










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.83 no.1 Vicente López jun. 2014
ARTÍCULOS ORIGINALES
Pollen production pattern in the capitulum of the cultivated sunflower (Helianthus annuus L.)
Patrón de producción polínica en el capítulo del girasol cultivado (Helianthus annuus L.).
Astiz V1 & LF Hernández2,3
1 Laboratorio de Calidad Industrial de Granos. Chacra Experimental Integrada Barrow (convenio INTA-MAA), 7500, Tres Arroyos, Argentina.
2 Laboratorio de Morfología Vegetal. Departamento de Agronomía, Universidad Nacional del Sur. Bahía Blanca, 8000, Argentina
3 Comisión de Investigaciones Científicas de la Pcia. de Buenos Aires (CIC). La Plata, 1900, Argentina.
Address Correspondence to: Luis F. Hernández, tel: +54 0291 4566130, e-mail: lhernan@criba.edu.ar
Recibido / Received 15.X.2013.
Aceptado / Accepted 6.II.2014.
Abstract. The intra-plant pollen production in cultivated sunflower (Helianthus annuus L.) was quali- and quantitatively studied. Two modern self-pollinated, high oleic sunflower hybrids were grown during two consecutive seasons: 2009 in three planting dates, and 2010 in two planting dates. Two cross-pollinated varieties of sunflower, Hopi and Havasupai, were also studied to determine if an anomalous intracapitulum pollen gradient observed in preliminary studies could be a remnant character inherited from primitive genotypes. Pollen grains per flower (PGF), anther volume (AV), pollen grain volume (PGV) and pollen viability (PV) were measured in three capitulum sectors, external (ES), middle (MS) and internal (IS). At harvest, fully developed fruits (FDF) and incompletely developed fruits (IDF) per capitulum sector were counted. Based on seasonal averages of hybrids and planting dates, the capitulum IS showed the highest PGF number (40864 ± 6519) followed by the MS (35243 ± 5242) and the ES (28273 ± 3102). Anther length (AL) and width (AW) were higher in the IS (5.3 and 0.8 mm, respectively) compared with the ES (6.4 and 0.6 mm, respectively); ES and the MS had intermediate values (5.2 and 0.7 mm, and 4.9 and 0.6 mm, respectively). No significant differences (p=0.18) were observed in PGF per sector in capitula of Havasupai (average= 42679 ± 4107). On the other hand, Hopi fowers of the MS and IS produced a larger number of pollen grains (average=53412 ± 3553) than those of the ES (37050 ± 5947). PGV showed no significant differences in both hybrids, capitulum sectors and growing seasons (average= 22168 ± 1042 µm3). A positive relationship between AL and AW and the total number of pollen grains was also observed. Pollen viability in both growing seasons was greater than 97% in all cases. At harvest, in all cases, the higher number of FDF was observed in the ES and MS (598 and 456 respectively) and the number of IDF was higher in the IS (132). Given that intra-plant pollen production of both, pollen quantity and viability were not limiting, the origin of IDF in different regions of the capitulum could not be explained by the lack of pollen. The finding of an atypical positive centripetal gradient of pollen grains in the capitulum as compared with other racemose inflorescences is interpreted as an adaptive response to the time sequence of anthesis in this species to ensure a homogeneous pollination in the capitulum.
Keywords: Helianthus annuus L.; Pollen production; Pollen viability; Crop yield.
Abbreviations used: ES: capitulum external sector; MS: capitulum middle sector; IS: capitulum internal sector; AL: anther length; AW: anther width; IDF: incomplete developed fruits; FDF: fully developed fruits; PGV: pollen grain volume; PGF: pollen grains per foret; FPArea: area covered by pollinators on the capitulum surface.
