










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.83 no.2 Vicente López dic. 2014
ARTÍCULOS ORIGINALES
Biology of Dittrichia viscosa, a Mediterranean ruderal plant: a review
Biología de Dittrichia viscosa, una planta ruderal del Mediterráneo: revisión
Parolin P, M Ion Scotta, C Bresch
French National Institute for Agricultural Research (INRA)
Theoretical and Applied Ecology in Protected Environments and Agrosystems (TEAPEA)
1355, BP 167, 06903 Sophia Antipolis, France.
Address Correspondence to: Dr. Pia Parolin, e-mail: Pia.Parolin@sophia.inra.fr
Recibido / Received 1.VI.2013.
Aceptado / Accepted 12.VI.2013.
Abstract. Dittrichia viscosa is a common plant species in the Mediterranean region which is adapted to a wide range of environmental stresses. It is an important species that can be used for phytoremediation, as bioaccumulator or bioindicator, and it has the potential for being employed in integrated pest management in the Mediterranean ecosystem due to its entomophilous character.
Keywords: Allelopathy; Bioindicator; Biological control; Dittrichia; Inula; Habitat management; Host plant; Organic horticulture.
Resumen. Dittrichia viscosa es una especie vegetal común en la región mediterránea que está adaptada a un amplio rango de factores de estrés ambiental. Es una especie importante que puede utilizarse para fitorremediación, como bioacumulador o bioindicador, y tiene potencial para el empleo en el manejo integrado de plagas en el ecosistema mediterráneo debido a su carácter entomófilo.
Palabras clave: Alelopatía; Bioindicadores; Control biológico; Dittrichia; Inula; Manejo del hábitat; Planta hospedadora; Horticultura orgánica.
INTRODUCTION
Dittrichia viscosa L. (W. Greuther) (Asteraceae) is a herbaceous perennial Mediterranean plant species. It occurs in ruderal environments (e.g., along roads) and is an important natural food source for caterpillars of butterflies and moths; they feed on D. viscosa plants which provides plant sap for growth and development (Alomar et al., 2002; Perdikis et al., 2007, 2008). In its natural range, it is the host plant of efcient predators such as the mirid bugs of the genus Macrolophus or beneficial insects which control the olive fruit-fly (Fimiani & Digilio, 1993; Alomar et al., 2002; Broumas et al., 2002; Favas et al., 2003; Perdikis et al., 2007, 2008; Franco-Micán et al., 2010; Ingegno et al., 2011). In some Mediterranean agro-ecosystems it has been therefore employed to reduce the number of several pests in crops (Perdikis et al., 2007). Dittrichia viscosa probably has a high potential to conserve and augment the presence of predators. Its peculiarity to attract and feed the caterpillars of predatory insects, and because of its wide range of tolerance to environmental stresses, makes it potentially suitable for biological control in the Mediterranean region. Since this plant species is highly resistant to adverse conditions and occurs in degraded environments, it has characteristics which enable it to become an invasive weed (DPIPWE, 2011; Sinden et al., 2004). Because of this, its use for biological control must be restricted to its native range: the Mediterranean. This whole region, since thousands of years, has been traditionally used for agriculture (Fig. 1).

Fig. 1. Dittrichia viscosa. (A) A panicle showing its capitules. (B) Capitule detail. (C) View of the whole plant. (D) Hairy leaves. (E) Fruiting plants.
Fig. 1. Dittrichia viscosa. (A) Se observan los capítulos en una panoja. (B) Detalle del capítulo. (C) Vista de la planta completa. (D) Hojas pilosas. (E) Plantas en fructificación.
The aim of the present paper is to review the ecological, physiological and phenological characteristics of D. viscosa.
Taxonomy of D. viscosa
Dittrichia viscosa L. (W. Greuther) is an evergreen perennial, flowering plant species of the family of Asteraceae (Anderberg, 1991; Anderberg et al., 2005; Pelser et al., 2010). The so-called False Yellowhead or Woody Fleabane formerly belonged to the genus Inula (Brullo & De Marco, 2000). Two subspecies and one variety have been classified: D. revoluta (synonym: Inula viscosa subsp. revoluta = D. viscosa subsp. revoluta) that occurs only in SW Portugal, and D. orientalis (syn.: I. viscosa var. angustifolia) that occurs in the Eastern Mediterranean (Brullo & De Marco, 2000); besides D. viscosa, other species such as D. graveolens and D. maritima are common in the Mediterranean region.
Distribution, habitat requirements and invasiveness
Dittrichia viscosa is common throughout the Mediterranean Basin. Its native range includes the coasts of southern Europe [France (e.g., Fig. 2), Spain, Greece, Italy, Bulgaria] and Turkey, the Middle East (Israel, Jordan and Syria) as well as northern Africa (Algeria, Egypt, Libya).

Fig. 2. Dittrichia viscosa in Southern France. A, October (autumn): plants which were not cut for several years (in flower at the background), and smaller plants which were cut in July (summer). B, January (winter): taller plants without green leaves (at right of the background), small plants full of green leaves.
Fig. 2. Dittrichia viscosa en el Sur de Francia. A, Octubre (otoño): plantas que no fueron cortadas durante varios años (en floración en el fondo), y plantas más pequeñas que fueron cortadas en Julio (verano). B. Enero (invierno): plantas más altas sin hojas verdes (a la derecha en el fondo), plantas pequeñas completamente con hojas verdes.
Dittrichia viscosa has the potential to be a significant environmental weed due to its very efficient seed production and dispersal, and it is well adapted to disturbed open situations and dry ecosystems (Groves, 1991). In fact, it is established in the Azores, Belgium, and Great Britain (DAISIE, 2012). It is now common also in other regions of Europe and North America. In Western Australia it is considered a serious en- vironmental weed marked for eradication and prohibited import to Australia (Sinden et al., 2004, DPIPWE, 2011). The economic impact of D. viscosa is mostly relevant to natural areas and roadsides, where its establishment would lead to greater cost burdens associated with its control and removal (Scott et al., 2008).
Dittrichia viscosa is a hemicryptophyte with a remarkable pioneer character found in a variety of habitats in the Mediterranean climate without a clear microhabitat restriction (Scott et al., 2008). Typically, the climate exhibits long, dry, hot, high-irradiance summers (Di Castri, 1973). Most plants native to this climate, such as D. viscosa, have adaptations to withstand the limitations imposed. There are winter annuals which germinate with the onset of autumn rains and complete their life cycle early in the next summer (Mooney, 1987).
Dittrichia viscosa may be considered a ruderal plant species due to its abundance in areas that have been modified and altered by anthropic activities (Murciego et al., 2007). The typical habitats of D. viscosa are dry riverbeds and abandoned fields, roadsides, walking trails, or even urban areas. It occurs on clay and sandy soils, rocky coasts or in natural swamps and other wetlands (Warlop et al., 2010), and it is light demanding. It is not entirely dependent on fre or mechanical disturbance to be competitive (Sinden et al., 2004).
Dittrichia viscosa also occurs on ultramafic sites where it responds well to high nickel and magnesium concentrations (Ater et al., 2000). In a study on nitrogen-driven biodiversity, D. viscosa was the only species which responded significantly to increased N, i.e., its cover decreased in control plots, but increased in fertilized plots (Dias et al., 2011). For this reason, it could be used as an indicator of N enrichment in plantations covered by Mediterranean maquis.
Biology of D. viscosa
Morphological and anatomical characteristics. Dittrichia viscosa is a perennial herb, classifed as nanophanerophyte (Conesa et al., 2011) or sometimes as chamaephyte with a woody base (Weker & Fahn, 1981).
The roots can be quite substantial, even in small plants. Specific root length is approximately 116 cm/g and the ratio of below-ground to above-ground biomass is 0.24 (Stavrianakou et al., 2006); however, there is no tuber at the base.
It has upright stems, with bifurcation starting low on the stem. Branches are many, spreading, irregularly angled. The canopy is very dense. It reaches 150 cm of height (Warlop et al., 2010). It is covered by glandular hairs.
The leaves are sticky, hence the name "viscosa". The shape of the pubescent leaves is elongated, the borders serrulate to denticulate. Mature leaves reach an average size of 7 x 1.8 cm, and average leaf thickness is 197µm (Stavrianakou et al., 2006). Specific leaf area is 502 cm2/g on average; total leaf number per plant averages 130.9, and total leaf area per plant comprises 200 cm2 (Stavrianakou et al., 2006).
