, 2001). The Tamaulipan shrubland occurs extending from the coastal plain of the Gulf of Mexico to the southern rim of Texas State, USA. In these arid and semi-arid zones of northwestern Mexico a great variability in climatic and edaphic conditions causes extremely diverse shrublands in terms of the species composition, height, density and plant associations. The various species occurring in this region can be categorized in several groups based on their ecological adaptations and forestry use. The vegetative associations are composed of almost 60 woody species, and many of them are important in forestry and silvopastoral activities such as the production of timber, posts, firewood, food, medicines, handicrafts, and forage (Berni et al., 1979; Reid et al., 1987; Bainbridge et al., 1990; Alden, 1995).
Woody plants of dryland forests are an economically important forage resource in the rural population (Maydel, 1996). Grazing and fragmentation at a large scale have been practiced for 350 years in these areas. However, the vegetation of this region has been lately undergoing important changes in structure, floristic composition, and a slow (and perhaps irreversible) degradation. This has been the result of diverse human activities such as the selective plant cutting of various genus: Prosopis, Helietta, Condalia and Acacia (Ffolliott et al., 1982; Pasiecznik et al., 2001; Estrada et al., 2004, 2005).
People have altered the ecological balance over a large surface area because of overexploitation, degradation and elimination of the woody plants from the shrublands to produce wood, timber and charcoal. The result of their activity has been unfortunate for the plant resource sustainability, the elimination of the environmental service that this plant resource offers, and the socioeconomic profits that contribute to the community dwellers. The situation becomes even worse when the incidence of climate disturbances such as drought are considered, which may carry to the desertification of the zone (Burkart, 1976, Eviner, 2003; Fahring, 2003; Bucur, 2006; Carrillo et al., 2008).
Research on structure and use of matorral lands is increasingly important for the sustained well-being of the rural population, who largely depends on the plant resource. Nevertheless, the literature on structural analysis and development of matorral species is insuficient to elucidate the relationships between vegetation changes and causal factors. The objective of this study was to know the floristic diversity of the shrubby vegetation occurring under the current natural conditions. This will contribute to the implementation of a management program for the sustainable utilization of the vegetal resource present in semiarid zones.
MATERIALS AND METHODS
Study area. This study on the growth and structural vegetation analysis was conducted in a typical semiarid zone of the Gulf of Mexico coastal plain. This is because the vegetation of this area is mostly inhabited by a woodland community dominated by small trees and shrubs, commonly referred to as the "thorny and spiny matorral" or the "tamaulipan thorny scrub" (Rzedowski, 1978). The matorral extends from the Gulf coast to the southern rim of Texas State, USA (Foroughbakhch, 2009). The species present are of shrub type with dominance of deciduous thorny elements, but thorns are lacking in some of them. The overall vegetation is in a transition state between the desert microphylous scrub, the submountain matorral, the mesquite shrub woodland and the low thorny forest growing at the low altitudes, low plains and hills (Heiseke & Foroughbakhch, 1985).
The investigation was carried out in an area of 65103 km2 (23° 10' 27'' to 27° 46' 06'' North, 98° 26' 24'' to 101° 13' 55'' West) which included the Rio Grande river hydrological basin (39661 km2), San Fernando-Soto La Marina area (11521.7 km2) and El Salado area (12373 km2).
The climate is in extreme contrasting. The hot and dry climate predominate and, according to the Köpen classification (García, 2004), it is associated to the (1) "B" dry, (2) "Bw" arid or very dry, or (3) "Bs" semi-arid or semi-dry climate. Weather is very hot most of the year, mainly in the plains, but from November to January it is cold. The mean annual temperature is 22.3 °C with a large difference between winter and summer (-2.3 to 41.1 °C). Hails and frosts usually occur every year even after the beginning of the growing season in March. The long-term mean annual precipitation varies from 380 mm (arid zones) to 749 mm (semiarid zones), with two peaks in late May and July - September, and severe drought periods in June and winter. The water budget is unbalanced. The ratio of precipitation to free evaporation is 0.48, and precipitation to potential evaporation is 0.62 (Navar et al., 1999).
The soils of the region are basically stony of Upper Cretaceous siltstone. Commonly, depth of the A horizon is about 30 cm in arid zones, and 40 cm in semiarid zones, with a pH of 7.5 - 8.5 and abundant limestone. The C horizon underneath is about 80 cm deep and contains fissured limestone. In the FAO classification, these soils are classified as rendzine, vertisoil, feozem and castañozem with low organic matter content, and low levels of phosphorus and nitrogen (FAO-UNESCO, 1974). Underground water is hard but not saline.
