










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.82 no.2 Vicente López dic. 2013
ARTÍCULOS ORIGINALES
Population biology of Elaeodendron transvaalense Jacq. in the presence of harvesting
Efectos de la cosecha de la corteza en la biología de una población de Elaeodendron transvaalense Jacq.
Tshisikhawe MP & MW Van Rooyen
Department of Plant Science, University of Pretoria, 0002 Pretoria, South Africa.
Address Correspondence to: M Peter Tshisikhawe, Department of Botany, University of Venda, Private Bag X5050, Tohoyandou, 0950, South Africa. Tel.: +27 15 9628450; fax: +27 15 9628648, e-mail: tshisip@univen.ac.za, peter.tshisikhawe@univen.ac.za
Recibido / Received 22.XI.2012.
Aceptado / Accepted 12.I.2013.
Abstract. Elaeodendron transvaalense Jacq. is one of the medicinal plant species commonly used by people in the Venda region. The species is known to treat a variety of diseases. It is widely used and important to traditional healers. Because of this, it found its way into the muthi markets, and it is among the seven most commonly traded plant species in the Venda region. This study investigated the impact of bark harvesting on the population structure of this species. The study revealed that although the level of bark harvesting is high, the species is able to resist that stress since it is a fne-grained species. The population also showed the ability to regenerate as it exhibited an inverse J-shaped curve. The crown health status was generally good although some individuals, contributing 9% of the sample, had dead crowns, which are a cause for concern. A linear relationship was noticed between areas harvested and stem circumference, which is understandable considering the large surface area of harvestable bark on bigger individuals.
Keywords: Bark harvesting; Matrix modeling; Medicinal plants; Muthi markets; Population growth rate.
Resumen. Elaeodendron transvaalense Jacq. es una de las especies vegetales medicinales comúnmente utilizada por los habitantes en la región de Venda. La especie permite tratar una variedad de enfermedades. Es ampliamente usada e importante para los curanderos tradicionales. Debido a esto, llegó a los mercados muthi, y está entre las siete especies de plantas más comercializadas en la región de Venda. Este estudio investigó el impacto de la cosecha de la corteza en la estructura de la población de esta especie. El estudio mostró que aunque el nivel de cosecha es alto, la especie es capaz de resistir el estrés desde que es una especie de grano fino. La población también mostró la capacidad de regenerarse desde que exhibió una curva de J inversa. El estado de salud de la corona generalmente fue bueno aunque algunos individuos, que fueron el 9% de la muestra, tuvieron coronas muertas, lo que es un motivo de preocupación. Se observó una relación lineal entre la circunferencia del tallo y áreas cosechadas. Esto se entiende considerando la gran área de superficie de corteza cosechable en los individuos más grandes.
Palabras clave: Cosecha de corteza; Modelado de matrices; Plantas medicinales; Mercados muthi; Tasa de crecimiento de la población.
INTRODUCTION
Elaeodendron transvaalense Jacq., a species belonging to the Celastraceae family, is one of the medicinal plant species used very often by people around the Venda region located in the Limpopo province of South Africa. The species is among the seven medicinal plant species most commonly traded in the muthi markets around Venda (Tshisikhawe, 2002). Elaeodendron transvaalense is one of the medicinal plant species that is facing a serious threat of extinction as a result of bark overharvesting from their stems.
Elaeodendron transvaalense is used for a variety of diseases and hence its reference as "mukuvhazwivhi" by traditional healers, which literally translated means "sin-washer". The following are some of its medicinal uses (Mabogo, 1990; Van Wyk et al. 1997; Tshisikhawe, 2002; Steenkamp, 2003; Bessong et al., 2005; Samie et al., 2005): cleaning of stomach from any disorder, and treatment of (1) ulcers; (2) venereal diseases (STDs); (3) fungal infections; (4) piles and haemorrhoids in humans and domestic animals; and (5) dysmenorrhoea.
The root or stem bark decoction or infusion is taken orally in cupfuls three to four times a day (Mabogo, 1990; Van Wyk & Gericke, 2000; Tshikalange et al., 2008). The medicinal material is also prepared as a powder, and taken as tea or mixed with soft porridge. In 1988, the Chiang Mai Declaration noted that medicinal plants form the basis of medicines used by the majority of the population of most developing countries. As a result, the loss of certain medicinal plant species, and a reduced supply of other important plant species, would have a direct impact on human health and wellbeing (Bodeker, 1995).