The upper leaves are clearly stem-clasping through their base. The leaves are close to and under the flower. The involucres with bracts are unequal, external and outwardly viscous. The yellow capitula (=flowers) are numerous and grouped as a composite cluster. Dittrichia viscosa has long inflorescences, and the corolla of its yellow flowers is 2 cm in diameter, the ligules distinctly exceed the involucre.
The fruit is a 2 mm long achene with a hairy pappus with 15 bristles.
Dittrichia viscosa is well known for its strong odor (Wang et al., 2004). The young stems and leaves are covered with glandular hairs which exude a sticky, strong-smelling oil. The leaves possess long protective hairs, numerous glandular hairs and stomatal complexes on both the abaxial and adaxial surfaces (Nikolakaki & Christodoulakis, 2004). Glandular hair density, on average, is 14 hairs/mm on the abaxial leaf surface, and 11 hairs/mm on the adaxial surface (Stavrianakou et al., 2006). The protective hairs have complex structures, and secrete throughout the life span of the leaf (Werker & Fahn, 1981; Omezzine et al., 2011). The glandular hairs secrete lipids, polysaccharides and protein (Werker & Fahn, 1981).
Phenology. Dittrichia viscosa starts sprouting at the end of March or April (early European spring; Fig. 3). The main growth is completed by May, and the starch accumulation in the whole plant ends up in June (Meletiou-Christou et al., 1998; Scott et al., 2008; Parolin et al. 2013a). There is a secondary growth period in October/November (European autumn) and the leaves fall in December (winter) (Meletiou-Christou et al., 1998) (Fig. 2).
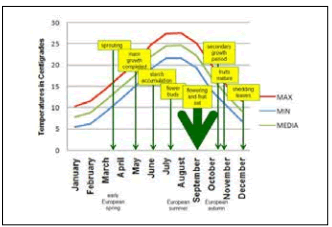
Fig. 3. Phenology of D. viscosa, and mean monthly minimum, average and maximum temperatures along the annual cycle.
Fig. 3. Fenología de D. viscosa, y temperaturas mínima, promedio y máxima mensuales promedio durante el ciclo anual.
At the end of July (European summer), flower buds are present. From September, both flowering and fruiting occur. Its prolonged flowering lasts for a long period, towards the end of the warm season, from August to October. Its fruits are mature in October and November and they are wind dispersed (Parolin et al., 2013a).
Regeneration and cultivation
Flowers are hermaphroditic and insect pollinated. Reproduction occurs via seeds which are commonly dispersed by wind and water. They can also be spread during soil movement (DPIPWE, 2011). Deep dormancy has been demonstrated in the laboratory which is broken by a lack of light (Sinden et al., 2004), but the longevity and viability of seeds is not known. Dispersal occurs when the pappus catches the wind or allows the plant to float in running water. Since it is a prolific seed producer it represents a threat as an invasive species (e.g., in the higher rainfall regions of southern Australia: Sinden et al., 2004).
In its native range, D. viscosa germinates mostly after autumn rainfalls but if adequate moisture is available it may occur at any time (Perez-Fernandez et al., 2006). Germination appears to be favored by ground disturbance and fre. Seed germination occurs from 15 to 30 °C with maximum germination at 25 °C (Doussi & Tanos, 1997). Temperaturegrowth experiments indicated that the ideal temperature range is from 10 to 24 °C (Scott et al., 2008).
Dittrichia viscosa shows a great tolerance to drought, germinating even at water potentials of approximately -0.5 MPa (Pérez-Fernández et al., 2006). In an experiment, germination rate increased in response to low osmotic potentials (Pérez-Fernández et al., 2006). The presence of sticky and mucilaginous villi may help to reduce the superficial tension between the ground and the seed, and therefore they are able to retain a greater amount of water in a smaller period of time. Thus, soil water becomes almost immediately available, enhancing germination. However, after removing the appendices, the seeds of D. viscosa reached high levels of germination under various osmotic potentials (Pérez-Fernández et al., 2006). Without the anatomical structures of water retention, seed germination was highest (64.5%) due to the fact that the villi and the seeds are impregnated with a sticky and hydrophobic substance. Thus, when eliminating the villi, any amount of water can reach the seed, activating its germination, even at extremely low values of water potential (Pérez-Fernández et al., 2006).
Being a ruderal plant species, D. viscosa is expected to germinate easily and to have little requirements for growth (Warlop et al., 2010). We sowed seeds of D. viscosa in a greenhouse in spring (19/04/2012) in a mixture of 1/3 perlite and 2/3 loamy soil, at an average temperature of 25 ± 2 °C and RH of 70 ± 10%. The seeds germinated after 8-10 days, but their growth was slow; four weeks after sowing the plants attained 15-20 cm height.
Dittrichia viscosa can be regenerated in several ways. One is by seeds which can be collected at the end of October. High quantities of seeds are then available and easy to handle; however, germination rates are poor and the delay until flowers are present is long (Warlop et al., 2010). Seeds can be sown in autumn, at the edges of hedges (i.e., saving the middle of fields), at a distance of 45-50 cm from one another and lightly pressed into the soil with a roll. They will germinate the following year without further interventions (ITAB, 2005). It is also possible to plant D. viscosa in early autumn (September-October, to avoid frosts); watering is necessary for the first year in the periods of absence of rain.
A second, faster method is through cuttings planted in pots.
A third possibility is the transplantation of whole plants: transplanting 10 cm height plants in February-March is useful to ensure the establishment of D. viscosa (CIVAM, 2011), and it is quickly efficient because plants have flowers from the first year on, but the extraction of the root system for transplanting is not easy.
When used to control olive fruit-flies, D. viscosa should be planted close to olive trees so that interactions of the arthropods on D. viscosa and olive trees may take place. However, they should not be planted too close to avoid (1) hindering labor or (2) enhancing competition between plants.
If the plant is cut prior to fowering (e.g., in September), sprouting is favored and D. viscosa will have leaves throughout the winter (FEDER, 1997; Alomar & Albajes, 2003).
Its growth must be controlled by regularly cutting back part of the plants, because it can become very large and inhibit agricultural practices (Warlop et al., 2010). In May-June, the plants should be cut back to a height of 30-40 cm (CIVAM, 2011).
Metabolism and secondary compounds
Dittrichia viscosa performs annual fuctuations in the content of soluble sugars, starch, total lipids and nitrogen (Meletiou-Christou et al., 1998). Dittrichia viscosa is closely dependent on arbuscular mycorrhiza for growth, and fungi afect the water content in its shoots (Roldan-Fajardo, 1994). Under water stress, growth is suppressed but glandular hair density is not affected (Karageorgou et al., 2002). The concentration of leaf exudate phenolics increases under water stress and decreases under additional nutrients, predicting environmental effects on carbon-based defensive secondary metabolites (Karageorgou et al., 2002). It is noteworthy that water stress almost doubled the concentration of epicuticular material (Karageorgou et al., 2002).
Its aboveground biomass accumulation was not affected under shade conditions, yet the total leaf area was considerably increased at the expense of dry mass invested on a surface area basis (Karageorgou et al., 2002). Plants grown under shade exhibited lower root dry mass and higher total leaf area (Karageorgou et al., 2002).
Soluble sugars increase in all plant parts during the growth period in summer and the following drought period, and decline thereafter, which is typical for a hemicryptophyte accumulating reserves (Meletiou-Christou et al., 1998). Fluctuations of storage compounds contribute to the adaptive mechanisms produced under stress conditions (Larcher & Tomaser-Tin, 1988). There is a summer peak of leaf soluble sugars in August. Thereafter, concentrations decline due to leaf senescence. Starch content, which is easily converted to soluble carbohydrates under stress conditions, is depleted earlier, influenced by climatic conditions. Starch is accumulated in all plant parts during the main growth period from March to June, in agreement with the postulate that in Mediterranean areas a maximum in carbon input occurs during spring (Tenhunen et al., 1984). Sugars are produced in excess until June and condensed to starch (Meletiou-Christou et al., 1998); thereafter, a continuous decrease is observed until the dehiscence of the leaves (Meletiou-Christou et al., 1998).
The decrease of starch during summer is accompanied by a depletion of total lipids while osmotically active soluble sugars increase (Meletiou-Christou et al., 1998). Hence, less water is wasted during the drought stress, at the expense of reserves stored in leaves and roots (Amundson et al., 1995).
Lipids are accumulated in young, expanding leaves from April onwards in the growth period and depleted during the summer drought (Meletiou-Christou et al., 1998). The maximum concentration of lipids (45 mg/g DW) is reached in June and decreases in senescing leaves until their dehiscence in November (Meletiou-Christou et al., 1998).