Resumen. Se estudió cualitativamente y cuantitativamente la producción de polen intra-planta en el girasol cultivado (Helianthus annuus L.). Dos híbridos de girasol alto oleico se cultivaron durante dos temporadas consecutivas: 2009, en tres fechas de siembra, y 2010 en dos fechas de siembra. También se estudiaron dos variedades de girasol de polinización cruzada, Hopi y Havasupai, para determinar si un gradiente anómalo de polen intracapítulo, observado en estudios preliminares, podría ser un carácter remanente heredado de genotipos primitivos. Los granos de polen por flor (PGF), el volumen de la antera (AV) y del grano de polen (PGV) y la viabilidad del polen (PV) se analizaron en tres sectores del capítulo, externos (ES), medio (MS) e interno (IS). A la cosecha se contaron en cada sector del capítulo los frutos completamente desarrollados (FDF) y los frutos de desarrollo incompleto (IDF). Considerando los promedios estacionales de los híbridos y fechas de siembra, el sector interno (IS) del capítulo presentó el mayor número de PGF (40864 ± 6519), seguido por el MS (35243 ± 5242) y el ES (28273 ± 3102). El largo (AL) y ancho (AW) de la antera fueron mayores en el IS (5,3 y 0,8 mm, respectivamente) en comparación con el ES (6,4 y 0,6 mm, respectivamente); el ES y el MS presentaron valores intermedios (5,2 y 0,7 mm, y 4,9 y 0,6 mm, respectivamente). No se observaron diferencias significativas (p = 0,18) en el PGF por sector en los capítulos de Havasupai (media = 42679± 4107). Por otro lado, las flores del MS de Hopi produjeron un mayor número de granos de polen (promedio = 53412 ± 3553) que las del ES (37050 ± 5947). El PGV no mostró diferencias significativas en ambos híbridos, sectores del capítulo y estaciones de crecimiento (promedio= 22.168 ± 1.042 µm3). También se observó una relación positiva entre el AL, el AW y el número total de granos de polen. La viabilidad del polen en las dos temporadas de crecimiento fue superior al 97% en todos los casos. A la cosecha, en todos los casos, se observó el mayor número de FDF en el ES y MS (598 y 456, respectivamente) y el número de la IDF fue mayor en el IS (132). Dado que la producción de polen intra-planta, tanto la cantidad como la viabilidad, no fueron limitantes, el origen de los IDF en diferentes regiones del capítulo no podría explicarse por la falta de polen. El hallazgo de un gradiente de granos de polen positivo centrípeto y atípico en los capítulos en comparación con otras inflorescencias racimosas, se interpreta como una respuesta adaptativa a la secuencia temporal de la antesis en esta especie para asegurar una polinización homogénea.
Palabras clave: Helianthus annuus L.; Produccion de polen; Rendimiento; Viabilidad del polen.
Abreviaturas usadas: ES: sector externo del capítulo; MS: sector intermedio del capítulo; IS: sector interno del capítulo; AL: longitud de la antera; AW: ancho de la antera; IDF: frutos con desarrollo incompleto; FDF: frutos totalmente desarrollados; PGV: volumen del grano de polen; PGF: granos de polen por flor; FPArea: superficie del capítulo visitada por los polinizadores.
INTRODUCTION
The most common reasons for the origin of seedless or "incompletely developed" fruits or IDF (Lindstrom et al., 2004) in the sunflower capitulum are failures in fertilization, physiological or morphological defects in the ovary and embryo, and vascular deficiencies at the receptacle-ovary interphase (Hernández & Orioli, 1991; Alkio & Grimm, 2003; De Grandi-Hofman & Chambers, 2006).
Regarding pollination, even though the literature indicates that the sunflower is a self-incompatible insect-pollinated plant, to reduce the dependence on pollinators for seed production under field conditions, breeders have broken the selfincompatibility system and selected self-fertility in cultivated varieties (Roumet et al., 2012 and references therein). The selective pressures on self-compatible or self-incompatible varieties might have been very different, but always expected to have an optimum pollen presentation pattern producing very different "optimal solutions". Whereas in self-incompatible varieties, fruits cannot be produced without visits of pollinators and the patterns of pollen presentation could be crucial for maximum seed set, pollinators are not strictly required for seed set in selfcompatible varieties. Thus, the production of pollen could account by itself for successful pollination and eventually yield.
From a biological point of view, most primitive sunflower varieties and old varieties are self-incompatible and require cross-pollination for fruit setting. The choice of pollinators among those genotypes producing large amounts of efficient pollen could be of strategic importance for fruit production. This choice could be particularly important in modern sunflower hybrids.
It has been shown that inadequate pollination in sunflower is directly related to IDF (Hernández, 2008). A detailed study of visit path patterns of diurnal pollinators, mainly honeybees (Apis mellifera L.), showed a significant negative correlation (R2 = -0.61; p<0.05) between the areas covered by the insect foraging paths (FPArea) versus the total IDF counted in the capitulum (Hernández, 2008). A low coverage of insect visitation (FPArea = 0-30%) resulted in poor seed set (IDF ranging from 10 to 17%), while a significant insect visitation (FPArea = 60-90%) decreased the number of IDF by 5 to 9% (Hernández, 2008).
In sunflower, IDF causes important reductions in seed and oil production. Knowledge of pollen production, and determination of the reasons and the degree of the observed reduction in fecundity, are essential pre-requisites for sunflower management, improvement and breeding programs.
To date, knowledge about the amount of pollen grains produced by different florets of the sunflower inflorescence (capitulum) has received little attention, and only a few papers are available regarding this topic (Hernández, 2008; Astiz et al., 2011; Astiz & Hernández, 2013).
The aim of the present work was to determine quali- and quantitatively the intraplant pollen production and IDF generation in sunflower. High oleic hybrids were used, since these genotypes have recently been incorporated into the value chain of sunflower production, and have relevant importance for the future sunfower production in Argentina.
MATERIALS AND METHODS
Plant material. Two self-fertile high-oleic sunflower hybrids, Dekasol Oilplus (DKO) 3845 and DKO3945 (Monsanto Argentina S.A.I.C), were sown at the Agronomy Department -UNSur (Bahía Blanca, 38° 45' S; 62° 11' W). Experiments were conducted during two consecutive growing seasons: 2009 with three planting dates, and 2010 with two planting dates. Preliminary results obtained in our laboratory showed an anomalous intracapitulum pollen gradient, quite different from other racemose inflorescences. To help determine if these results could correspond to a remnant character inherited from primitive genotypes, two crosspollinated varieties of sunflower (i.e., Hopi and Havasupai: Heiser, 1951) were also grown during the second growing season.