Intense and frequent harvesting of bark from species with a high market demand often results in removal of bark from around the tree stems. Trees die thereafter, and the species becomes rare over time. Because of the demand of E. transvaalense as a medicine, it is important to assess the effects of bark harvesting on its population structure (Tshisikhawe, 2002). The population structure can be assessed by an analysis of the frequency distribution of stems across diameter classes (Condit et al., 1998; Lykke, 1998; Niklas et al., 2003b; Lawes et al., 2004). The stem size class distribution data can also be used to assess the potential sustainable use of the population (Everard et al., 1994; Everard et al., 1995; Obiri et al., 2002; Lawes & Obiri, 2003; Gaugris et al., 2007; Gaugris & Van Rooyen, 2007). Also, investigating the various aspects of the life cycle of a plant (e.g. age/size at flowering, seed output per size class) is crucial to gain an understanding of the dynamics of the population (Solbrig, 1980). Knowledge of the demographic variables of a population can be used in a more refned analyses of it population, such as matrix analysis (Caswell, 2001; Crone et al., 2011).
The objective of the current study was to investigate the impact of bark harvesting on a population of Elaeodendron transvaalense in the Venda region. Firstly, the population structure was examined, and the extent of bark harvesting was evaluated in terms of target size classes, and the effects on crown health and seed production. Secondly, a matrix analysis was performed to establish which size class contributed most to population growth rate, and should be selected for future conservation eforts. Tirdly, data were used to evaluate the potential for sustainable harvesting of the species by means of the species grain concept.
Spatial grain of regeneration has been used in landscape ecology to describe the coarseness in texture or granularity of spatial elements composing an area (Lawes & Obiri, 2003). Trough the spatial graining species were categorized as fine-, intermediate- or coarse-grained.
MATERIALS AND METHODS
The species. Elaeodendron transvaalense (Celastraceae) is a shrub or medium tree which can sometimes reach a height of 10 to 15 m. The species is widespread, although it is not common, at low altitudes in open woodlands. It grows from KwaZulu-Natal, Swaziland, Mpumalanga and through the northern parts of South Africa into Mozambique, Zimbabwe, Botswana and Zambia. The bark, which is used medicinally, is pale grey. Sometimes it has very small fissures, and breaks up into small blocks, especially in older individuals (Van Wyk, 1996; Palgrave, 1988). Leaves are simple, usually set at twig terminals, and are browsed by wildlife.
Study site. Data on population parameters were collected from an E. transvaalense population in the Tshirolwe area in the Venda region, Limpopo province. The Venda region is located in the northern part of the Limpopo province (23° 45' - 25° 15' S; 29° 50' - 31° 30' E). The Tshirolwe study area lies 38 km north of the town of Louis Trichardt, and 50 km west of the town of Thohoyandou in the Vhembe District Municipality of the Limpopo province. The study area is a communal area, which is accessible to the community of Tshirolwe without any restriction. The area is mostly used for grazing livestock and firewood collection.
According to Acocks (1988) the study area is part of the Northeastern Mountain Sourish Mixed Bushveld. However, Mucina & Rutherford (2006) classified it as Soutpansberg Mountain Bushveld. The vegetation type is regarded as 'Vulnerable'; approximately 21% of the area was cleared from woody vegetation, and dedicated to maize cultivation (Mucina & Rutherford, 2006). The area has a semi-arid climate, with the rainfall pattern infuenced by the Soutpansberg mountain range (Berger et al., 2003). The study area receives rainfall from October to March, and has a dry period extending from April to October. Mean annual rainfall in the Tshirolwe area (data from the closest weather station at Siloam, Weather Bureau 1998) is 390 mm. The study area rests on the gneisses of the Limpopo belt and Bandelierkop Complex (Berger et al., 2003). The Tshirolwe study area is situated within the Nzhelele-Formation, which is one of the seven units that constitute the Soutpansberg group of the volcano-sedimentary succession.
Sampling procedures. Eleven transects of 100 x 5 m were constructed to sample the required data. The coordinates of each transect were recorded using a 12 channel Garmin Geographic Positioning System (GPS) (Garmin International, Kansas City). A rope was used to delineate the transects during data collection. The following data were recorded on E. transvaalense individuals:
Stem circumference (in cm): measured with a measuring tape above the basal swelling.
Plant height (in m): measured with a measuring tape and/or graduated height rod.
Crown health: estimated using a 0 - 5 point scale as follows:
0 - no crown at all,
1 - severe crown damage,
2 - moderate crown damage,
3 - light crown damage,
4 - traces of crown damage,
5 - healthy crown.