In D. viscosa, the minimum foliar nitrogen concentrations are recorded in autumn and the maximum values in winter and spring. Total nitrogen levels are considerably increased by additional nutrients and shade, but remain unaffected by water stress (Karageorgou et al., 2002). In another study, Meletiou-Christou et al. (1998) argue that lipid concentrations ranged from 1.1 to 3.1 kJ/g DW; an increase was observed in young expanding leaves and the maximum value was recorded in June. During the following months, the energy content of storage substances decreased continuously until leaf senescence and defoliation. Total-N and protein-N concentrations declined in young, expanding leaves, from March until June; the relationships between total-N and soluble sugars, and between total-N and starch were negative. In early spring, the concentration of soluble-N increased in all parts; new leaves attained high proportions of soluble-N. An increase of soluble-N was observed in young, expanding leaves, and a decrease in senescing leaves before their dehiscence. In the roots soluble-N increased in autumn in parallel with leaf senescence, and decreased during the growth of new leaves. The decrease of nitrogen concentrations during the growth period in the leaves of Mediterranean shrubs has been considered a result of a dilution effect (Field & Mooney, 1983); from August until November, during leaf senescence, foliage exports carbon, and the negative relationships between nitrogen and starch, and between nitrogen and soluble sugars result primarily from the decrease of carbohydrate concentrations.
Dittrichia viscosa responded significantly to increased N, i.e., it decreased in control plots, but it increased in fertilized plots (Dias et al., 2011). A sink-storage system may be functioning between stems and roots which show annual fuctuations of the energy content of storage substances (Meletiou-Christou et al., 1998).
Dittrichia viscosa is rich in epicuticular flavonoids, and 22 flavonoid aglycone structures were found in exudates, including three flavanones (Wollenweber et al., 1991). The flavonoids' spectral profile shows maxima in the harmful UV-B (280-320 nm) region, with a peak at 290 nm (Stephanou & Manetas, 1997).
Dittrichia viscosa has a (1) low specific plant weight, (2) high relative water content, (3) high content of chlorophyll, and (4) high ratio of chlorophyll:nitrogen (FEDER, 1997).
Photosynthesis and related parameters of D. viscosa indicate chlorophyll concentrations of approximately 23.5 µg/cm2; chlorophyll fluorescence Fv/Fm ratios of 0.83, and a photosynthetic capacity in healthy plants of approximately 14.3 µmol O2/m2/s (Stavrianakou et al., 2006). Boron deficiency caused a dramatic restriction of growth, but did not have any negative effect on parameters related to photosynthesis (Stavrianakou et al., 2006).
The species is rich in secondary compounds such as flavonoids, sesquiterpenes and essential oils (Shtacher & Kashman, 1971; Grande et al., 1985; Wollenweber et al., 1991; Perez-Alonso et al., 1996; Abu Zargaet al., 1998; De Laurentis et al., 2002; Nikolakaki & Christodoulakis, 2004; Miguel et al., 2008). More than 80 volatile oils were identifed in its leaves and flowers (Perez-Alonso et al., 1996; De Laurentis et al., 2002; Miguel et al. 2008). The major components of the oil were 3-methoxy cuminyl isobutyrate (12%), a-cadinol (6.3%), eudesm-6-en-4 a -ol (4.8%) and d-cadinene (4.6%) (Miguel et al., 2008). There are marked diferences between the essential oil obtained from the aerial parts and that isolated from the roots (Perez-Alonso et al., 1996). In the roots, the major volatile constituents were 3-methoxy-p-cymene-7-yl isobutyrate and isovalerate and neryl isovalerate (Perez-Alonso et al., 1996).
The aerial parts of D. viscosa contain triterpenoids as free alcohols, acetates or fatty esters (Grande et al., 1992). Leaf extracts contain oxygenated sesquiterpene compounds to a greater extent (De Laurentis et al., 2002), and flavonoids responsible for antioxidant effects (Schinella et al., 2002). Flower extracts contain especially large amounts of monoterpenes, sesquitepene hydrocarbons, oxygenated aromatic compounds and fatty acid esters (De Laurentis et al., 2002).
The amount and the allelopathic potential of the watersoluble leaf epicuticular exudates increase during the dry, hot, and sunny summer (Stephanou & Manetas, 1997). Besides the seasonal variations, geographic variations were also found. The percentage of monoterpene and sesquiterpene compounds depend on the occurrence of the plants in mountainous or seashore environments (Chamaco et al., 2000; De Laurentis et al., 2002).
Pharmaceutical applications and use
Innumerate publications state the medicinal and pharmacological potential of D. viscosa and the most important applications of its extracts which have balsamic, healing, antipyretic, antidiabetic, antiphlogistic, anti-viral, anti-fungal, anti-bacterial, and antiseptic properties (Lauro & Rolih, 1990; Barrero et al., 2008). Dittrichia viscosa has been used in folk medicine and is of major interest for many pharmaceutical industries (Perez-Alonso et al. 1996; Schinella et al., 2002; Nikolakaki & Christodoulakis, 2004; Celik & Aslantürk, 2010). From ancient times it has been used to treat wounds and injuries, bruises, and intestinal disorders (Perez-Alonso et al., 1996; De Laurentis et al., 2002). Extracts from D. viscosa display anti-inflammatory activity and free radical-generating systems which protect against enzymatic and non-enzymatic lipid peroxidation in model membranes, and show scavenging property on the superoxide radical (Schinella et al., 2002). It has antiulcerogenic and abortive effects (Al-Dissi et al., 2001; Nikolakaki & Christodoulakis, 2004). Skin contact with D. viscosa can cause allergic reactions in humans (DPIPWE, 2011) but it can also cure skin diseases (e.g., edemas and chronic dermatitis) due to the acute anti-inflammatory activity of the sesquiterpenoids ilicic acid and inuviscolide, isolated from D. viscosa (Máñez et al., 1999; Hernández et al., 2001). The antimycotic activity of the plant extract against dermatophytes is due to the decline in chitin content, a very important constituent of fungal cell wall (Mamoci et al., 2011). Celik & Aslantürk (2010) state that despite the beneficial effects of D. viscosa as a medicinal herb, it can cause serious problems and damage on cells when is used improperly.
Borneol and bornyl and isobornyl acetates are valuable ingredients in perfumery and pharmacy (Perez-Alonso et al., 1996). Therefore, the essential oil of D. viscosa may be suited to be an alternative source of these components.
Plant species resistance to stress
The secreted substances play a defensive role of leaf surfaces against microorganisms (Karageorgou et al., 2002), and have a strong absorbance in the ultraviolet light region (Stephanou & Manetas, 1995, 1997). However, when the exudate was experimentally removed, and the leaves were exposed to visible light supplemented with ultraviolet-B radiation, no effects on photosystem II photochemical eficiency or stomatal functions were detected and no epidermal browning was caused. This implies that the leaves of D. viscosa are resistant to ultraviolet-B radiation damage, and that the antitranspirant water conserving and allelopathic functions of the epicuticular material may predominate over its potential to protect against the ultraviolet-B radiation (Stephanou & Manetas, 1995, 1997).
The exudates have strong cytotoxic and genotoxic characteristics (Celik & Aslantürk 2010), resulting in allelopathic effects on seeds of other plants. For example, seed germination and radicle growth of co-occurring species have been inhibited (Stephanou & Manetas, 1997; Omezzine et al., 2011). The substances leach into the soil with heavy rains. Since the material is not toxic to the mother plant, D. viscosa has a competitive advantage over other species (Stephanou & Manetas, 1997). The allelopathic efect of D. viscosa might be an important mechanism for its success as an invasive species (Omezzine et al., 2011).
Phytomanagement against pollution and contaminated soils
D. viscosa has a high ability to grow on soils with high nickel, magnesium, or arsenic concentrations (Conesa et al., 2011, DPIPWE, 2011) and was proposed as bioindicator plant of these elements in the soil (Ater et al., 2000). Baker (1987) calls D. viscosa a pseudo-metalophyte because it can grow on soils which are contaminated by heavy metals. Moreover, it has a high potential as a bioaccumulator. On contaminated mine-tailing soils in Sardinia, the leaves of D. viscosa contained the highest concentrations of trace elements (Jimenez et al., 2011). It displayed very high metal concentrations in aboveground biomass (mean average of Zn: 1680 mg/kg, Pb: 420 mg/kg, Cd: 28 mg/kg), and was classified as good candidate for a phytoextraction procedure (Barbafieri et al., 2011). In Spain, D. viscosa growing in a mine showed a significant antimony bioaccumulation (Murciego et al., 2007). In a Mediterranean salt marsh polluted by mining wastes it had the highest metal and arsenic concentrations out of 21 plant species analysed (~270 mg/kg Pb, ~640mg/kg Zn, ~23 mg/kg As) (Conesa et al., 2011). The bioconcentration factor - the dry plant tissue/soil metal concentration quotient - calculated for D. viscosa was the highest out of the 21 plant species when the shoots were considered (Conesa et al., 2011).