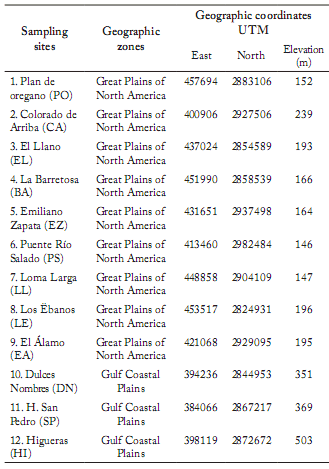
Selection of sampling sites. Sampling sites were delimited based on thematic maps of land use and vegetation, scale 1:50000, and digital ortophotos published by the INEGI (Medina & Natividad, 1998). They were numbered in a progressive form and selected at random to determine the minimum size of the quadrat, according to the species-area curve (Mueller-Dombois & Ellenberg, 1974; Franco et al., 2001). Twelve of these sites were visited for their correct localization, and the recognition of the existent vegetal communities. On each selected site, a GPS receiver (Global Positioning System) was used to register the geographical coordinates and the elevation above the sea level (Table 1).
Table 1. Sampling sites in northeastern Mexico and their geographical coordinates.
Tabla 1. Sitios de muestreo en el noreste de México y sus coordenadas geográficas.
Vegetation structure analysis. The minimum area at each site was estimated following Mueller-Dombois & Ellenberg (1974) and Salvador & Álvarez (2004) to obtain a representative botanical composition. The species occurring in each subsample were registered, and the minimum sampling area was determined as the surface where at least 95% of the species of the plant community were present.
A 10 x 10 m quadrat was delimited at random within each study site. The average distance among plots varied according to the botanical composition and the heterogeneity degree. All the tree, shrub and herbaceous species present within each of the quadrats were identified. Measurements of plant (1) height (m), (2) basal diameter (cm), (3) crown projection area (m2), (4) dominance (absolute and relative), (5) frequency (%) and (6) density (absolute and relative) were obtained (Avery & Burkhart, 2002; Diéguez et al., 2003). The following equations were employed to obtain some of the measured variables (Graf & Sayagues, 2000; Mostacedo & Fredericksen, 2000):
Absolute density = Number of individuals of any given species / Sampling area
![]()
Absolute dominance of any given species = Mean basal area / tree x No of trees of the study species
Relative dominance of any given species =
Relative frequency of any given species =
The importance value (IV) was calculated for each species according to the following equation:
Diversity and Similarity between communities. The parameters given below (i.e., richness, diversity, dominance and similarity) were estimated as follows:
Richness: The index of Margalef (Dmg) was selected to consider the relation between the number of occurring species in the community or stratum (S) and the total number of individuals of all species (N) (Gurevitch et al., 2002):
Dmg = S-1/ ln N
Diversity: The index of Shannon-Weinner (H') was chosen to have a measurement of the diversity of species and its proportional abundance in the community (Franco et al., 2001):
H'= -∑ [pi ln (pi)]
Where: pi = the proportion of individuals in the ith (pi = ni/N),
n = number of individuals of the ith species,
N = number of individuals of all the species.
Equitativity: It is a measurement of the balance in the abundance of any species in the community. In this case a measurement derived from the index of Shannon-Weiner (H') was used. Calculations were made following Gurevitch et al. (2002):
E= H'/ln S (see H' and S above for explanation)
Dominance: The index of Simpson (L) was used to estimate the probability that two individuals taken at random from the community would belong to the same species:
L = ∑ pi2 (Gurevitch et al., 2002)
Similarity: The similarity between communities was estimated at a general level, and for each one of the evaluated strata. For their estimation, the qualitative indices of Sorensen and Jaccard were used as follows:
Jaccard Index: Sj = a / a + b + c
Sörensen Index SSD = 2a / 2a + b + c
Where: a = No. of species at both sites; b = No. of species at the second site, and c = No. of species at the first site.
RESULTS
Floristic Composition. A total diversity of 160 taxa pertaining to 46 families was found. The families with greater number of taxa were Poaceae, Asteraceae and Cactaceae with 19, 18 and 14 taxa respectively, followed by Fabaceae with 9 taxa, and Euphorbiaceae and Acanthaceae with 8 taxa in both cases. The rest of the families mostly presented less than 3 taxa, and 23 of them were represented by a single taxon. The most common species present in all sampled communities were Celtis pallida, Opuntia leptocaulis, Prosopisglandulosa and Ruellia nudiflora. Other species, also common at the study sites, included Acacia rigidula, Acleisanthes obtuse, Castela texana, Dyschoriste decumbens, Karwinskia humboldtiana, Opuntia engelmannii, Tridens muticus and Ziziphus obtusifolia.