Defoliation is widely used as an indicator for the vitality of forest trees and the degree of damage (Zierl, 2004). Crown health was assessed on a 0 - 5-point scale with 0 indicating 100% crown mortality and 5 indicating a healthy crown (Sunderland & Tako, 1999) and gave a good indication of overall tree health.
Bark removal area: breadth and width of the harvested area were measured with a tape measure.
Seed count: seeds were counted from one branch of a tree and an estimate for the tree was made. The estimates were considered minimal estimates of total seed production (Schwartz et al., 2002).
For the size class analysis, stem circumference measurements were classified into 13 size classes with 20 cm intervals, the first class being the 0-20 while the last one was the 241-260 class interval. Natural logarithmic transformations of the density of the size classes (D) [Condit et al., 1998; of the type ln (D+1)] were used to standardize the data (Niklas et al., 2003b) before calculating the least square linear regressions. The value of 1 was added as some size class bins were not represented (Lykke, 1998).
The mean circumference of the population, the "centroid", was calculated. A centroid skewed to the left of the midpoint of the size class distribution indicates a young and growing population, whereas one skewed to the right indicates an older, relatively undisturbed population (Niklas et al., 2003b). To estimate the harvesting pressure on an individual plant, a ratio of area harvested: stem circumference was calculated. This ratio was used to examine the relationship between harvesting pressure and crown health.
The subcanopy or canopy densities were calculated as the number of individuals ≤60 cm circumference or larger than 60 cm circumference, respectively, per unit surface area within the study area. The use of subcanopy and canopy densities, associated with frequency determinations, allow the grain of a species to be determined. The concept of species grain was developed for forests (Midgley et al., 1990); however, it has been successfully applied to woodlands by Gaugris et al., (2007) to establish which species could be harvested sustainably. The graphical model of Lawes & Obiri (2003) to determine species grain by plotting canopy density (X-axis) and subcanopy density (Y-axis) was used. The critical lower bounds for canopy and subcanopy densities of 10 and 30 individuals per ha of Lawes & Obiri (2003) were retained in this study.
A stage-class matrix analysis was performed using three stages, namely: seedlings; juvenile (non-fowering); and mature (fowering) plants. The transition matrix was derived using the age of transitions of the oldest seedling and vegetative stages. The matrix analysis was performed using the Matlab computer package version 7.0 which is regarded as the most appropriate package for these analyses (Caswell, 2001). An elasticity analysis was subsequently performed (Caswell, 2001; Norris & McCulloch, 2003).
RESULTS AND DISCUSSION
Population structure. The population status of E. transvaalense as reflected on the size-class distribution resembled the typical reverse J-shaped curve (Fig. 1) although some classes had few individuals of almost similar size-class ranges. Three ideal types of size-class distribution can be recognized for tree populations (Peters, 1996; Cunningham, 2001). The typical reverse J-shaped curve or negative exponential curve indicates continuous recruitment of young stems; the bellshaped curve indicates a lack of seedlings and young plants, and the straight horizontal line indicates relatively low numbers of seedlings and young plants. In a closed-canopy environment the reverse J-shaped curve is considered to indicate species which are tolerant to shade or competition, while the bell-shaped or straight line curves will indicate shade-intolerant or competition-intolerant species. A low number of seedlings may also be due to irregular recruitment opportunities.

Fig. 1. Size-class distribution of harvested and unharvested individuals in a population of Elaeodendron transvaalense sampled in 2004 at Tshirolwe, in the Venda region, Limpopo.
Fig. 1. Distribución de clases de tamaño de individuos cosechados y no cosechados en una población de Elaeodendron transvaalense muestreada en 2004 en Tshirolwe, en la región de Venda, Limpopo.
At a plant community level it has been established that the majority of the species increasingly resides in the smallest size-class (Niklas et al., 2003a; Guedje et al., 2007), and that in fact species richness is a size-class dependent phenomenon. Large size-class individuals in rare species were found in small numbers thereby attributing it to the rareness of the species. The fact that the E. transvaalense population sampled has showed few individuals in the large classes indicates that it is not abundant, and that it may become increasingly rare in the near future since it is unable to absorb the harvesting pressure.
A high abundance of individuals in smaller size classes, which leads to an inverse J-shaped size class distribution, is generally regarded as an indicator of adequate regeneration and population maintenance (Peters, 1996; Condit et al., 1998; Lykke 1998; Niklas et al., 2003a; Ganesan & Siddappa, 2004). The abundance of seedlings is therefore a manifestation of successful seed germination and establishment in the E. transvaalense population, although lack of adult size classes is a cause for concern. However, the position of the centroid at 49.12 cm, which was left-skewed in relation to the midpoint of the circumference distribution of 130 cm stem circumference, indicate the healthy status of the population, which is hindered by the lack of enough adult individuals in the greater stem circumference size classes.