Dittrichia viscosa showed a high transfer factor - i.e. the ratio of heavy metal concentrations in shoots to that in roots, a good index of translocation in a plant - for all the studied elements and plant species (Nogales & Benitez, 2006; Conesa et al., 2011). The contents of Zn, Fe and Al oxides, and the concentrations of trace metals found in D. viscosa were closely related to the total concentrations in the soil where this species grew in (Jimenez et al., 2011). This indicates that D. viscosa is suitable for phytostabilization, because it would allow its harvest without restrictions (Nogales & Benitez, 2006). Also Baker (1987) and Jimenez et al. (2011) state its potential to be used as phytostabilizer in polluted environments where it is essential to have a plant vegetation capable of developing in these environments in order to stabilize the spoil heaps and prevent the spreading of contaminants by leaching or wind dispersion (Hashimoto et al., 2008). Dittrichia viscosa may be useful in metal-polluted environments as it is one of the few species capable of growing in soils highly contaminated with Pb, Zn and Cu (Nogales & Benitez, 2006; Jimenez et al., 2011).
Dittrichia viscosa is even suggested for the combined use of biostabilized olive-derived wastes (Nogales & Benitez, 2006). Dittrichia viscosa could constitute an alternative and appropriate tool in phytostabilization processes in soils contaminated by heavy metals in the Mediterranean area where the plant is distributed widely and olive waste is produced in huge quantities.
Allelopathic potential
Flavonoids and other secondary compounds are often implicated in allelopathic reactions, usually inhibiting seed germination and root growth of other plant species (Rice, 1979; Levizou et al., 2002; Celik & Aslantürk, 2010; Omezzine et al., 2011). Under its influence, root length is reduced and the formation of root hairs is completely suppressed, the frequency of cell divisions in the meristematic zone is decreased, and cuticular transpiration is reduced (Stephanou & Manetas, 1995; Levizou et al., 2002). The extracts have cytotoxic effects on cell division in other plant species so that root growth is significantly inhibited (Celik & Aslantürk, 2010), but they are not toxic for the seeds of the mother plant (Levizou et al., 2002). In this way, D. viscosa has a competitive advantage over other germinating species. This is emphasized by the seasonal peak in the concentration of the active material, exactly in the period when most Mediterranean plants germinate, when the autumn rains set in and the allelopathic substances are washed into the soil (Stephanou & Manetas, 1997; Levizou et al., 2002). Leaf phenolic concentration in the leachates varies according to environmental conditions and is correlated with the extent of allelopathic interference (Stephanou & Manetas, 1997; Levizou et al., 2002).
Leaf epicuticular substances were also shown to be strong allelopathy agents for N2-fixing soil cyanobacteria, decreasing dramatically the photosynthetic assimilation of CO2 and increasing the heterocyst-to-vegetative cell ratio and most likely the assimilation of N2 of the cyanobacteria (Stamatakis& Konstantopoulou, 2001).
Flavonoids and other phenolics accumulate in periods when resource availability is low and growth is restricted (Gershenzon, 1994); then, more carbon can be diverted to the production of carbon-based secondary metabolites. Karageorgou et al. (2002) demonstrated that the leachable allelopathic material per plant was enhanced by nutrient addition and reduced by water stress.
Control of plant diseases, fungicidal and nematicidal effects
Leaf extracts of D. viscosa also have fungicidal, nematicidal, and antibacterial activity (Qasem et al., 1995; Maoz et al., 1999; Oka et al., 2001; Cafarchia et al., 2001; Mamoci et al., 2011). Its essential oils displayed anti-Helicobacter activity (Miguel et al., 2008). The natural products of these plants therefore can be employed in combating widespread infections. Plant extracts are also a source of bioactive ingredients with significant acaricidal activity that could lead to the development of new and safe bio-acaricides (Mansour et al., 2004).
Dittrichia viscosa may be used as a herbal source for fungicidal preparations against foliar diseases caused by pathogens belonging to the families Oomycetes, Ascomycetes, and Basidiomycetes (Wang et al., 2004). The material strongly inhibited spore germination and mycelial growth in a variety of hystopathogenic fungi (Stavrianakou et al., 2005). Downy mildew was controlled by extracts of D. viscosa, and the two major inhibitory compounds were identified as tomentosin and costic acid, which also displayed excellent activity against the disease in the field (Cohen et al., 2006). The phytotoxic effect is differential and tissue specific with concentrations in the following decreasing order: leaf > flower > stem > root (Omezzine et al., 2011). The fungistatic activity of the plant extract increases with plant age, and shoots show more inhibitory effects than roots, while high temperatures drastically reduce the antifungal efects of D. viscosa (Qasem et al., 1995).
The sesquiterpene tayunin isolated from leaves of D. viscosa inhibits the growth of the fungi Microsporum canis and Trichophyton rubrum (Maoz et al., 1999). Antifungal effects of shoot extracts or the incorporation of dried shoot material of D. viscosa in the growing media inhibited the growth of wheat leaf spot fungus (Helminthosporium sativum) and tomato wilt fungus (Fusarium oxysporum) (Qasem et al., 1995).
Disease control efficacy of leaf extracts was similar from material collected from May to October, suggesting that, for practical use, harvests can be conducted during most of the growing season (Wang et al., 2004). Also Cohen et al. (2006) found no seasonal fuctuations in the control efficacy of extracts from D. viscosa. Wang et al. (2004) found that "dried leaves of Inula viscosa stored at room temperature for a year produced an extract as effective as that from newly harvested dried leaves or as that from dry leaves kept on the bench for 4 years. These findings show that the active ingredients are stable, which is valuable information for industrial production of the extracts". On the other hand, Nikolakaki & Christodoulakis (2004) found that anti-fungal factors produced in tissue cultures were not found in the leaves, because yeasts attack the leaves, which deteriorate before producing calluses. Calluses formed prior to yeast infection develop a zone of inhibition against contaminating yeasts indicating the production and release of anti-fungal factors from undifferentiated cultured cells. This observation is of pharmaceutical importance and needs to be further investigated.
Applications as secondary plants in biological pest control
Biological pest control is a method to control pests without the application of potentially dangerous pesticides using tritrophic interactions as key function. Pest populations are reduced by natural enemies, also known as biological control agents, e.g. predators, parasitoids, and pathogens. The predator, parasite, or disease will attack the harmful pest. In order to increase and stabilize the presence of natural enemies, secondary plants may be employed which increase the occurrence of beneficial organisms such as insects by providing shelter and alternative prey. It is a common practice that plants are added to a crop system with the aim of increasing the efficiency of biological control (Ripper, 1944; Juniper & Southwood, 1986; van Lenteren, 1995; van Lenteren & Woets, 1998; Cortesero et al., 2000; Halaji et al., 2000; Gurr et al., 2004; Wäckers et al., 2005; Pontin et al., 2006; Vattala et al., 2006; Straub et al., 2008; Lundgren et al., 2009; Lambion, 2011; Parolin et al., 2014).
There are different ways to employ secondary plants (Parolin et al., 2012a, b); they can be either located near agricultural crops, in the spontaneous flora or planted on purpose in a mix of local species, on the border of crop felds or outside of greenhouses (Pasini et al., 1998; Gabarra et al., 2004; Lambion & Boisnard, 2009; Nannini et al., 2011). In greenhouses of Spain, the installation of M. caliginosus was more efficient when D. viscosa was distributed between the crops rather than grouping the plants at the border (FEDER, 1997).
Dittrichia viscosa may act as companion plant by enhancing chemical defence of the crop plants through its allelopathic characteristics (Parolin et al., 2012a). It may also act as insectary plant (Bas et al., 2003). This is a flowering plant which attracts and possibly maintains, with its nectar and pollen resources, a population of natural enemies. D. viscosa attracts pollinators because of its long period of flowering and strong smell. It may also act as banker plant (Alomar & Albajes, 2003; ITAB, 2005; Sacco et al., 2009; Ingegno et al., 2011), a rearing and release system of natural enemies (Frank, 2010; Huang et al., 2011; Parolin et al., 2012a, 2012b).