After seedling emergence, plant density was adjusted to 6 plants/m2 by manual thinning. Each growing season, a completely randomized split-plot experimental design was used, with hybrid assigned to main plots and planting date to subplots. Three replicates were used per treatment. Each subplot had three rows 0.70 m apart and 6.0 m long.
Soil moisture was kept near field capacity by drip irrigation, and weeds were controlled manually. At the developmental stage V4 (Schneiter & Miller, 1981), plants were fertilized with 50.0 kg N/ha using NH4NO3. During the two-year study, mean air temperature (°C), solar radiation (mmol/m
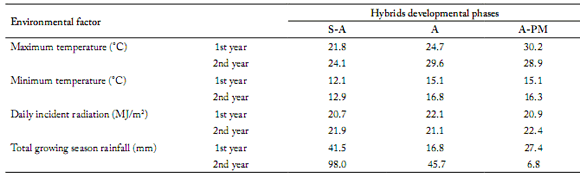
Table 1. Mean environmental data for three developmental crop phases for each year. Anthesis (A) and physiological maturity (PM) dates used for defning the mean crop stages represent the average for both hybrids in each experimental year. Crop phase A represents the 15-d interval centered on the mean date of full anthesis. Crop interval S-A is the time elapsed between sowing and 7 days before the mean date of full anthesis. A-PM is the interval between 7 days after full anthesis and the mean date of PM for both hybrids.
Tabla 1. Valores promedio de las variables ambientales durante las tres fases de desarrollo del cultivo para cada año. Las fechas de la antesis (A) y la madurez fisiológica (PM) utilizadas para definir estos estadios del cultivo corresponden a las fechas promediadas para ambos híbridos en cada año experimental. La fase A representa el intervalo de 15 días centrado en la fecha intermedia de plena floración. El intervalo S-A es el tiempo transcurrido entre la siembra y 7 días antes de la fecha media de plena foración. A-PM es el intervalo entre 7 días después que se completó la antesis y la fecha media de PM para ambos híbridos.
Pollen count and viability determination. Three plants per replicate were used for measurements. Quali- and quantitative pollen analysis were made in three capitulum sectors, external (ES), middle (MS) and internal (IS), each equal to 1/3 of the capitulum radius. Pollen grains per flower (PGF) were counted as follows: fve pre-anthesis flowers (i.e., 25 anthers) per capitulum sector, planting date, season, and genotype were removed from the inflorescences, and placed in 2.5 mL Eppendorf™ vials containing FAA fxative solution (Ruzin, 1999). The pollen was removed from the anthers at room temperature within 5-7 days from sampling. To facilitate the removal of pollen grains from the anthers, FAA was drained from each vial and replaced by 2 mL of a 1% aqueous solution Triton™ X-100 (Octyl Phenoxy polyethoxyethanol). Flowers were then crushed with a glass rod into the vial and stirred in a vertical vortex mixer to achieve a homogeneous suspension of pollen. A Neubauer hemocytometer was used to count the pollen grains, following Godini (1981). Two drops of the suspension containing the pollen grains were placed with a micropipette into each of the two main chambers of the hemocytometer. After placing the cover glass, pollen grains were counted in each of the nine small chambers forming the two main chambers. The number of pollen grains per anther was consequently calculated as the product of the observed pollen grains in each small chamber by a dilution factor, divided by the number of anthers in each vial (i.e., 25). Finally, the number of pollen grains per flower was calculated by multiplying the number of anthers per flower times the number of pollen grains per anthers.
Pollen viability (PV) was determined using Alexander's stain (Alexander, 1969) using Malachite green, which stains the cell wall cellulose, and Acid Fuchsine, which stains the protoplasm. Viable pollen grains stain dark, while non-viable ones remain light green. Pollen grain diameter was measured in samples of 60 grains taken from fowers on each capitulum sector. Pollen grain volume (PGV) was then calculated using the formula of the sphere. Using five fowers per each capitulum sector used for pollen count, anther length (AL) and width (AW) were also measured considering a transverse distance of both pollen sacs at the equatorial zone. Those values were used to estimate the volume of each pollen sac (VPS), calculated as the volume of an ellipsoid of revolution: PSV (mm3) = 4/3 . π . [(AW/2)2. AL]. Anther volume was then obtained as: AV (mm3) = PSV (mm3) . 4.
Pollen grain diameter and anther size measurements were made microscopically using an ocular micrometer.
Yield analysis. Six plants per replicate were randomly chosen and labeled at the reproductive stage R5.1 (Schneiter & Miller, 1981). The harvest was made with a fruit moisture content of 13 to 18%. Capitula were taken to the laboratory for analysis where they were split into three sectors, IS, MS and ES. The fruits were counted per sector and classified into two categories: fully developed fruits (FDF) and IDF (Lindström et al., 2004).