The linear regression on the natural logarithm of the density in the size classes against the size class midpoint (Fig. 2) produced a significant linear regression (r² = 0.678; y = -0.014x + 4.279; p = 5.38 x 10-4). The slope and Y-axis intercept of this equation can be used in the future to compare other populations of E. transvaalense. It can also be used to monitor and compare the same Tshirolwe population over time to detect changes in population structure (Gaugris & Van Rooyen, 2010).
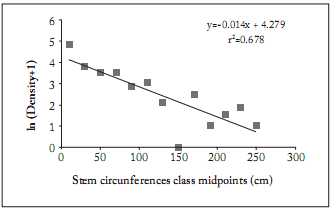
Fig. 2. The regression of ln (D + 1) versus stem circumference in a population of Elaeodendron transvaalense sampled in 2004 at Tshirolwe, in the Venda region, Limpopo.
Fig. 2. Regresión de ln (D + 1) versus circunferencia del tallo en una población de Elaeodendron transvaalense muestreada en 2004 en Tshirolwe, en la región de Venda, Limpopo.
Although long-term population monitoring data would be optimal to detect trends in population structure, Kohira & Ninomiya (2003) have indicated that there is merit in using the size-class distribution with a single-year data. Furthermore, a range of techniques have been devised to obtain as much information as possible from single surveys. The assessment of population structure with single-year data gives an essential head start for conservation eforts with a small amount of resources.
There was a significant positive correlation between plant height and stem circumference until an optimum height was achieved (Fig. 3; r2 = 0.4138; y = 0.0898x + 1.294; p= 6.99 x 10-21). Some individuals of stem circumference between 10 cm and 40 cm achieved a maximum height of more than 8 m.

Fig. 3. Positive linear relationship between stem circumference and plant height in a population of Elaeodendron transvaalense sampled in 2004 at Tshirolwe, in the Venda region, Limpopo.
Fig. 3. Relación lineal positiva entre la circunferencia del tallo y la altura de la planta en una población de Elaeodendron transvaalense muestreada en 2004 en Tshirolwe, en la región de Venda, Limpopo.
Harvesting. Forty eight percent of the E. transvaalense individuals sampled were not harvested (Table 1). Most of the unharvested individuals were seedlings. The large number of unharvested seedlings indicates that the population should potentially be able to recover if harvesting intensity is reduced, although it still needs to be monitored. In contrast, most of the larger size classes showed that 100% of the individuals had signs of harvesting.
Table 1. Extent of harvesting on Elaeodendron transvaalense individual trees.
Tabla 1. Grado de cosecha de la corteza en árboles individuales de Elaeodendron transvaalense.

Some individuals showed severe bark removal, with some of the individuals ending up dead due to harvesting pressure. Harvesting area increased with an increase in stem circumference (Table 1; Fig. 4; r2 = 0.6219, and y = 0.1437x - 0.1662). This is understandable because large trees have more available bark to harvest whereas seedlings do not have bark.
Overharvesting could be causing the absence of individuals either harvested or unharvested in the larger than 140 to 160 cm circumference size class in the studied E. transvaalense population. The three size classes most afected by the bark removal practices were the >180-200, >200-220, and >220-240 cm circumference classes (Fig. 4). These three class categories also constituted 30% of the individuals that showed 100% crown mortality.

Fig. 4. Relationship between the stem circumference classes and mean harvested area in a population of Elaeodendron transvaalense sampled in 2004 at Tshirolwe, in the Venda region, Limpopo.
Fig. 4. Relación entre las clases de circunferencia del tallo y el área promedio cosechada en una población de Elaeodendron transvaalense muestreada en 2004 en Tshirolwe, en la región de Venda, Limpopo.
Crown health. The crown health of the E. transvaalense population was generally not in a good state (Fig. 5). Ten percent (10%) of the E. transvaalense population crowns were found completely dead. Five percent (5%) had severe crown damage while 10% had moderate crown damage. Twentynine percent (29%) of the individuals sampled showed some traces of crown damage while 19% of individuals showed relatively healthy crowns. There was a negative, but not significant relationship, between the size of the individual and crown health (Fig. 6; r2 = 0.1464; y = -0.0096x + 3.7846; p = 0.10171.