Dittrichia viscosa is useful in hosting the natural enemies of common pests in the Mediterranean region. It plays an important role in the conservation and/or increase of predator populations in agro-ecosystems. For example, it can be a useful reservoir of aphid parasitoids (Kavallieratos et al., 2002), phytoseiid mites (Tixier et al., 2000), predators like Eupelmus urozonus which control the gall-forming olive fies Myopites stylata (Franco-Micán et al., 2010; Warlop et al., 2010), or the mirid predator Macrolophus caliginosus, just to name a few.
Dittrichia viscosa is the main natural host for mirid bugs such as Macrolophus melanotoma or M. caliginosus, and contributes considerably to its maintenance in agro-ecosystems, its winter survival, and its colonization of neighbouring vegetable crops (FEDER, 1997; Pasini et al., 1998; Alomar & Albajes, 2003; Bas et al., 2003; Favas et al., 2003; Sanchez et al., 2003; Gabarra et al., 2004; Perdikis et al., 2007). This predator is useful against pests on pepper and tomato (Cano et al., 2009), cultivated Pelargonium plants (pest: Cacyreus marshalli; Sacco et al., 2009). It is very active and moves among plants, and is capable of dispersing and establishing on crops up to 20 m from the secondary plants (Alomar & Albajes, 2003). M. caliginosus can successfully develop and reproduce on D. viscosa, even when no pests are present, and the polyphagous mirid predator needs to feed only on the plant. D. viscosa is among the most promising host plant species due to its rusticity, its high ratio of predators:phytophages and its ability to shelter M. caliginosus and D. errans (Lambion & Boisnard, 2009; Lambion, 2011). D. viscosa alone sheltered many more predatory bugs than the spontaneous flora did (Lambion, 2011) and the maintenance of non-cultivated host plants of D. viscosa with Macrolophus species in field edges could contribute in the colonization of nearby crop felds (Perdikis et al., 2007). Alomar et al. (2002) showed that great numbers of M. caliginosus entered tomato fields surrounded by unaltered and complex vegetation, in which D. viscosa occurred.
However, a recent study showed that the presence of D. viscosa can also result in an overinfestation with pests (Parolin et al., 2013b). Although D. viscosa was efcient as a biocontrol plant (sensu Parolin et al., 2014) for several species combinations, its presence caused an extreme increase of the population of the whitefy pest Trialeurodes vaporariorum in a greenhouse, and in the same experiment did not act as banker plant for Macrolophus pygmaeus (Parolin et al., 2013b).
A special regard must be given to the traditional land use form and its management of D. viscosa, which is one of the plant species historically associated with olive groves, important cultivations in the Mediterranean (ITAB, 2005; Franco-Micán et al., 2010). Its flowers are infested by Myopites stylata, a dipteran which causes gall formation and whose larvae are parasitized by the generalist parasitoid Eupelmus urozonus associated with D. viscosa, and which overwinters in the larvae of M. stylata on D. viscosa. Since E. urozonus is also one of the main parasitoids associated with the major key pest of the olive crop, the olive fruit-fly (Bactrocera oleae), this parasitic complex is very valuable to control the olive fruit-fly (Broumas et al., 2002; Petacchi et al., 2003; ITAB, 2005; Boccaccio & Petacchi, 2009; Franco-Micán et al., 2010; Warlop et al., 2010). Therefore, in the presence of D. viscosa, the olive fruit-fly in winter is predated by E. urozonus, which in turn in summer predates efficiently the olive fruit-fly (ITAB, 2005).
The galls formed on D. viscosa can harbor 1-4 larvae of parasitoids, with gall size being an important factor affecting the number of emerging adults (Franco-Micán et al., 2010). Individuals belonging to different insect families emerged from a same gall (Franco-Micán et al., 2007). The number of families and adults present in galls changed depending on the locality where galls were collected (Franco-Micán et al., 2010).
Traditionally, olive plantations were associated to highly diversify the natural herbaceous vegetation, which directly influenced the presence of beneficial insects (ITAB, 2005). They were then believed to be competing for water and nutrients with the major agronomic crops, and to offer a home for plant-eating insects which would damage the crops (ITAB, 2005). For decades, the spontaneous herbs were therefore eradicated, resulting in poor vegetation and arthropod fauna present and eliminating all kind of self-regulation of the system. The costs to control pests, diseases, nutritional degradation, and erosion without relying on the self-regulated system are very high (Dominguez-Gento et al., 2002). Olive culture is subjected to soil erosion with 80 millions of tons of soil being lost every year only in Andalusia, leading to impoverishment and desertification, leaching and groundwater pollution, and finally to the disappearance of the native fauna and flora (Vallebrera & Perdrix, 2003). The value of intact landscape structures associated to agrobiodiversity (Altieri & Nicholls, 2001) and the inclusion of high diversity of autochthonous and functional types species are important components for biological control (Tscharntke et al., 2007).
Dispersal from refuges and inter-crop movement are keyfactors (Alomar & Albajes, 2003). Even in highly diversifed agro-ecosystems, there is no guarantee that the useful predators move from the local plants (e.g., D. viscosa) to adjacent crop fields and into greenhouses (FEDER, 1997; Pasini et al., 1998; Tixier et al., 2000; Alomar & Albajes, 2003; Lambion & Boisnard, 2009; Lambion, 2011). Gabarra et al. (2004) showed that D. viscosa facilitated considerably the entrance of mirid bugs to greenhouses. In general, the composition of the vegetation, and the topography surrounding greenhouses may favor the colonization of the crops by M. caliginosus, but also the early immigration of whitefy adults (Gabarra et al., 2004).
Another issue of using wild plants for biological control is that they may be vectors for possibly dangerous organisms, such as viruses. A risk assessment of including D. viscosa as possible host of pests and viruses that could damage the crops was performed by Cano et al. (2009), and showed that no viruses were detected. Furthermore, neither by mechanical inoculation nor by whitefy transmission could D. viscosa be infected with viruses (Cano et al., 2009). The same was found by Alomar as described in FEDER (1997), and by Perdikis et al. (2011).
CONCLUSIONS
The present review shows that D. viscosa offers a large variety of ecosystem functions for its employment in the Mediterranean basin. D. viscosa is an important species that can be used for phytorremediation, bioaccumulator or bioindicator in the Mediterranean ecosystem. We hope that this review contributes to the understanding of the interesting and widespread D. viscosa which grows in the Mediterranean ecosystem.