Density, Canopy Cover and Similarity between Communities. The total number of species in any given community varied from 22 to 75, with a general average of 46.91 species per community. The vegetal stratum with the highest diversity was the herbaceous layer, whereas the tree stratum displayed the lowest diversity.
The average density of individuals per site was 274015.48 individuals/ha (range= 97900 - 466233), where 91% of them corresponded to the herbaceous layer.
The average cover per site was 203, 2%, taking into consideration that overlapping between individuals might occur between the accumulated cover from the three strata, and even from the interior of each stratum. The maximum vegetation cover was reached in the locality "Dulces Nombres" (Gulf Coastal Plains) with 314, 08%, where vegetation was very dense and uncovered soil surface area was small. On the other hand, the minimum canopy cover was reached in "Plan de Orégano" (108.37%) on the Great Plains of North America; at this location, the plant community had very few tree individuals and large uncovered soil surface areas.
All communities located on the Great Plains of North America showed a greater similarity than those at the Gulf Coastal Plains, with values between 71% ("Plan de Oregano") and 49% ("Loma Larga") (Table 2). Within the Gulf Coastal Plains, "Hacienda San Pedro" showed a similarity in most cases lower than 40%, and "Puente del Río Salado" and "Higueras" in the range from 30 to 50%.
Table 2. Similarity matrix between communities (considering the three layers) according to the indices of Jaccard and Sorensen.
Tabla 2. Matriz de similaridad entre comunidades (considerando los tres estratos) de acuerdo a los índices de Jaccard y Sorensen.
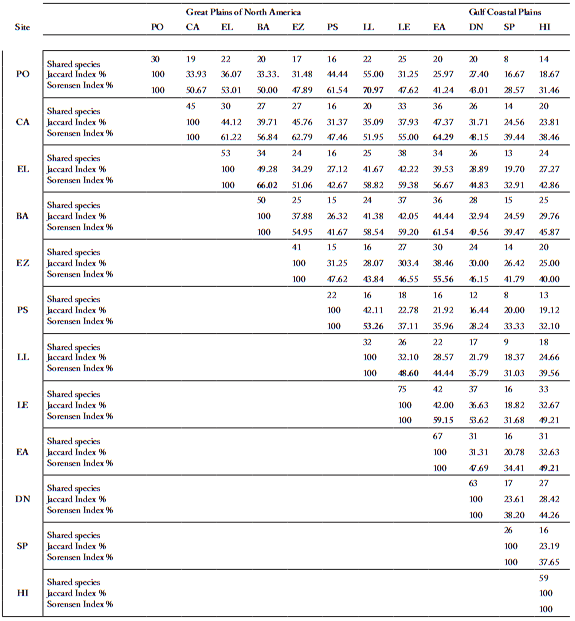
PO= Plan de oregano, CA = Colorado de Arriba, EL = El Llano, BA= La Barretosa, EZ= Emiliano Zapata, PS= Puente Río Salado, LL= Loma Larga, LE=
Los ébanos, EA= El Alamo, DN= Dulces Nombres, SP= San Pedro, HI= Higueras.
Frequency. In the shrub and arboreal strata there were a total of 62 taxa in the 12 localities, with an average of 21.75 taxa per site. Three species Celtis pallida, Opuntia leptocaulis, and Prosopis glandulosa appeared in all the study communities, and showed the greatest values of frequency (100%). The high frequencies of Opuntia engelmanii (91.67%) and Acacia rigidula, Castela texana, Karwinskia humboldtiana and Ziziphus obtusifolia were also relevant, and all of them were present in 10 sites (83.33%). The rest of the species exhibited smaller frequencies, and approximately a third of them were hardly present in a single site (Table 3). Acacia rigidula was second in the rank, despite occurring in 50% of the localities. The rest of the species presented smaller frequencies and three species (Acacia farnesiana, Cercidium sp. and Havardia pallens) appeared only in one community.
Table 3. Species with higher frequency (%) in the herbaceous, shrub and arboreal strata in the studied communities.
Tabla 3. Especies con mayor frecuencia (%) en los estratos herbáceo, arbustivo y arbóreo en las comunidades estudiadas.