Fig. 5. Crown health status of Elaeodendron transvaalense population in the Tshirolwe study area, Venda region, Limpopo, as determined by a survey in 2004. Crown health was assessed on a scale of 0-5 with 0 indicating 100% crown mortality and 5 indicating a healthy crown.
Fig. 5. Estado de salud de la corona en una población de Elaeodendron transvaalense determinada en un estudio en 2004 en el área de estudio Tshirolwe, región de Venda, Limpopo. La salud de la corona fue determinada en una escala de 0 a 5, con 0 indicando 100% de mortalidad de la corona y 5 indicando una corona sana.

Fig. 6. Stem circumference versus crown health status in a population of Elaeodendron transvaalense sampled in 2004 at Tshirolwe, in the Venda region, Limpopo.
Fig. 6. Circunferencia del tallo versus estado de salud de la corona en una población de Elaeodendron transvaalense muestreada en 2004 en Tshirolwe, en la región de Venda, Limpopo.
Although bark removal seemed to be the most likely factor contributing to the loss of crown health in the case of the Tshirolwe Elaeodendron transvaalense population, Zierl (2004) cautioned that it is important to devote more effort to the identification of other possible stress factors that may cause tree decline. In some cases, tree declines have been attributed to environmental stresses such as water availability or exceptionally high or low temperatures (Zierl, 2004).
In the Tshirolwe E. transvaalense population stress factors such as herbivory, trampling by livestock and wood harvesting for firewood were evident. The livestock observed in the study area were goats and cattle. In the population under study a number of seedlings were browsed. The effect of herbivory on seedling survival will have to be monitored in future studies. Fortunately, the collection of wood for firewood, which is very prominent in the area, is only done for E. transvaalense after the individuals have died from ring-barking.
Regeneration. The relationship between seed production and the size of the plant showed a very low coeficient of determination, indicating that plant size was a poor predictor of seed production (Fig. 7). In general, irrespective of the few individuals bearing seeds, seedling establishment seemed to be high due to large number of seedlings observed (Fig. 1).

Fig. 7. Stem circumference versus seed count per individual.
Fig. 7. Circunferencia del tallo versus número de semillas por individuo.
Regeneration in a forest or woodland is an indicator of the well-being of the forest (Murthy et al., 2002). Studies related to regeneration of a specific species or to the forest in general have always looked at factors responsible for degradation. In spite of the large number of seedlings, the seedlings of E. transvaalense were suppressed by herbivory. The effect of herbivory was largely counteracted by the ability of E. transvaalense to develop lignotubers which store starch and enable the seedling to develop quickly after being browsed. The lignotuber is a storage organ, which resprouts vigorously after stress effects like herbivores or fire. In the current study, resprouts were generally classified as seedlings. This was because it could be established that seedlings were resprouts from lignotubers. Seedling resprouting from lignotubers could give a false impression of the success of regeneration by seeds.
Stem growth rate. When analysing the stem growth increment of E. Transvaalense, a positive linear relationship was observed between the annual growth increment and the stem circumference size (Fig. 8; r2 = 0.8618; y = 0.0452x + 3.9228; p = 3.05 x 10-12). The mean stem diameter increment for the entire sample was 2.57 cm per annum. Although this growth rate appears to be high, it compares very well with growth rates of other woodland savanna species such as Garcinia livingstonei (2.6 cm/year), Sclerocarya birrea (1.33 cm/year) and Albizia versicolor (1.20 cm/year) (Gaugris et al., 2008).
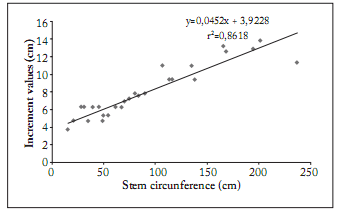
Fig. 8. Elaeodendron transvaalense annual stem circumference increment as a function of stem circumference at Tshirolwe, Venda region between 2004 and 2005.
Fig. 8. Incremento de la circunferencia anual del tallo de Elaeodendron transvaalense en función de la circunferencia del tallo en Tshirolwe, región de Venda, entre 2004 y 2005.
In many tree species, the growth rate of a tree changes with its life history (Kurokawa et al., 2003). Trees are expected to have their highest growth rate at middle size stages before growth is limited by the metabolic rate and reproduction. The mean stem circumference increment value of individuals in their vegetative stage was 5.74 cm (1.83 cm diameter increment) while it doubled to 10.56 cm (3.36 cm diameter increment) in the fowering stage (Fig. 8).