REFERENCES
1. Abu Zarga, M.H., E.M. Hamed, S.S. Sabri, W. Voelter & K.-P. Zeller (1998). New sesquiterpenoids from the Jordanian medicinal plant Inula viscosa. Journal of Natural Products 61: 798-800. [ Links ]
2. Al-Dissi, N.M., A.S. Salhab & H.A. Al-Hajj (2001). Effects of Inula viscosa leaf extracts on abortion and implantation in rats. Journal of Ethnopharmacology 77: 117-121. [ Links ]
3. Alomar, O. & R. Albajes (2003). Habitat management for conservation of the native predator Macrolophus caliginosus. Landscape Management for Functional Biodiversity IOBC wprs Bulletin 26: 7-12. [ Links ]
4. Alomar, O., M. Goula & R. Albajes (2002). Colonization of tomato fields by predatory mirid bugs (Hemiptera: Heteroptera) in northern Spain. Agriculture, Ecosystems and Environment 89: 105-115. [ Links ]
5. Altieri, M.A. & C.I. Nicholls (2001). Biodiversity and pest management in agroecosystems. Food Products Press, New York. [ Links ]
6. Amundson, R.G., R.J. Kohut & J.A. Laurence (1995). Influence of foliar N on foliar soluble sugars and starch of red spruce saplings exposed to ambient and elevated ozone. Tree Physiology 15: 167-174. [ Links ]
7. Anderberg, A.A. (1991). Taxonomy and phylogeny of the tribe Inuleae (Aasteraceae). Plant Systematics and Evolution 176: 75-123. [ Links ]
8. Anderberg, A.A., P. Eldenäs, R.J. Bayer & M. Englund (2005). Evolutionary relationships in the Asteraceae tribe Inuleae (incl. Plucheeae) evidenced by DNA sequences of ndhF; with notes on the systematic position of some aberrant genera. Organisms Diversity and Evolution 5: 135-146. [ Links ]
9. Ater, M., C. Lefèbvre, W. Gruber & P. Meerts (2000). A phytogeochemical survey of the fora of ultramafic and adjacent normal soils in North Morocco. Plant and Soil 218: 127-135. [ Links ]
10. Baker, A.J.M. (1987). Metal tolerance. New Phytologist 106: 93-111. [ Links ]
11. Barbafieri, M., C. Dadea, E. Tassi, F. Bretzel & L. Fanfani (2011). Uptake of heavy metals by native species growing in a mining area in Sardinia, Italy: discovering native flora for phytoremediation. International Journal of Phytoremediation 1: 985-97. [ Links ]
12. Barrero, A.F., M.M. Herrador, P. Arteaga & J.V. Catalan (2008). Dittrichia viscosa L. Greuter: Phytochemistry and biological activity. Natural Product Communications 3: 1799-1804. [ Links ]
13. Bas, C., J. Arnó & O. Alomar (2003). Effects of prey, Bemisia tabaci, and host plant switching on life-history parameters of the predator Macrolophus caliginosus. Session IV Biological Control. Abstract 3rd International Bemisia Workshop, Barcelona March 2003. 90 p. [ Links ]
14. Boccaccio, L. & R. Petacchi (2009). Landscape effects on the complex of Bactrocera oleae parasitoids and implications for conservation biological control. Biocontrol 54: 607-616. [ Links ]
15. Broumas, T., G. Haniotakis, C. Liaropoulos, T. Tomazou & N. Ragoussis (2002). The efficacy of an improved form of the masstrapping method, for the control of the olive fruit fy, Bactrocera oleae (Gmelin), Dipt., Tephritidae. pilotscale feasibility studies. Journal of Applied Entomology 126: 217-223. [ Links ]
16. Brullo, S. & G. De Marco (2000). Taxonomical revision of the genus Dittrichia (Asteraceae). Portugaliae Acta Biologica 19: 341-354. [ Links ]
17. Cafarchia, C., N. De Laurentis, M.A. Milillo, V. Losacco & V. Puccini (2001). Fungistatic activity of a sesquiterpene lactone (tomentosin) isolated from fresh Inula viscosa (Asteraceae) flowers from the Pulgia region. Parasitologia 43: 117-121. [ Links ]
18. Cano, M., E. Vila, D. Janssen, G. Bretones, E. Salvador, L. Lara & M.M. Tellez (2009). Selection of refuges for Nesidiocoris tenuis (Het.: Miridae) and Orius laevigatus (Het.: Anthocoridae. virus reservoir risk assessment. IOBC/WPRS Bulletin 49: 281-286. [ Links ]
19. Celik, T.A. & O.S. Aslantürk (2010). Evaluation of cytotoxicity and genotoxicity of Inula viscosa leaf extracts with Allium test. Journal of Biomedicine and Biotechnology 2010: 1-8. [ Links ]
20. Chamaco, A., A. Fernandez, C. Fernandez & J. Altarejos (2000). Composition of essential oil of Dittrichia viscosa. Eppos 29: 3-8. [ Links ]
21. CIVAM (2011). Les Conseils de Saison du CIVAM Oléicole 13 : 1-4. [ Links ]
22. Cohen, Y., W. Wang, B.-H. Ben-Daniel & Y. Ben-Daniel (2006). Extracts of Inula viscosa control downy mildew of grapes caused by Plasmopara viticola. Phytopathology 96: 417-424. [ Links ]
23. Conesa, H.M., A. Maria-Cervantes, J. Alvarez-Rogel & M.N. Gonzalez-Alcaraz (2011). Influence of soil properties on trace element availability and plant accumulation in a Mediterranean salt marsh polluted by mining wastes: Implications for phytomanagement RID A-3481-2009. Science of the Total Environment 409: 4470-4479. [ Links ]
24. Cortesero, A.M., J.O. Stapel & W.J. Lewis (2000). Understanding and manipulating plant attributes to enhance biological control. Biological Control 17: 35-49. [ Links ]
25. DAISIE (2012). Delivering Alien Invasive Species Inventories for Europe, http://www.europe-aliens.org/speciesFactsheet.do?speciesId=22359#. Last access June 2013. [ Links ]
26. De Laurentis, N., V. Losacco, M.A. Milillo & O. Lai (2002). Chemical investigations of volatile constituents of Inula viscosa (L.) Aiton (Asteraceae) from different areas of Apulia, Southern Italy. Delpinoa 44: 115-119. [ Links ]
27. Di Castri, F. (1973). Climatographical comparisons between Chile and the western coast of North America. In: Di Castri F. & Mooney H.A. (eds), pp. 21-36. Mediterranean-type ecosystems. Springer Verlag, Berlin, Germany. [ Links ]
28. Dias, T., S. Malveiro, M.A. Martins-Loucao, L.J. Sheppard & C. Cruz (2011). Linking N-driven biodiversity changes with soil N availability in a Mediterranean ecosystem. Plant and Soil 341: 125-136. [ Links ]
29. Dominguez-Gento, A., J. Roselló & J. Aguado (2002). Diseño y manejo de la diversidad vegetal en agricultura ecológica. Phytoma-España y Sociedad Española de Agricultura Ecológica, València.132 p. [ Links ]
30. Doussi, M.A. & C.A. Tanos (1997). Ecophysiology of seed germination in composites inhabiting fire-prone Mediterranean ecosystems. In: Ellis R.H., Black M., Murdoch A.J., Hong T.D. (eds) pp. 641-649. Basic and applied aspects of seed biology. Kluwer Academic Publishers: Dordrecht, Holland. [ Links ]
31. DPIPWE (Department of Primary Industries, Parks, Water and Environment) (2011). False Yellow Head Weed Management Plan. Statutory Weed Management Plan as at June 2011. Tasmania / Australia. http://www.dpiw.tas.gov.au/inter.nsf/Attachments/LBUN-8EZ5HA?open, last access June 2011. [ Links ]
32. Favas C., D. Perdikis & D. Lykouressis (2003). Biological characteristics of Macrolophus caliginosus (Hemiptera: Miridae) when feeding on the non-cultivated plant Dittrichia viscosa (Asteraceae). IOBC/wprs Bulletin 26: 1-139. [ Links ]
33. FEDER (1997). Control biológico por conservación: papel de las hierbas espontáneas y los cultivos como refugios y fuentes de depredadores. Alomar O. (ed), Unpubl. Report, 13pp. INIASC00 [ Links ]
34. Field, C. & H.A. Mooney (1983). Leaf age and seasonal efects on light, water, and nitrogen use in a California shrub. Oecologia 36: 348-355. [ Links ]
35. Fimiani, P. & M.C. Digilio (1993). Bionomics of Myopites stilata F. (Diptera, Tephritidae) and its natural enemies in Vivara Island (Gulf of Naples). Bollettino della Societá Naturalistica di Napoli 101:53-63. [ Links ]
36. Franco-Micán, S., J. Castro & M. Campos (2010). Preliminary study of the parasitic complex associated with Dittrichia viscosa in Andalusia (Spain). IOBC/WPRS Bulletin 53: 139-143. [ Links ]
37. Frank, S.D. (2010). Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biological Control 52: 8-16. [ Links ]
38. Gabarra, R., O. Alomar, C. Castañé, M. Goula & R. Albajes (2004). Movement of greenhouse whitefy and its predators between inand outside of Mediterranean greenhouses. Agriculture Ecosystems Environment 102: 341-348. [ Links ]