Statistical analysis. All data were statistically analyzed by a three-way analysis of variance (ANOVA) using the Infostat statistical software package v. 2010 (Di Rienzo et al., 2008). The least significant difference (LSD) test was used to separate differences between planting dates, hybrids, capitulum sectors and their interactions. In the case of cross-pollinated varieties one-way ANOVA was used to assess differences between capitulum sectors. Probabilities equal to or less than 0.05 were considered significant for main efects and interactions.
RESULTS
Pollen analysis. Table 2 shows pollen grain number per flower (PGF) for each capitulum sector in the different planting dates and for both hybrids and experimental years. In the first year, the internal sector showed the highest number of pollen grains per flower (30979 to 40448 PGF), while lowest values were shown in the external sector (21924 to 31625 PGF); the middle sector (26063 to 35396 PGF) showed intermediate values. The differences between the external and internal sectors were significant (p<0.05) in both hybrids for the three planting dates, except for the case of the third planting date when DKO3945 did not show significant differences (p=0.25), with values between 27167 and 30979 PGF (Table 2).
Table 2. Pollen grain number per flower for each capitulum sector (external, middle and internal) and three planting dates in the first year (2009) and two planting dates in the second year (2010), for each hybrid: Dekasol Oilplus 3845 (DKO3845) and Dekasol Oilplus 3945 (DKO3945). For each planting date and within capitulum sector, letters that differ within columns indicate values that are significantly different (p=0.05). (Data from Astiz, V. & L.F. Hernández, 2013. Phyton, Int J Exp Bot 82: 297-302).
Tabla 2. Polen Número de granos de polen por flor para cada sector del capítulo (externo, medio e interno) y tres fechas de siembra en el primer año (2009) y dos fechas de siembra en el segundo año (2010), para cada híbrido: Dekasol Oilplus 3845 (DKO3845) y Dekasol Oilplus 3945 (DKO3945). Para cada una de las fechas de siembra y dentro de cada sector del capítulo, las letras que difieren en las columnas indican los valores que son significativamente diferentes (p=0,05). (Datos de Astiz, V. & L.F. Hernández, 2013. Phyton, Int J Exp Bot 82: 297-302).

In the second year, pollen grain number per fower showed highly significant differences (p<0.01) among capitulum sectors, showing again the highest pollen production in the internal sector (43542 to 50250 PGF), and the lowest one in the external sector (27007 to 32000 PGF) (Table 2).
In both experimental years and in all planting dates, both hybrids showed a similar variation in the number of pollen grains per flower according to the sampled capitulum sector. Thus, the highest production of pollen was observed in flowers at the internal sector, followed by those at the middle sector, and finally the lowest production was observed in flowers at the external sector.
No significant differences (p=0.18) were observed in PGF per capitulum sector in capitula of Havasupai (Table 3). On the other hand, in Hopi, fowers of the MS and IS produced a larger number of pollen grains than those of the ES (Table 3).
Table 3. Anther length (AL, mm) and width (AW, mm); anther volume (AV, mm3) and number of pollen grains per fower (PGF) for each capitulum sector in both cross-pollinated varieties: Havasupai and Hopi.
Tabla 3. Longitud de la antera (AL, mm) y su ancho (AW, mm); volumen de la antera (AV, mm3) y número de granos de polen por for (PGF) para cada sector del capítulo, en las dos variedades de polinización cruzada: Havasupai y Hopi.
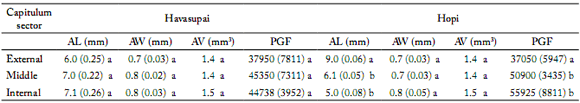
Values in parentheses = ± 1SD. Values followed by different letters are significantly different (p<0.05).
There was a significant interaction (p>0.05) between hybrids and seasons. Anther length, width and volume in both hybrids showed significant differences (p<0.05) among sectors, with the lowest values of the three variables in the ES and the highest ones in the IS (Table 4). In cross-pollinated varieties, only significant differences (p<0.05) were observed in the anther length of Hopi, with the highest values (9.0 mm and 6.1 mm) for ES and MS, respectively, and the lowest ones for IS (5.0 mm). Havasupai did not show significant differences (p>0.05) in any of the anther size parameters (Table 3). Pollen viability in the hybrids was over 97%, and significant differences were found between the external and the middle sectors (Table 4).
Table 4. Anther length (AL, mm) and width (AW, mm); anther volume (AV, mm3), pollen grain volume (PGV, µm3) and pollen viability (PV, %) for each capitulum sector. Pooled data across all plantings dates for both hybrids.
Tabla 4. Longitud de la antera (AL, mm) y su ancho (AW, mm); volumen de la antera (AV, mm3), volumen del grano de polen (PGV, µm3) y viabilidad del polen (PV, %) para cada sector del capítulo. Los datos presentados son la combinación de resultados para todas las fechas de siembra para los dos híbridos.