Population growth rate. After subjecting the matrix derived from E. transvaalense data through the lambda script on the Matlab programme, lambda was found to be 1.041. When using a constant transition matrix for multiplication, the prediction of future population size was generally of little relevance (Desmet et al., 1996; Morris & Doak, 2002). Because λ > 1, it showed that there should be an exponential increase in the population.
An elasticity analysis was performed to evaluate the relative importance of the population projection matrix cell entries and lower-level parameters on lambda. This analysis can be used to determine the stages of a species' life cycle that should be targeted for management actions (Link & Dohetry, 2002; Norris and McCullogh, 2003; Crone et al., 2011). Elasticity analysis showed that the highest elasticity value was in the cell, indicating the probability of a vegetative individual remaining in the vegetative stage, which had a value of 0.6420. This means that 64.2% of the influence on λ can be ascribed to this stage. It therefore indicates that for management purposes, it can be important to put more effort into protecting plants that are in the vegetative stage.
Species grain. The species grain concept provides information on whether a tree species can potentially sustain moderate harvesting levels or whether it may not survive such harvesting (Obiri et al., 2002). This approach provides a useful framework upon which to base operational bark harvesting rates.
The population of E. transvaalense under study could be classified as a fine-grained species (Fig. 9). It would therefore appear possible to harvest this species sustainably. In the case of E. transvaalense, individuals are not used for construction or other purposes, and bark-harvesting therefore represents the only form of harvesting. According to Obiri et al. (2002), the species grain theory suggests that fine-grained species should be able to withstand moderate levels of use. Therefore with proper harvesting techniques, E. transvaalense, which is only utilized for medicinal purposes, may survive such moderate harvesting.

Fig. 9. Species grain of the Elaeodendron transvaalense population of Tshirolwe from data collected in 2004.
Fig. 9. Grano de la especie en una población de Elaeodendron transvaalense en Tshrolwe de datos obtenidos en 2004.
CONCLUSIONS
The use of a size-class distribution analysis provided a practical field method for investigating the population structure of E. Transvaalense, and illustrated the response of the population to harvesting pressures. The data collected during this once-of survey can be used for monitoring changes in the population structure over time in the presence of harvesting.
The study has shown that the utilization of E. transvaalense by local people around Venda is currently very high. Despite the reasonable level of seedling establishment, the destruction rate of large trees is a point of concern. Bark harvesting for medicinal purposes is the major contributor to the loss of E. transvaalense individuals, since people only utilize it for firewood after it has died from debarking, and is dry. Cultivation intervention should be considered to reduce the stress experienced by E. transvaalense.
The matrix analysis allows one to answer a number of questions that cannot be answered by simple calculations. However, to improve the matrix analysis it is important to get repeated data on every individual in the population. Data should be recorded for a number of years in order to get a clear picture in terms of changes that occur. Data on mortality are especially needed to improve the parameterization of the cell entries in the transition matrix. This kind of information can also indicate the longevity of the individual.
ACKNOWLEDGEMENTS
We thank Mr Abraham Mukhadakhomu for assisting during data collection. Te assistance of Dr Vigdis Vandvik (from the Institute of Biology of the University of Bergen in Norway) and Dr Zuzana Munzbergova (from the Institute of Botany, Academy of Science of the Czech Republic) is gratefully acknowledged. This research project was supported by the National Research Foundation of South Africa.