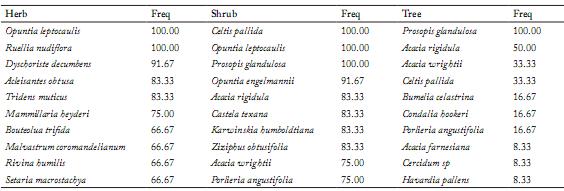
Freq= Frequency.
Freq = Frecuencia, Herbs= Hierbas, Shrub= arbusto, Tree = árbol.
Prosopis glandulosa, a component of the tree stratum, showed quite greater frequency values than those observed for the herbaceous stratum. However, its relative frequency was low in many of the sites, with values generally smaller than 5%, although in the "Hacienda San Pedro" it reached a relative frequency of 20%.
Importance Value (IV). Prosopis glandulosa was the species that presented the greatest importance value (IV) in the tree stratum of all communities, with an average of 80%, followed by Havardia pallens and Acacia wrightii, which showed average values of 21 and 14%, respectively (Table 4). The higher importance value for all strata was obtained by P. glandulosa in all communities. Its IV reached 100% in three of the study communities, more than 75% in six communities, and it exceeded 40% in three of the remaining communities. The species with greater IVs after P. glandulosa were H. pallens (21%), A. wrightii (19%) and A. rigidula (18%).
Table 4. Species with higher average importance values (IV) in the herb, shrub and arboreal strata from the studied communities.
Tabla 4. Especies con mayores valores de importancia (IV) promedio en los estratos herbáceo, arbustivo y arbóreo de las comunidades estudiadas.
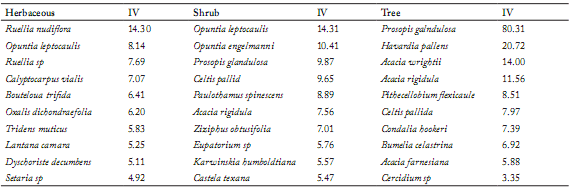
For the Shrub and herbaceous strata only those species occurring in at least 25% of the sites were considered. In the tree stratum all species were considered.
IV = Valor de importancia, Shrub = Arbustos, Tree = árbol.
Diversity Parameters. Richness (Dmg). Within the shrub stratum, the community with greater richness [according to the index of Margalef (Dmg)] was "Dulces Nombres" with a Dmg = 5.54, followed by "El Llano" (Dmg = 5.02) and "Los Ebanos" (Dmg = 4.86). The shrub communities with smaller richness were: "Loma Larga" and "Hacienda San Pedro" (Dmg = 2.73 in both cases), and "Puente Rio Salado" (Dmg = 2.00) (Table 5).
Table 5. Diversity values obtained in the sampling sites.
Tabla 5. Valores de diversidad obtenidos en los sitios de muestreo.
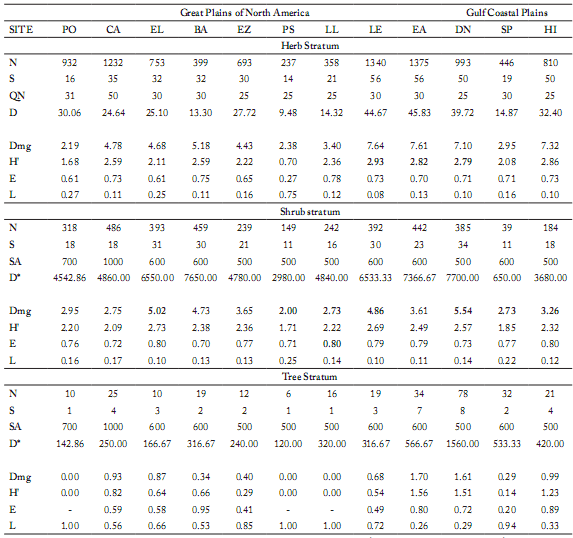
N = number of sampled individuals; S = Number of species; QN = quadrat number (1 m²); D = Density (# individuals/m²); Dmg = Margalef index (Richness); H' = Shanon Weiner's index (Diversity); E = Equitativity of Shannon; L = Simposn's index (Dominance); SA = Sampling area; D*= Density (# individuals/ha); D = Density (# individuals/m2).
PO= Plan de oregano, CA = Colorado de Arriba, EL = El Llano, BA= La Barretosa, EZ= Emiliano Zapata, PS= Puente Río Salado, LL= Loma Larga, LE= Los ébanos, EA= El Alamo, DN= Dulces Nombres, SP= San Pedro, HI= Higueras.