Values in parentheses = ± 1SD. Values followed by different letters are significantly different (p<0.05).
Anther length and width showed a direct relationship with pollen grain count (Figs. 1 and 2). The IS showed the largest anther length and width, followed by the MS and ES (Figs. 1 and 2). This was observed in both hybrids (Fig. 1) and in open pollinated varieties (Fig. 2), indicating a positive relationship between the anther dimensions and the number of pollen grains produced. No significant differences were observed (p=0.38) in pollen grain volume among the three capitulum sectors (Table 4). However, anther size parameters (AL and AW) showed significant differences, with higher values for the internal sector (25%) than those for the external one (Table 4). This was positively related to the number of pollen grains per fower produced in each sector, which was 27% higher in the fowers of the IS than in those of the ES.
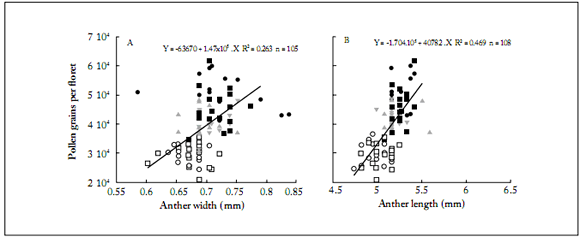
Fig. 1. Number of pollen grains per flower based on anther either width (A) or length (B) in each hybrid for each capitulum sector. DKO3845: capitulum sector external (○), middle (▲) and internal (); DKO3945: capitulum sector external (), middle (▼) and internal (■). In Fig. 1A arrows denote points that were considered outliers for the linear fit.
Fig. 1. Número de granos de polen por flor en función del ancho (A) o la longitud (B) de la antera en cada híbrido para cada sector del capítulo. DKO3845: sector del capítulo externo (), medio (▲) e interno (); DKO3945: sector del capítulo externo (○), medio (▼) e interno (■). En la Fig. 1A las fechas indican los puntos que se consideraron valores atípicos para el ajuste lineal.
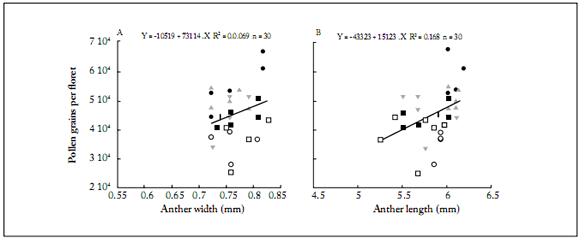
Fig. 2. Number of pollen grains per flower based on anther either width (A) or length (B) in each open pollination variety, Hopi and Havasupai, for each capitulum sector.
Havasupai: capitulum sector. external (⃞), middle (▼) and internal (■); Hopi: capitulum sector. external (○), middle (▲) and internal (•); A: Anther length (mm). B: Anther width (mm).
Fig. 2. Número de granos de polen por flor en función del ancho (A) o la longitud (B) de la antera en cada variedad de polinización abierta, Hopi y Havasupai, para cada sector del capítulo.
Havasupai: sector del capítulo externo (⃞), medio (▼) e interno (■); Hopi: sector del capítulo externo (○), medio (▲) e interno (•); A: Longitud de la antera longitud (mm). B: ancho de la antera (mm).
The open pollinated varieties showed more variability for AL and AW (Fig. 2). The magnitude of the average values of these variables was higher than that for the hybrids (Fig. 1). In both hybrids, data were distributed in a such a way that two clusters can be distinguished (Fig. 1). The first one corresponds to the highest AL, AW and PGF values, from the capitulum internal sector (black symbols in Fig. 1); the second one corresponds to the lowest AL, AW and PGF values from the external sector (white and grey symbols in Fig. 1). In the open pollinated varieties this pattern was not so clear, with a higher variation with respect to the hybrids (Fig. 2).
Yield analysis. In both hybrids and planting dates, the number of FDF showed significant differences (p<0.05) among capitulum sectors, with the highest value of FDF in the ES (598), followed by the MS (456) and the IS (358) (Table 5). Significant differences for IDF (p<0.05) were also found among the capitulum sectors, with a higher number of IDF in IS (132), than in MS (83) and ES (62); no significant differences were found between MS and ES (Table 5). Seed set was significantly different between the IS (74.8%) and the ES and MS (92.4% and 85.6%, respectively; Table 5).
Table 5. Number of fully developed fruits (FDF) and incompletely developed fruits (IDF), and seed set (%) for each capitulum sector, on average for hybrids and planting dates. Values followed by different letters are significantly different (p<0.05).
Tabla 5. Número de frutos completamente desarrollados (FDF) y frutos con desarrollo incompleto (FDI), y producción de semilla (%) para cada sector del capítulo, promediado para los híbridos y fechas de siembra. Los valores seguidos por letras diferentes son significativamente diferentes (p<0,05).

Values followed by diferent letters are significantly different (p<0.05).