REFERENCES
1. Acocks, J.P.H. (1988). Veld Types of South Africa. 3rd edition. Memoirs of the Botanical survey of South Africa. No. 57. 146 p. [ Links ]
2. Berger, K., J.E. Crafford, I. Gaigher, M.J. Gaigher, N. Hahn & I. Macdonald (2003). A first synthesis of the environmental, biological and cultural assets of the Soutpansberg. Leach printers, Louis Trichardt, South Africa. 66 p. [ Links ]
3. Bessong, P.O., C.L. Obi, M.L. Andreola, L.B. Rojas, L. Pousegu, E. Igumbor, J.J.M. Meyer, S. Quideau, & S. Litvak (2005). Evaluation of selected South African medicinal plants for inhibitory properties against human immunodefciency virus type 1 reverse transcriptase and integrase. Journal of Ethnopharmacology 99: 83-91. [ Links ]
4. Bodeker, G.C. (1995). Introduction. Medicinal plants for Conservation and Healthcare. Institute of Health Sciences, University of Oxford, Oxford, UK, pp 4. [ Links ]
5. Caswell, H. (2001). Matrix population models: Construction, Analysis and Interpretation. 2nd edition. Sinauer Associates, Inc. Publishers, Massachusetts, USA. 722 p. [ Links ]
6. Condit, R., R. Sukumar, S. Hubbel & R. Foster (1998). Predicting population trends from size distributions: a direct test in a tropical tree community. American Naturalist 152: 495-509. [ Links ]
7. Crone, E.E., E.S. Menges, M.M. Ellis, P. Bierzychudek, J. Ehrlen, T.N. Kaye, T.M. Knight, P. Lesica, W.F. Moris, G. Oostermeijer, P.F. Quintana-Ascencio, A. Stanley, T. Ticktin, T. Valverde & J.L. Williams (2011). How do plant ecologists use matrix population models? Ecology Letters 14: 1-8. [ Links ]
8. Cunningham, A.B. (2001). Applied ethnobotany: people, wild plant use and conservation. Earthscan Publication, London, UK. 300 p. [ Links ]
9. Desmet, P.G., C.M. Shackleton & E.R. Robinson (1996). The population dynamics and life-history attributes of a Pterocarpus angolensis DC. population in the Northern Province, South Africa. South African Journal of Botany 62:160-166. [ Links ]
10. Everard, D.A., J.J. Midgley & G.F. Van Wyk (1995). Dynamics of some forests in KwaZulu-Natal, South Africa, based on ordinations and size class distributions. South African Journal of Botany 61: 283-292. [ Links ]
11. Everard, D.A., G.F. Van Wyk & J.J. Midgley (1994). Disturbance and the diversity of forests in Natal, South Africa: lessons for their utilisation. Strelitzia 1: 275-285. [ Links ]
12. Ganesan, R. & S.R. Siddappa (2004). Regeneration of Amla, an important non-timber forest product from Southern India. Conservation and Society 2: 365-375. [ Links ]
13. Gaugris, J.Y. & M.W. Van Rooyen (2007). The structure and harvesting potential of the sand forest in Tshanini Game Reserve, South Africa. South African Journal of Botany 73: 611-622. [ Links ]
14. Gaugris, J.Y. & M.W. Van Rooyen (2010). The effect of herbivores and humans on the Sand Forest species of Maputaland, northern KwaZulu-Natal, South Africa Ecological Research 26: 365-376. [ Links ]
15. Gaugris, J.Y., M.W. Van Rooyen, & J.P. Bothma (2008). Growth rate of selected woody species in the northern Maputaland, KwaZuluNatal, South Africa. South African Journal of Botany 74: 85-92. [ Links ]
16. Gaugris, J.Y., C.A. Vasicek & M.W. Van Rooyen (2007). Selecting tree species for sustainable harvest and calculating their sustainable harvesting quota in Tshanini Conservation Area, Maputaland, South Africa. Ethnobotany Research and Applications 5: 373-389. [ Links ]
17. Guedje, N.M., P.A. Zuidema, H. During, B. Foahom & J. Lejoly (2007). Tree bark as a non-timber forest product: The effect of bark collection on population structure and dynamics of Garcinia lucida Vesque. Forest Ecology and Management 240: 1-12. [ Links ]
18. Kohira, M. & I. Ninomiya (2003). Detecting tree populations at risk for forest conservation management: using single-year vs. longterm inventory data. Forest Ecology and Management 174: 423-435. [ Links ]
19. Kurokawa, H., T. Yishida, T. Nakamura, J. Lai & T. Nakashizuka (2003). The age of tropical rain-forest canopy species, Borneo ironwood (Eusideroxylon zwageri), determined by 14C dating. Journal of Tropical Ecology 19: 1-7. [ Links ]
20. Lawes, M.J., H.A.C. Eeley, Shackleton, C.M. & B.G.S. Geach (2004). Indigenous forests and woodlands in South Africa: Policy, People and Practice. University of KwaZulu-Natal Press, Pietermaritzburg. [ Links ]
21. Lawes, M.J. & J.A.F. Obiri (2003). Using the spatial grain of regeneration to select harvestable tree species in subtropical forest. Forest Ecology and Management 184: 105-114. [ Links ]