39. Gershenzon, J. (1994). Metabolic costs of terpenoid accumulation in higher-plants. Journal of Chemical Ecology 20: 1281-1328. [ Links ]
40. Grande, M., F. Piera, A. Cuenca, P. Torres & S. Bellido (1985). Flavonoids from Inula viscosa. Planta Medica 39: 414-419. [ Links ]
41. Grande, M., P. Torres, F. Piera & I.S. Bellido (1992). Triterpenoids from Dittrichia viscosa. Phytochemistry 31: 1826-1828. [ Links ]
42. Groves, R.H. (1991). The biogeography of Mediterranean plant invasions. In: Groves, R.H. & F. di Castri (eds.), pp. 427-438. Biogeography of Mediterranean invasions. Cambridge University Press, City. [ Links ]
43. Gurr, G.M., S.D. Wratten, J. Tylianakis, J. Kean & M. Keller (2004). Providing plant foods for insect natural enemies in farming systems: balancing practicalities and theory. In: Wackers, F.L., P.C.J. van Rijn & J. Bruin (eds), pp. 326-347. Plant-derived food and plant-carnivore mutualism. Cambridge University Press, Cambridge. [ Links ]
44. Halaji, J., A.B. Cady & G.W. Uetz (2000). Modular habitat refuge enhance generalist predators and lower plant damage in soybeans. Environmental Entomology 29: 383-393. [ Links ]
45. Hashimoto, Y., H. Matsufuru & T. Sato (2008). Attenuation of lead leachability in shooting range soils using poultry waste amendments in combination with indigenous plant species. Chemosphere 73: 643-649. [ Links ]
46. Hernández, V., del Carmen Recio, M., Máñez, S., Prieto, J.M., Giner, R.M. & Ríos, J.L. (2001). A mechanistic approach to the in vivo anti-inflammatory activity of sesquiterpenoid compounds isolated from Inula viscosa. Planta Medica 67: 726-731. [ Links ]
47. Huang, N., A. Enkegaard, L.S. Osborne, P.M.J. Ramakers, G.J. Messelink, J. Pijnakker & G. Murphy (2011). The banker plant method in biological control. Critical Reviews in Plant Sciences 30: 259-278. [ Links ]
48. Ingegno, B.L., M.G. Pansa & L. Tavella (2011). Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus (Heteroptera: Miridae). Biological Control 58: 174-181. [ Links ]
49. ITAB (2005). Produire des fruits en agriculture biologique. Alter Agri 72 :1-32. [ Links ]
50. Jimenez, M.N., G. Bacchetta, M. Casti, F.B. Navarro, A.M. Lallena & E. Fernandez-Ondono (2011). Potential use in phytoremediation of three plant species growing on contaminated mine-tailing soils in Sardinia. Ecological Engineering 37: 392-398. [ Links ]
51. Juniper, B.E. & T.R.E. Southwood (1986). Insects and the Plant Surface, Edward Arnold, London. Pages. [ Links ]
52. Karageorgou, P., E. Levizou & Y. Manetas (2002). The influence of drought, shade and availability of mineral nutrients on exudate phenolics of Dittrichia viscosa. Flora 197: 285-289. [ Links ]
53. Kavallieratos, N.G., G.J. Stathas, C.G. Athanassiou & G.T. Papadoulis (2002). Dittrichia viscosa and Rubus ulmifolius as reservoirs of aphid parasitoids (Hymenoptera : Braconidae : Aphidiinae) and the role of certain coccinellid species. Phytoparasitica 30: 231-242. [ Links ]
54. Lambion, J. (2011). Functional biodiversity in Southern France: a method to enhance predatory mirid bug populations. Acta Horticulturae 915. [ Links ]
55. Larcher, W. & W. Thomaser-Thin (1988). Seasonal changes in energy content and storage patterns of Mediterranean sclerophylls in a northernmost habitat. Acta Oecologica 9: 271-283. [ Links ]
56. Lauro, L. & C. Rolih, C. (1990). Observations and research on an extract of Inula viscosa Ait. Bollettino della Società Italiana di Biologia Sperimentale 66: 829-834. [ Links ]
57. Levizou, E., P. Karageorgou, G.K. Psaras & Y. Manetas (2002). Inhibitory effects of water soluble leaf leachates from Dittrichia viscosa on lettuce root growth, statocyte development and graviperception. Flora 197: 152-157. [ Links ]
58. Lundgren, J.G., K.A.G. Wyckhuys & N. Desneux (2009). Population responses by Orius insidiosus to vegetational diversity. Biocontrol 54: 135-142. [ Links ]
59. Mamoci, E., I. Cavoski, V. Simeone, D. Mondelli, L. Al-Bitar & P. Caboni (2011). Chemical composition and in vitro activity of plant extracts from Ferula communis and Dittrichia viscosa against postharvest fungi. Molecules 16: 2609-2625. [ Links ]
60. Máñez, S., M.C. Recio, I. Gil, C. Gómez R.M. Giner, P.G. Waterman & J.L. Ríos (1999). A glycosyl analogue of diacylglycerol and other antiinflammatory constituents from Inula viscosa. Journal of Natural Products 62: 601-604. [ Links ]
61. Mansour, F., H. Azalzeh, B. Saad, Y. Tadmor, F. Abo-Moch & O. Said (2004). The potential of middle eastern flora as a source of new safe bio-acaricides to control Tetranychus cinnabarinus, the carmine spider mite. Phytoparasitica 32: 66-72. [ Links ]
62. Maoz, M., Y. Kashman & I. Neeman (1999). Isolation and identification of a new antifungal sesquiterpene lactone from Inula viscosa. Planta Medica 65: 281-282. [ Links ]
63. Miguel, G., L. Faleiro, C. Cavaleiro, L. Salgueiro & J. Casanova (2008). Susceptibility of Helicobacter pylori to essential oil of Dittrichia viscosa subsp revoluta RID E-6147-2010 RID F-4813-2011. Phytotherapy Research 22: 259-263. [ Links ]
64. Mooney, H.A. (1987). The impact of environmental stress on plant performance in Mediterranean climate ecosystems. In: Tenhumen,J.D., F. M. Catarino, O.L. Lange, W.G. Oechel (eds.), pp. 661-668. Plant response to stress: functional analysis in Mediterranean ecosystems. Springer Verlag, Berlin, Germany. [ Links ]
65. Murciego, A.M., A.G. Sánchez, M.A.R. González, E.P. Gil, C.T. Gordillo, J.C. Fernández & T.B. Triguero (2007). Antimony distribution and mobility in topsoils and plants (Cytisus striatus, Cistus ladanifer and Dittrichia viscosa) from polluted Sb-mining areas in Extremadura (Spain). Environmental Pollution 145: 15-21. [ Links ]
66. Nannini, M., F. Corda, F. Foddi, L. Manca, C. Anca, G. Murgia, R. Pisci & F. Sanna (2011). A survey of natural enemies of whitefly (Homoptera: Aleyrodidae) on selected weeds in a Mediterranean tomato production area. In: Vandekerkhove, B. (ed.). pp. 199-209. Proceedings 63rd International Symposium on Crop Protection. Gent, May 24, 2011. [ Links ]
67. Nikolakaki, A. & N.S. Christodoulakis (2004). Leaf structure and cytochemical investigation of secretory tissues in Inula viscosa. Botanical Journal of the Linnean Society 144: 437-448. [ Links ]
68. Nogales, R. & E. Benitez (2006). Absorption of zinc and lead by Dittrichia viscosa grown in a contaminated soil amended with olive-derived wastes. Bulletin of Environmental Contamination and Toxicology 76: 538-544. [ Links ]
69. Oka, Y., B.H. Ben-Daniel & Y. Cohen (2001). Nematicidal activity of powder and extracts of Inula viscosa. Nematology 3: 735-742. [ Links ]
70. Omezzine, F., A. Rinez, A. Ladhari, M. Farooq & R. Haouala (2011). Allelopathic potential of Inula viscosa against crops and weeds. International Journal of Agriculture and Biology 13: 841-849. [ Links ]
71. Parolin P., C. Bresch, R. Brun, A. Bout, R. Boll, N. Desneux & C. Poncet (2012a). Secondary plants used in biological control: a review. International Journal of Pest Management 58: 91-100. [ Links ]
72. Parolin, P., C. Bresch, C. Poncet & N. Desneux (2012b). Functional characteristics of secondary plants for increased pest management. International Journal of Pest Management 58: 369-377. [ Links ]
73. Parolin P., Ion Scotta M . & Bresch C. (2013a): Notes on the phenology of Dittrichia viscosa. Journal of Mediterranean Ecology 12: 27-35. [ Links ]
74. Parolin P., Bresch C., Ottenwalder L., Ion-Scotta M., Brun R., Fatnassi H. & Poncet C. (2013b): False yellowhead (Dittrichia viscosa) causes over infestation with the whitefy pest (Trialeurodes vaporariorum) in tomato crops. International Journal of Agricultural Policy and Research 1(10): 311-318. [ Links ]
75. Parolin P., Bresch C., Poncet C., Desneux N. (2014): Introducing the term 'Biocontrol Plants' for Integrated Pest Management. Scientia Agricola 71: 77-80. [ Links ]