DISCUSSION
The existence of a positive centripetal gradient of pollen grain number per fower in the sunflower capitula constitutes a remarkable finding. This positive centripetal gradient of intra-inflorescence pollen, unusual in many plant species, might be considered an adaptive response to the time sequence of anthesis in this species to ensure a homogeneous pollination of the capitulum. Thus the flowers of the SE, which open first, are more likely to have an allogamous pollination, while those that open later (SI) would be forced to autogamy because of temporal, positional and morphological disadvantages.
This intra-inflorescence fluctuation might be caused by its architecture (Buide, 2008). In species with racemose inflorescences the flowers that open first are more likely to receive external pollen, while those that open later (internal sector of the capitulum) will mostly receive pollen from the same plant (Ashman & Hitchens, 2000; Buide, 2004). Also, there is a clear tendency from the basal (flowers that open first) to the distal positions (flowers that open later): distal flowers are typically smaller and produce less and smaller ovules, fruits and seeds. However, they may have larger stigmata, and produce larger amount of pollen (Ashman & Hitchens, 2000).
These results have not been previously described for the Asteraceae in general or for the study genus in particular, since it is always assumed that the amount of pollen in inflorescences declines centripetally (Buide, 2008). However, currently available evidence indicates that this gradient can be generated as the result of gibberellin concentrations. It has been reported that a flowering GA3 concentration in the sunflower capitulum declines towards its center (Duca, 2006). Since GA3 arrests microsporogenesis (Duca et al., 2008; Kumar & Srivastava, 2009), it could act as a positional control for pollen grain production.
Gradients may also be due to an architectural effect, and to an anthesis temporal dynamic rather than to resource availability. Peripheral flowers (external 1/3 to 1/2 of the capitulum radius) are more likely to receive external pollen; flowers that open later (internal 1/3 of the capitulum radius) will mostly receive pollen of the same plant (Buide, 2004, 2008).
The primitive-domesticated Helianthus annuus L. lines, Hopi and Havasupai, showed the same quantitative intra-capitulum pattern of pollen grains. Could it be a genus character? In this sense, a positive centripetal gradient of the intra-inflorescence pollen production could be considered as an adaptive response to the time- sequence of the anthesis that will ensure a homogeneous pollination of the capitulum in the study species. Thus, flowers of the ES, that open first, are more likely to be allogamous while those that open later (IS) would be forced to autogamy.
In some sunflower lines there is evidence of a negative centripetal gradient of pollen viability. It has been observed that in some short statured sunflowers, pollen viability ranges from 60-80% in the peripheral fowers to less than 5% in the internal ones (Ramos et al., 2013).
According to the results shown here, we can infer that when viability is low, there is a need for pollen grains to compensate for the lack of capacity to provide male gametes during fertilization. In the sunflower capitulum the time to complete microsporogenesis is shorter in the anthers of central flowers than in peripheral ones (Dr. C. Sala, Nidera Arg., pers. comm.). Our results would add to the scenario presented in the sunflower capitulum upon completion of anthesis, in agreement with the pollen compensation mechanism previously mentioned
Figure 3 summarizes a global hypothesis of the pollen reduction pattern in the sunflower capitulum according to the four variables described above: pollen viability, gibberellin concentration, rate of microsporogenesis and pollen grain number per flower. Only the latter one has been proved in the present work.

Fig. 3. Schematic representation of a sunfower capitulum showing the proposed model for the response of the four variables described in this work, along the capitulum radius. These variables would explain the positive centripetal gradient of pollen grains per flower observed in the study.
Fig. 3. Representación esquemática del capítulo de girasol mostrando el modelo propuesto para la respuesta de las cuatro variables descriptas en este trabajo, a lo largo del radio de capítulo. Estas variables explicarían el gradiente positivo y centrípeto del número de granos de polen por flor observado en este estudio.
Finally, higher pollen ofer of the internal flowers, as a reward factor to pollinators, deserves a special attention. It is clear that when anthesis is taking place in the central region of the capitulum, the external flowers have already been fertilized and their ovaries are in active development. Therefore, due to a temporal and positional advantage, these flowers have already "won" the competition for resources when the central ones are opening. In this sense a higher pollen production in central flowers of the inflorescence could be an attractive mechanism for pollinators.
Other rewards of the flowers (nectar) have been found of high importance in determining the behavior of pollinators when visiting sunflowers (Nef & Simpson, 1990). However, nectar production related with foret position was not studied here. Nevertheless, it is known that central (distal) florets produce less nectar than flowers of the external (proximal) sector in racemose inflorescences (Devlin et al., 1987; Kudo et al., 2001). This is the result of competition between flowers and fruits for carbohydrates (Pleasants, 1983). Perhaps the change in pollen amount produced by the inner florets could possibly alter the foraging behavior of pollinators because of the reduced amount of nectar offered.
This intra-inflorescence variation in the amount of pollen per flower observed in the present study has been apparent in plants with specialized reproductive systems, mainly monoecious species, where male and female functions are in different flowers.