22. Link, W.A. & P.F. Doherty (2002). Scaling in sensitivity analysis. Ecology 83: 3299-3305. [ Links ]
23. Lykke, A.M. (1998). Assessment of species composition change in saVanna vegetation by means of woody plants' size class distributions and local information. Biodiversity and Conservation 7: 1261-1275. [ Links ]
24. Mabogo, D.E.N. (1990). The ethnobotany of the Vhavenda. Master of Science dissertation, University of Pretoria, Pretoria. [ Links ]
25. Midgley, J., A. Seydack, D. Reynell & D. Mckelly (1990). Fine-grain pattern in southern Cape plateau forests. Journal of Vegetation Science 1: 539-546. [ Links ]
26. Morris, W.F. & D.F. Doak (2002). Quantitative conservation biology: theory and practice of population viability analysis. Sinauer Associates, Sunderland, Massachusetts. 480 p. [ Links ]
27. Mucina, L. & M.C. Rutherford (2006). The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute, Pretoria. 807 p. [ Links ]
28. Murthy, I.K., K.S. Murali, G.T. Hegde, P.R. Bhat & N.H. Ravindranath (2002). A comparative analysis of regeneration in natural forests and joint forest management plantations in UttaraKannanda district, Western Ghats. Current Science 83: 1358-1364. [ Links ]
29. Niklas, K.J., J.J. Midgley & R.H. Rand (2003a). Size-dependent species richness: trends within plant communities and across latitude. Ecology Letters 6: 631-636. [ Links ]
30. Niklas, K.J., J.J. Midgley & R.H. Rand (2003b). Tree size frequency distributions, plant density, age and community disturbance. Ecology Letters 6: 405-411. [ Links ]
31. Norris, K. & N. Mcculloch (2003). Demographic models and the management of endangered species: a case study of the critically endangered Seychelle magpie robin. Journal of Applied Ecology 40: 890-899. [ Links ]
32. Obiri, J.A.F., M.J. Lawes & M. Mukolwe (2002). The dynamics and sustainable use of high value tree species of the coastal Pondoland forests of the Eastern Cape Province, South Africa. Forest Ecology and Management 166: 131-148. [ Links ]
33. Palgrave, K.C. (1988). Trees of Southern Africa. 4th edition. Struik Publishers, Cape Town. 804 p. [ Links ]
34. Peters, C.M. (1996). The Ecology and Management of Non-Timber Forest Resources. World Bank Technical Paper No. 332. Washington, D.C., U.S.A. [ Links ]
35. Samie, A., C.L. Obi, P.O. Bessong & N. Lall (2005). Activity profiles of fourteen selected medicinal plants from rural Venda communities in South Africa against fifteen clinical bacterial species. African Journal of Biotechnology 4: 1443-1451. [ Links ]
36. Schwartz, M.W., T.M. Caro & T. Banda-Sakala (2002). Assessing the sustainability of harvest of Pterocarpus angolensis in Rukwa Region, Tanzania. Forest Ecology and Management 170: 259-269. [ Links ]
37. Solbrig, O.T. (1980). Demography and evolution in plant populations. Blackwell Scientific Publishers, California. 328 p. [ Links ]
38. Steenkamp, V. (2003). Traditional herbal remedies used by South African women for gynecological complaints. Journal of Ethnopharmacology 86:97-108. [ Links ]
39. Sunderland, T.C.H., & C.T. Tako (1999). The exploitation of Prunus africana on the island of Bioko, Equatorial Guinea. A report for People and Plants Initiatives, WWF-Germany and the IUCN/SSC Medicinal Plant Specialist Group. [ Links ]
40. Tshikalange, T.E., J.J.M. Meyer, N. Lall, E. Muñoz, R. Sancho, M. Van De Venter & V. Oosthuizen (2008). In vitro anti-HIV-1 properties of ethnobotanically selected South African plants used in the treatment of sexually transmitted diseases. Journal of Ethnopharmacology 119: 478-481. [ Links ]
41. Tshisikhawe, M.P. (2002). Trade of indigenous medicinal plants in the Northern Province, Venda region: their ethnobotanical importance and sustainable use. M.Sc. dissertation, University of Venda for Science and Technology, Tohoyandou. [ Links ]
42. Van Wyk, B.E. & N. Gericke (2000). People's plants. Briza publications, Pretoria, South Africa. 351 p. [ Links ]
43. Van Wyk, B.E., B. Van Oudtshoorn & N. Gericke (1997). Medicinal plants of South Africa. Briza Publications, Pretoria, South Africa. 336 p. [ Links ]
44. Van Wyk, P. (1996). Field guide to the trees of the Kruger National Park. Struik Publishers, Cape Town, South Africa. 272 p. [ Links ]
45. Weather Bureau (1998). Climate of South Africa: Climate statistics up to 1990. WB 42. Government Printer, Pretoria. [ Links ]
46. Zierl, B. (2004). A simulation study to analyze the relations between crown condition and drought in Switzerland. Forest Ecology and Management 188: 25-38. [ Links ]