Diversity (H'). Those communities with greater H' were the same than those with greater richness in the shrub and herbaceous strata. Within the shrub stratum, the greater values of H' were found in "El Llano" (2.73), "Los Ebanos" (2.69) and "Dulces Nombres" (2.57). The communities with smaller diversity were "Colorados de Abajo" (H'= 2.09) and "Hacienda San Pedro" (H'= 1.85) in the shrub stratum, and "Puente del Río Salado" (H'= 0.70) in the herbaceous stratum (Table 5).
Equitativity (E). The equitativity in the shrubby stratum reached its highest value in the communities "El Llano", "Loma Larga" and "Higueras", with a value of E = 0.80. On the other hand, the communities with minor equitativity values were "Colorados de Abajo" (E = 0.72), "Puente del Río Salado" (E = 0.71) and "La Barretosa" (E = 0.70) (Table 5).
Dominance (L). The community with greater dominance in the herbaceous and shrub strata was "Puente del Río Salado" (Great Plains of North America) with values of L = 0.75 and L = 0.25, respectively. It was followed by "Hacienda San Pedro" (Gulf Coastal Plains, L = 0.22) in the shrub stratum. The rest of the communities displayed dominance values in the rank of 0.08-0.17, being "El Llano" (L = 0.10) and "Los Ebanos" (L = 0.10) (Great Plains of North America) or "Los Ebanos" (L = 0.08) the communities with the smallest dominance values in the shrub or herbaceous stratum, respectively (Table 5).
DISCUSSION
Comparisons between different ecosystems in semiarid zones are important to determine general trends to find out explanatory variables that could predict changes that may occur after any alteration to the environment due to usual anthropogenic activities in those areas. However, these trait comparisons need to be made in an integrated manner between the diferent disciplines involved in the study.
The number of total species by community varied enormously from the Great Plains of North America ("Puente del Río Salado", "Los Ebanos") to the Gulf Coastal Plains ("Higueras" and "H. San Pedro"). The most common species, considering those present in all the sampled communities, were Ruellia nudiflora, Opuntia leptocaulis, Celtis pallida and Prosopis glandulosa. Other species also common in the study sites were Opuntia engelmannii, Dychoriste decumbens, Acacia rigidula, Castela texana, Karwinskia humboldtiana, Ziziphus obtusifolia, Acleisanthes obtuse and Tridens muticus.
It is important to emphasize that we detected the presence of two endangered species during the study in compliance with the Ofcial Mexican NOM-059-SEMARNAT-2001 (SEMARNAT, 2002). The species were Echinocereus poselgeri (Cactaceae), which has the status "subject to special protection", and Manfreda longiflora (Agavaceae), with "threatened status". In both cases, deforestation and extensive cattle production are the factors that contribute most to that negative impact, and inflict loss and deterioration of plant habitats. Another relevant factor for survival is the nodricismo required by the species as in the case of E. poselgeri. Likewise, the main problem for the establishment of M. longiflora is the modification of the habitat, and the succulent character of the plant, which makes it attractive to predators. Similar results were obtained by Velazco-Macias et al. (2011).
In the herbaceous layer, maximum and minimum density values were found in the communities at "El Alamo" and "Puente del Río Salado", respectively.
Regarding the average cover by site, it was observed a great overlapping between individuals as a result of the accumulated cover from the three strata, and even from the interior of each layer. The maximum vegetation cover was reached in the locality "Dulces Nombres", community with very dense vegetation and little uncovered surface area. On the other hand, the minimum canopy cover appeared in "Plan de Orégano", which had a vegetal community with very few trees and large, open uncovered soil surface areas.
According to the indices of Jaccard and Sörensen, the communities with greater similarity were "Plan de Oregáno" and "Loma Larga", "El Llano" and "La Barretosa", and "Colorados de Arriba" and "El Alamo". The communities that showed least similarity with those remaining were "Hacienda San Pedro", "Puente del Río Salado" and "Higueras", with values in the rank from 30 to 50%.
The general appearance of thornscrub communities is determined by the occurrence of small trees from 3 to 6 m in height present in the shrub and tree strata, which are generally thorny species that vary in density and size (Bainbridge et al., 1990). Humidity plays an important role in determining the exuberance degree and the species associated to the thornscrub and woodland, which affect the density and the insolation degree throughout the depth of the plant layer. It might be observed that the presence of cetaceous species is common under higher insolation conditions, whereas the occurrence of climbing plants associated to different strata is favored under shady vegetation areas.