However, in inflorescences with hermaphroditic florets, considerable variation can occur in the energy investment, particularly focused to build a better display or attractive structures (petal size, nectar volume), biomass distribution and reproductive potential (Stephenson, 1981; Digle, 1995). Although the average energy investment for the production of pollen is higher than that required to produce ovules (190 Cal vs. 133 Cal, respectively; Smith & Evenson, 1978), in the case of sunflower, with uniovulate florets, this should not be an issue.
Regarding anther size, we found a good relationship between the length and the width of the anthers and the total number of pollen grains. Since the volume of each grain (PGV) does not fluctuate among capitulum sectors (Table 4), the gradient of pollen quantity observed may in part be attributed to the anther size. The fluctuation of these anther parameters in the primitive varieties of sunflower studied in the present work was higher than that observed in the hybrids (Fig. 2). This may be due to the fact that the processes of genetic improvement and selection in the sunflower have led to characteristics that determine that the anther dimension fluctuate less in the current hybrids than in wild species (Poverene et al., 2004). Nevertheless, the anther size/pollen grains relationship per capitulum sector in the primitive varieties was consistent with the response observed in the hybrids. Thereafter, anther size would be an inherited character of the genus that was not modified during genetic improvement in sunflower. Evidence in other species has revealed that a QTL for total anther length explained approximately 35% of the phenotypic variation for this trait (Georgiady et al., 2002; Conner, 2006; Brock et al., 2010).
CONCLUSIONS
It is not known whether failure of pollen germination on the stigma could occur or not. However, we can conclude that the formation of IDF in different regions of the capitulum would not depend on intraplant pollen offer, as both pollen offer and viability were not limiting. These results suggest that neither the amount nor the quality of the pollen would limit sunflower yield under the conditions of this study. The intra inflorescence positive centripetal gradient of pollen quantity observed in our investigation may respond to a legacy of the species as a strategy to invest resources in the sporophytic generation during the late stages of anthesis. Thus, the flowers of the ES, that open first, should have a greater chance of allogamous pollination, while those that open later (IS), would be forced to self-pollination due to temporal, positional and morphological disadvantages. This response could avoid photoassimilate supply competition between proximal and distal regions of the capitulum.
The identification of a positive centripetal gradient of pollen grain number per flower within the capitulum of both hybrids and the two cross-pollinated varieties disagrees with the quantitative description of pollen production in other species with racemose inflorescences. Results from the present study, regarding the fact that early-domesticated genotypes, Hopi and Havasapui, showed the same quantitative pollen pattern, suggests that this character would be a characteristic of the genus, and that it could be expressed as it was reported here under certain growth conditions.
ACKNOWLEDGMENTS
This work was funded by grants to LFH of the SGCyT-UNSur and the Comisión de Investigaciones Científicas (CIC), La Plata, Argentina. High oleic sunflower seed was kindly provided by Monsanto Seeds of Argentina. The provision of Hopi and Havasupai as well as valuable comments on the manuscript by Dr. C. Sala (Biotech. Dept., Nidera Argentina S.A.) are greatly appreciated. Results presented in this work were obtained while VA was holding a scholarship of the SGCyT-UNSur.
REFERENCES
1. Alexander, M.P. (1969). Differential staining of aborted and nonaborted pollen. Stain Tech 44: 117-122. [ Links ]
2. Alkio, A. & E. Grimm (2003). Vascular connections between the receptacle and empty achenes in sunfower (Helianthus annuus L.). Journal of Experimental Botany 54: 345-348. [ Links ]
3. Ashman, T. & M.S. Hitchens (2000). Dissecting the causes of variation in intra-inflorescence allocation in a sexuality polymorphic species, F. virginiana (Rosaceae). American Joournal of Botany 87: 197-204. [ Links ]
4. Astiz, V., L.A. Iriarte, A. Flemmer & L.F. Hernández (2011). Self-compatibility in modern hybrids of sunflower (Helianthus annuus L.). Fruit set in open and self-pollinated (bag isolated) plants grown in two different locations. Helia 54: 129-138. [ Links ]
5. Astiz, V. & L.F. Hernández (2013). Pollen production in sunflower (Helianthus annuus L.) is affected by air temperature and relative humidity during early reproductive growth. Phyton, International Journal of Experimental Botany 82: 297-302. [ Links ]
6. Brock, M.T., J.M. Dechaine, F.L. Iniguez-Luy, J.N. Maloof, J.R. Stinchcombe & C. Weinig (2010). Floral genetic architecture: An examination of QTL architecture underlying floral (co)variation across environments. Genetics 186: 1451-1465. [ Links ]