76. Pasini, C., F. D'Aquila, M. Gandolfo, M. Costanzi & L. Mirto (1998). Macrolophus caliginosus in biological control. Colture Protette 27: 43-46. [ Links ]
77. Pelser, P.B., A.H. Kennedy, E.J. Tepe, J.B. Shidler, B. Nordenstam, J.W. Kadereit & L.E. Watson (2010). Patterns and causes of incongruence between plastid and nuclear Senecioneae (Asteraceae) phylogenies. American Journal of Botany 97: 856-873. [ Links ]
78. Perdikis, D., Ch. Favas, D. Lykouressis & A. Fantinou (2007). Ecological relationships between non-cultivated plants and insect predators in agroecosystems: the case of Dittrichia viscosa (Asteraceae) and Macrolophus caliginosus (Hemiptera: Miridae). Acta Oecologica 31: 299-306. [ Links ]
79. Perdikis, D., A. Fantinou & D. Lykouressis (2011). Enhancing pest control in annual crops by conservation of predatory Heteroptera. Biological Control 59: 13-21. [ Links ]
80. Perdikis, D., Kapaxidi, E. & Papadoulis, G. (2008). Biological control of insect and mites pests in greenhouse Solanaceous crops. The European Journal of Plant Science and Biotechnology 2: 125-144. [ Links ]
81. Perez-Alonso, M.J., Velasco-Negueruela, A., Emin Duru, M., Harmandar, M. & García Vallejo, M.C. (1996). Composition of the volatile oil from the aerial parts of Inula viscosa. Flavour and Fragrance Journal 11: 349-351. [ Links ]
82. Perez-Fernandez, M. A., E. Calvo-Magro & D. Ferrrer-Castan (2006). Simulation of germination of pioneer species along an experimental drought gradient. Journal of Environmental Biology 27: 679-685. [ Links ]
83. Petacchi, R., I. Rizzi & D. Guidotti (2003). The "lure and kill" technique in Bactrocera oleae (Gmel.) control: effectiveness indices and suitability of the technique in areawide experimental trials. International Journal of Pest Management 49: 305-311. [ Links ]
84. Pontin, D.R., M.R. Wade, P. Kehrli & S.D. Wratten (2006). Attractiveness of single and multiple species flower patches to beneficial insects in agroecosystems. Annals of Applied Biology 148: 39-47. [ Links ]
85. Qasem, J.R., A.S. Al-Abed & H.A. Abu-Blan (1995). Antifungal activity of clammy inula (Inula viscosa) on Helminthosporium sativum and Fusarium oxysporum f. sp. lycopersici. Phytopathologia Mediterranea 34: 7-14. [ Links ]
86. Rice, E.L. (1979). Allelopathy - an update. Botanical Review 45: 15-109. [ Links ]
87. Ripper, W. (1944). Biological control as a supplement to chemical control of insect pests. Nature 153: 448-452. [ Links ]
88. Roldan-Fajardo, B.E. (1994). Effect of Indigenous arbuscular mycorrhizal endophytes on the development of 6 wild plants colonizing a semiarid area in south-east Spain. New Phytologist 127: 115-121. [ Links ]
89. Sacco, M., E. Arato, F. Pasin, G. Pellizzari & C. Pasini (2009). Remarks on the activity of Macrolophus caliginosus Wagner as predator of Cacyreus marshalli Butler, exotic phytophagous of cultivated Pelargonium. Protezione delle Colture 3: 21-24.
90. Sanchez, J. A., D.R. Gillespie & R.R. McGregor (2003). The effects of mullein plants (Verbascum thapsus) on the population dynamics of Dicyphus hesperus (Heteroptera : Miridae) in tomato greenhouses. Biological Control 28: 313-319. [ Links ]
91. Schinella, G.R., H.A. Tournier, J.M. Prieto, P. Mordujovich de Buschiazzo & J.L. Rios (2002). Antioxidant activity of anti-inflammatory plant extracts. Life Sciences 70: 1023-1033. [ Links ]
92. Scott, J.K., K.L. Batchelor, N. Ota & P.B. Yeoh (2008). Modelling climate change impacts on sleeper and alert weeds. Appendix B: Results of Climex Models Part 2. http://www.csiro.au/en/Outcomes/Climate/Adapting/Sleeper-Alert-Weeds-Appendix-B.aspx. [ Links ]
93. Shtacher, G. & Y. Kashman (1971). Chemical investigation of volatile constituents of Inula viscosa Ait. Tetrahedron 27: 1343-1349. [ Links ]
94. Silva, D., E. Denham, L. Faleiro, G. Miguel, C. Cavaleiro & L. Salgueiro (2005). Antimicrobial activity of the essential oils of Dittrichia viscosa subsp. viscosa on Helicobacter pylori. VI World Congress on Medicinal and Aromatic Plants for Human Welfare. WOCMAPII. Acta Horticulturae 680: 147-151. [ Links ]
95. Sinden J., R. Jones, S. Hester, D. Odom, C. Kalisch & R. James (2004). The economic impact of weeds in Australia. Report to the CRC for Australian Weed Management. Pp. 1-65. [ Links ]
96. Stamatakis K. & M. Konstantopoulou (2001). Allelopathy effects of aqueous rinses of Dittrichia viscosa (L.) on the photosynthesis and cell proliferation of N2-fxing soil cyanobacteria. Photosynthesis Research 69: 119-122. [ Links ]
97. Stavrianakou, S., G. Liakopoulos & G. Karabourniotis (2006). Boron defficiency effects on growth, photsynthesis and relative concentrations of phenolics of Dittrichia viscosa (Asteraceae). Environmental and Experimental Botany 56: 293-300. [ Links ]
98. Stavrianakou, S., V. Liakoura, E. Levizou, P. Karageorgou, C. Delis, G. Liakopoulos, G. Karabourniotis, G. Manetas & Y. Manetas (2005). Allelopathic effects of water-soluble leaf epicuticular material from Dittrichia viscosa on seed germination of crops and weeds. Allelopathy Journal 14: 35-41. [ Links ]
99. Stephanou, M. & Y. Manetas (1995). Allelopathic and water conserving functions of leaf epicuticular exudates in the Mediterranean shrub Dittrichia viscosa. Australian Journal of Plant Physiology 22: 755-759. [ Links ]
100. Stephanou, M. & Y. Manetas (1997). Seasonal variations in UV-B absorbing capacity and allelopathic potential of Dittrichia viscosa leaf rinsates. Canadian Journal of Botany 75: 1371-1374. [ Links ]
101. Straub, C.S., D.L. Finke & W.E. Snyder (2008). Are the conservation of natural enemy biodiversity and biological control compatible goals. Biological Control 45: 225-237 [ Links ]
102. Tenhunen, J.D., H.P. Meister, M.M. Caldwell & O.L. Lange (1984). Environmental constraints on productivity of the Mediterranean shrub Quercus coccifera. Options Mediterranean 1: 33-53. [ Links ]
103. Tixier, M.S., S. Kreiter, P. Auger, G. Sentenac, G. Salva & M. Weber (2000). Phytoseid mite species located in uncultivated areas surrounding vineyards in three French regions. Acarologia 41: 127-140. [ Links ]
104. Tscharntke, T., R. Bommarco, Y. Clough, T.O. Crist, D. Kleijn, T.A. Rand, J.M. Tylianakis, S. van Nouhuys & S. Vidal (2007). Conservation biological control and enemy diversity on a landscape scale. Biological Control 43 : 294-309. [ Links ]
105. Vallebrera, A. & V. Perdrix (2003). Contrôle de la mouche méditeranéenne des fruits (Diptera: Tephitidae) à travers des strategies de périmètre. Journées Techniques Nationales Fruits et Legumes Biologiques. Perpignan. [ Links ]
106. van Lenteren, J.C. & J. Woets (1998). Biological and Integrated Pest Control in greenhouses. Annual Review of Entomology 33: 239-269. [ Links ]
107. van Lenteren, J.C. (1995). Integrated pest management in protected crops. In: Dent, D. (ed.), pp. 311-343. Integrated Pest. Management, Chapman and Hall, London. Pages. [ Links ]
108. Vattala, H.D., S.D. Wratten, C.B. Phillips & F.L. Wäckers (2006). The influence of flower morphology and nectar quality on the longevity of a parasitoid biological control agent. Biological Control 39: 179-185. [ Links ]
109. Wäckers, F.L., P.C.J. van Rijn & J. Bruin (2005). Plant-provided food for carnivorous insects: a protective mutualism and its applications. Cambridge University Press. City. [ Links ]
110. Wang, W., B.H. Ben-Daniel & Y. Cohen (2004). Control of plant diseases by extracts of Inula viscosa. Phytopathology 94: 1042-1047. [ Links ]
111. Warlop, F., E. Spagnol, C. Conand, J. Lecomte, R. Gimilio & G. Delvare (2010). Pourquoi et comment propager l'Inule visqueuse dans les oliveraies? P.1-4. Plaquette GRAB. [ Links ]
112. Werker, E. & A. Fahn (1981). Secretory hairs of Inula viscosa - Development, Ultrastructure, and Secretion. Botanical Gazette 142: 461-476. [ Links ]
113. Wollenweber, E., K. Mayer & J.N. Roitman (1991). Exudate flavonoids of Inula viscosa. Phytochemistry 30: 2445-2446. [ Links ]