The physiognomy of the studied sites was dominated by shrub and tree plants. The most outstanding species by its height, density and cover were H. pallens, A. wrightii, A. rigidula, E. ebano and C. pallida. In all the studied sites, the mesquite populations corresponded to the species P. glandulosa. These results are in concordance with those of Jurado & Reid (1989) and Martinez & López (2003).
The most common species in addition to P. glandulosa in the tree stratum were Acacia rigidula, Acacia wrightii, Celtis pallida, Bumelia celastrina and Condalia hookeri.
In the shrub layer, the most common species were Celtis pallida, Opuntia leptocaulis, P.glandulosa, O. engelmannii, Acacia rigidula, Castela texana, Karwinskia humboldtiana and Ziziphus obtusifolia. In the herbaceous layer, the dominant species were Ruellia nudiflora, Opuntia leptocaulis, Dyschoriste decumbens, Acleisanthes obtusa, Tridens muticus and Mammillaria heyderi.
In general terms, the increase in the number of stems (and consequent enlargement of the crown area) plays an important role in the structure and distribution of thornscrub communities. This criterion was particularly noteworthy in the case of Prosopis spp; in most of the studied sites, it was common to observe its presence in disturbed areas due to cutting of trees for the supply of wooden pole fence stands and vegetal charcoal. Growth and proliferation of secondary stems as a consequence of damage or injury to plants of that species has been described elsewhere (e.g., Felker, 1996; Harris et al., 1998). Most woody trees in thornscrub communities are composed of only one stem each in undisturbed areas, but they tend to sprout out several branches after occurrence of any disturbance.
CONCLUSIONS
The physiognomy of the studied sites was dominated by shrub and trees species where Prosopis glandulosa, Acacia spp., Bumelia celastrina, Celtis pallida and Condalia hookeri were the most outstanding vegetation by its height, density and cover.
We found 160 taxa belonging to 46 families, being the families Poaceae [19], Asteraceae [18], Cactaceae [14], Fabaceae [8] and Euphorbiaceae [8] the most important in number. The total number of species by community varied from 22 ("Puente del Río Salado") to 75 ("Los Ebanos") with a general average of 46.91. The vegetal stratum with the highest diversity was the herbaceous layer with 136 taxa (and an average of 34.25 per studied site). The tree stratum displayed the smallest diversity with 13 taxa, and a mean value of 3.17 taxa per site.
The most common species in addition to P. glandulosa in the arboreal stratum were Acacia rigidula, Acacia wrightii, Celtis pallida, Bumelia celastrina and Condalia hookeri.
In the shrub layer, the most common species were Celtis pallida, Opuntia leptocaulis, P. glandulosa, O. engelmannii, Acacia rigidula, Castela texana, Karwinskia humboldtiana and Ziziphus obtusifolia. In the herbaceous layer, the dominant species were Ruellia nudiflora, Opuntia leptocaulis, Dyschoriste decumbens, Acleisanthes obtuse, Tridens muticus and Mammillaria heyderi.
The average density of individuals per site considering the three strata was 274015 individuals/ha, where 268400, 413 and 5177 plants/ha belonged to the herbaceous, shrub and tree layers, respectively.
The greatest importance value (IV) was shown by Ruellia nudiflora and Opuntia leptocaulis in the herbaceous stratum, P. glandulosa, O. leptocaulis and O. engelmannii in the shrub layer, and P. glandulosa for the tree layer.
The communities with the greatest floral richness (Dmg = 7.64) and diversity (H'= 2.93) were at "Los Ebanos" in the herbaceous layer, "Sweet Names" (Dmg = 5.54, H'= 2.69, respectively) in the shrub layer, and "the Alamo" (Dmg = 1.70, H'= 1.76, respectively) in the tree stratum.
The communities with greatest similarity values were located at "Plan del Orégano", "Loma Larga", "El Llano", "La Barretosa", "Colorados de Arriba" and "El Alamo", which were situated in the Great Plains of North America. The communities that showed the lowest similarities were at the "Puente del Río Salado", "Hacienda San Pedro" and "Higueras", these last two located in the Gulf Coastal Plains, with values of similarities ranking from 30 to 50% in comparison to the rest of the communities.