7. Buide, M.L. (2004). Intra-inflorescence variation in floral traits and reproductive success of the hermaprodite Silene acutifolia. Annals of Botany 94: 441-448. [ Links ]
8. Buide, M.L. (2008). Disentangling the causes of intrainflorescence variation in floral traits and fecundity in the hermaphrodite Silene acutifolia. American Journal of Botany 95: 490-497. [ Links ]
9. Conner, J.K. (2006). Ecological genetics of floral evolution. In: L.D. Harder and S.C.H. Barrett (eds.), pp. 260-277. Ecology and Evolution of Flowers. New York: Oxford University Press. [ Links ]
10. Connor, D.J. & A.J. Hall (1997). Sunflower physiology. In: A.A. Schneiter (ed.), pp. 113-182. Sunflower Technology and Production. Agronomy Monograph 35. American Society of Agronomy, Madison, USA. [ Links ]
11. DeGrandi-Hofman, G. & M. Chambers (2006). Effects of Honey Bee (Hymenoptera: Apidae) Foraging on seed set in self-fertile sunflowers (Helianthus Annuus L.). Environmental Entomology 35: 1103-1108. [ Links ]
12. Devlin, B., J. Brian Horton & A.G. Stephenson (1987). Patterns of nectar production of Lobelia cardinalis. American Midland Naturalist 117: 289-295. [ Links ]
13. Diggle, P.K. (1995). Architectural effects and the interpretation of patterns of fruit and seed development. Annual Review of Ecology, Evolution and Systematics 26: 531-552. [ Links ]
14. Di Rienzo, J.A., F. Casanoves, M.G. Balzarini, I. González, M. Tablada & C.W. Robledo (2008). InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
15. Duca, M. (2006). Spatial and temporal distribution of auxins and gibberellins in sunflowers (Helianthus annuus L.). Journal of Molecular Cell Biology 5: 43-49. [ Links ]
16. Duca, M., M. Port, M. Orozco-Cardenas & C. Lovatt (2008). Giberrellin-induced gene expression associated with cytoplasmic male sterility in sunflower. Biotechnology and Biotechnological Equipment 22: 691-698. [ Links ]
17. Georgiady, M.S., R.W. Whitkus & E.M. Lord (2002). Genetic analysis of traits distinguishing outcrossing and self-pollinations forms of currant tomato, Lycopersicon pimpinellifolium (Jusl.) Mill. Genetics 161: 333-344. [ Links ]
18. Godini, A. (1981). Counting pollen grains of some almond cultivars by means of an hemocytometer. Riv. Ortoforofruttic Ital 65: 173-178. [ Links ]
19. Heiser, C. (1951). The sunflower among the North American Indians. Proc Am Philos Soc 95: 432-448. [ Links ]
20. Hernández, L.F. & G.A. Orioli (1991). Role of different leaves of the sunflower (Helianthus annuus L.) plant during the grain filling period. Turrialba 41: 330-334. [ Links ]
21. Hernández, L.F. (2008). Visit path pattern of the honeybee (Apis mellifera L.) on the sunflower capitulum. Correspondence with the location of seedless and incompletely developed fruits. Helia 31: 1-16. [ Links ]
22. Kudo, G., T. Maeda & K. Narita (2001). Variations in floral sex allocation and reproductive success within inflorescences of Corydalis ambigua (Fumariaceae): pollination efficiency or resource limitation? Journal of Ecology 89: 48-56. [ Links ]
23. Kumar, G. & P. Srivastava (2009). Gibberellic acid-induced pollen mortality and abnormal microsporogenesis in Safflower. Cytologia 74:171-176. [ Links ]
24. Lindström, L.I., M.E. García & L.F. Hernández (2004). Morphology and distribution of incompletely developed fruits in sunflower (Helianthus annuus L.) capitula. XVI Int Sunf Conf Procs, Fargo, USA, ISA, pp. 333-337. [ Links ]
25. Nef, J.L. & B.B. Simpson (1990). The roles of phenology and reward structure in the pollination biology of wild sunflower (Helianthus annuus L., Asteraceae). Israel Journal of Botany 39: 197-216. [ Links ]
26. Pleasants, J.M. (1983). Nectar production patterns in Ipomopsis aggregata (Polemoniaceae). American Journal of Botany 70: 1468-1475. [ Links ]
27. Poverene, M., A. Carrera, S. Ureta & M. Cantamutto (2004). Wild Helianthus species and wild-sunfower hybridization in Argentina. Helia 27:133-142. [ Links ]
28. Ramos, M.L., E. Altieri, M. Bulos & C.A. Sala (2013). Phenotypic characterization, genetic mapping and candidate gene analysis of a source conferring reduced plant height in sunflower. Theoretical and Applied Genetics 126: 251-263. [ Links ]
29. Roumet, M., M.F. Ostrowski, J. David, C. Tollon & M.H. Muller (2012). Estimation of mating system parameters in an evolving gynodioecous population of cultivated sunflower (Helianthus annuus L.). Heredity 108: 366-374. [ Links ]
30. Ruzin, S.E. (1999). Plant Microtechnique and Microscopy. Oxford University Press, 322 p. [ Links ]
31. Schneiter, A.A. & J.F. Miller (1981). Description of Sunflower growth stages. Crop Science 21: 901-903. [ Links ]
32. Smith, C.A. & W.E. Evenson (1978). Energy distribution in reproductive structures of Amaryllis. American Journal of Botany 65: 714-716. [ Links ]
33 Stephenson, A.G. (1981). Flower and fruit abortion: proximate causes and ultimate functions. Annual Review of Ecology, Evolution and Systematics 12: 253-279. [ Links ]