REFERENCES
1. Alden, H.A. (1995) Hardwoods of North America. General Technical. Report FPL-GTR 83 USDA Forest Service Products. Laboratory, Madison, Wis. 136. [ Links ]
2. Avery, E. T. & H. Burkhart (2002). Forest measurements. Fifth edition. Mc. Graw Hill, New York, pp 144-167. [ Links ]
3. Bainbridge, D. A., R.A. Virginia & W.M. Jarrell (1990). Honey mesquite. A multipurpose tree for arid lands. Network Forest Tree. Winrock International. Morrilton, Arkansas. USA, pp 4. [ Links ]
4. Berni, C.A., E. Bolza & F.J. Christensen (1979). South American timbers: the characteristics, properties and uses of 190 species. Div. Bldg. Res. CSIRO, Melbourne, Australia. [ Links ]
5. Bucur, V. (2006). Acoustics of wood. 2nd ed. Springer Series in Wood Science, Springer, Berlin, Heidelberg, Germany, 387 p. [ Links ]
6. Burkart, A. (1976). A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). Journal of Arnold Arboretum 57: 450-525. [ Links ]
7. Carrillo, A., I. Mayer, G. Koch & F. Hapla (2008). Wood anatomical characteristics and chemical composition of Prosopis laevigata grown in the northeast of Mexico. IAWA Journal 29: 25-34. [ Links ]
8. Diéguez, A.U., M. Barrio A., F. Castedo D., A.D. Ruíz G., M.F. Alvaréz T., J.G. Alvaréz G. & A. Rojo A. (2003). Dendrometría. Ediciones Mundi-Prensa. Madrid, España. 327 p. [ Links ]
9. Estrada, C.E., M.C. Yen, S.A. Delgado & Q.J. Villarreal (2004). Legumbres del centro del estado de Nuevo León, México. Anales del Instituto de Biología. Universidad Nacional Autónoma de México. Serie Botánica 75: 73-85. [ Links ]
10. Estrada, C.A.E. (2005) Legumbres del estado de Nuevo León, México. In: R. Foroughbakhch, M.A. Alvarado, T. E. Torres, J.S. Marroquín (Eds.). Tópicos selectos de Botánica 2. Departamento de Botánica. Facultad de Ciencias Biológicas. Universidad Autónoma de Nuevo León, pp.73-94. [ Links ]
11. Eviner, V.T. (2003). Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystem processes. Annual Review of Ecology, Evolution and Systematics 34: 455-485. [ Links ]
12. Fahring, L. (2003). Effects of the habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34: 487-515. [ Links ]
13. FAO-UNESCO (1974). Soil Map of the World, Vol. I. UNESCO, Paris. [ Links ]
14. Ffolliott, P.F. & J.L. Thames (1982). Manual sobre taxonomía de Prosopis en Mexico, Perú y Chile. Universidad de Arizona. Tucson, Arizona, E.U.A Organización de las Naciones Unidas para la Alimentación y Agricultura. [ Links ]
15. Felker, P. (1996). Commercializing mesquite, leucaena and cactus in Texas. In: J. Janick (ed.) Progress in new crops. ASHS Press. Alexandria, VA. 137 p. [ Links ]
16. Foroughbakhch, R., L.A. Hauad, A.E. Céspedes, E.E. Ponce & N. González (2001) Evaluation of 15 indigenous and introduced species for reforestation and agroforestry in northeastern Mexico. Agroforestry Systems 51: 213-221. [ Links ]
17. Foroughbakhch, R., J.L. Hernández-Piñero, M.A. Alvarado-Vázquez, E. Céspedes Cabriales, A. Rocha-Estrada & M.L. Cárdenas-Avila (2009). Leaf biomass determination on woody shrub species in semiarid zones. Agroforestry Systems 77: 181-192. [ Links ]
18. Franco, L.J., A.G. de la Cruz, G.A. Cruz, R.A. Rocha, S.N. Navarrete, M.G. Flores, K.E. Miranda, C.S. Sánchez, L.G. Abarca & S.C. Bedia (2001). Manual de ecología. Ed. Trillas. Sexta Reimpresión, 266 p. [ Links ]
19. García, E. (2004). Modificaciones al sistema de clasificación climática de Köppen. 5th edition. Instituto de Geografía, Universidad Nacional Autónoma de México, 90 p. [ Links ]
20. Graf, E. & L. Sayagues (2000). Muestreo de vegetación. Unidad de sistemas ambientales. Facultad de Agronomía. Universidad de la República, 41p. [ Links ]
21. Gurevitch, J., S.M. Scheiner & G.A. Fox (2002).The Ecology of Plants. Sinauer Associates, Inc. Sunderland, Massachusetts, USA, 523 p. [ Links ]
22. Harris, P.J.C., N.M. Pasiecznik, M. Bradbury & L. Ramírez (1998). Problems and potential of Prosopis. In: Prendergast H.D.V., Etkin N.L. Harris D.R. & P.J. Houghton (Eds.). Plants for food and medicine. Royal Gardens, Kew. U.K. 293 p. [ Links ]
23. Heiseke, D. & R. Foroughbakhch (1985). El matorral como recurso forestal. Reporte Científico No. 1. Facultad de Ciencias Forestales. UANL. Linares, N. L. México. 31 p. [ Links ]
24. Jurado, E. & N. Reid (1989). Influencia de factores edáficos, topográficos y perturbación sobre el matorral espinoso tamaulipeco en Linares, N.L., México. Reporte Científico Facultad de Ciencias Forestales UANL México Nº 10, pp. 1-26. [ Links ]
25. Ludwig, J. A., J.F. Reynold & D.D. Whitson (1975). Size biomass relationships of several Chihuahuan desert shrubs. American Midland Naturalist 94: 451-461. [ Links ]
26. Martínez, AJ. & P.J. López (2003). Growth and Architecture of Small Honey Mesquites under Jackrabbit Browsing: Overcoming the Disadvantage of being Eaten. Annals of Botany 92: 365-375. [ Links ]
27. Maydel, H.J. von (1996). Appraisal of practices to manage woody plants in semiarid environment. In: S. Bruns, J. Furberg, O. Luukanen & P. Woods (Eds.). Dryland Forestry Research. Proceedings of IFS/IUFRO Workshop on dryland forestry research. Hyytiala, Finland July 31-August 4, 1995. International Foundation for Science, Stockholm, Sweden 264 p. [ Links ]
28. Medina, T.J.G. & B.L.A. Natividad (1998). Recursos naturales. Planeación integral. Ed. Trillas, 220 p. [ Links ]
29. Mostacedo, B. & T.S. Fredericksen (2000). Manual de métodos básicos de muestreo y análisis en ecología vegetal. Proyecto de Manejo Forestal Sostenible (BOLFOR) Santa Cruz Bolivia. Ed. Daniel Nash. Bolivia, 87 p. [ Links ]
30. Mueller-Dombois, D. & H. Ellenberg (1974). Aims and methods of vegetation ecology. Wiley & Sons Inc. 452 p. [ Links ]
31. Navar, J., F. Charles & E. Jurado (1999). Spatial variations of interception loss components by Tamaulipan thornscrub in northeastern Mexico. Forest Ecology and Management 124: 231-239. [ Links ]
32. Pasiecznik, N.M., P. Felker, J.C. Harris, L.N. Harsh, G. Cruz, J.C. Tewari, K. Cadoret & L.J. Maldonado (2001) The Prosopis juliflora - Prosopis pallida Complex: A Monograph. HDRA Coventry UK. Forestry Research Program. Department for International Development. 162 p. [ Links ]
33. Reid N, S.M. Stafford, P. Bayer-Munzel & J. Marroquin (1987). A research strategy for ecological survey: Floristic and lands use in the tamaulipan thornscrub, northeastern Mexico. In: Strategies for classification and management of native vegetation for food production in arid zones. General Technical Report RM, Tucson, Arizona, USA, 150: 32-38. [ Links ]
34. Rzedowski, J. (1978). Vegetación de México. Ed. Limusa. México, D.F. 432 p. [ Links ]
35. Salvador, F.J. & S.J. Alvarez (2004) Flora y vegetación. In: Técnicas de muestreo para manejadores de recursos naturales. In: F. Bautista, H. Delfín, J. Palacio & M. Delgado (Eds.). Universidad Autónoma de México Consejo Nacional de Ciencia y Tecnología, Instituto Nacional de Ecología, 327 p. [ Links ]
36. Secretaría de Medio Ambiente y Recursos Naturales (2002). NORMA Oficial Mexicana NOM-059-ECOL-2001, Protección ambiental-Especies nativas de México de flora y fauna silvestres. Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio. Lista de especies en riesgo. Diario Oficial de la Federación. Segunda sección. 6 de marzo de 2002. México, D.F. 1-81. [ Links ]
37. Velazco-Macías, C.G., G. Alanis-Flores, M. Alvarado-Vázquez, L. Ramírez-Freire & R. Foroughbakhch-Pournavab (2011). Endemic flora from Nuevo León, Mexico and adjacent states. Journal of the Botanical Research Institute of Texas 5: 275-298. [ Links ]

