










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkPhyton (Buenos Aires)
versión On-line ISSN 1851-5657
Phyton (B. Aires) vol.83 no.2 Vicente López dic. 2014
ARTÍCULOS ORIGINALES
Water stress response of Tricepiro and its progenitors Triticale and Trigopiro
Respuesta al estrés hídrico del tricepiro y sus progenitores, triticale y trigopiro
Ruiz MA1,3, A Golberg1,2, ML Molas1
1 INTA Estación Experimental Anguil - Ing. Agr. Guillermo Covas. Ruta 5, km 580. CC 11. (6326) Anguil, La Pampa.
2 Facultad de Agronomía, Universidad Nacional de La Pampa. Ruta 35, km 334. CC 300. CP 6300. Santa Rosa, La Pampa.
3 Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa. Uruguay 151. CC 300. (6300) Santa Rosa, La Pampa.
Address Correspondence to: María de los Ángeles Ruiz. Tel.: (+54) 2954-495057. Fax: (+54) 2954-495057, e-mail: ruiz.maria@inta.gob.ar
Recibido / Received 5.VI.2013.
Aceptado / Accepted 12.VI.2013.
Abstract. In semiarid regions rainfall is often scarce and erratic in space and time, making valuable the exploration of news genotypes with superior behavior in dry land systems. Tricepiro (X Triticosecale Wittmack x X Agrotriticum Ciferri & Giacom) is a synthetic cereal with long cycle and high biomass production in semiarid environments. However its performance under controlled water stress has not yet been evaluated. The aim of this work was to investigate the effect of water stress on tricepiro genotypes in comparison with their progenitors: triticale and trigopiro. Morphophysiological traits related to forage yield were evaluated in the greenhouse and the field. In the greenhouse, water potential, stomatal resistance, root biomass and wilted/green leaves were significantly associated with forage production under severe water stress [leaf water potential (WP) = -3 MPa]. In the field, under mild-moderate stress (leaf WP = -1.23 MPa), forage yield was highly related to plant height and leaf area index. We found that under water stress tricepiro performed similar to triticale, and both were superior to trigopiro. Among tricepiros, L62 and the line RC50 were the best performing genotypes. Hence, tricepiro has potential to diversify cropping systems in Semiarid Regions.
Keywords: Tricepiro; Synthetic cereal; Drought; Semiarid environment.
Resumen. En las regiones semiáridas las precipitaciones son a menudo escasas y erráticas tanto desde el punto de vista espacial como temporal. Estas condiciones hacen necesaria la búsqueda de nuevos genotipos adaptados a la sequía. El Tricepiro (X Triticosecale Wittmack x X Agrotriticum Ciferri & Giacom) es un cereal sintético de ciclo largo y gran producción de biomasa en ambientes semiáridos; sin embargo, su respuesta a condiciones de estrés hídrico todavía no se ha investigado. El objetivo de este trabajo fue investigar el efecto del estrés hídrico sobre genotipos de tricepiro en comparación con sus progenitores: triticale y trigopiro. Se evaluaron algunas características morfofisiológicas de los genotipos relacionadas con la producción de forraje, en invernáculo y a campo. En invernáculo, el potencial hídrico, la resistencia estomática, la biomasa de raíces y la relación hojas secas/hojas verdes estuvieron significativamente asociadas con la producción de forraje bajo estrés hídrico severo (potencial hídrico foliar = -3 MPa). A campo, bajo un estrés hídrico moderado (-1,23 MPa), la producción de forraje estuvo estrechamente relacionada con la altura de la planta y con el índice de área foliar. Las experiencias mostraron que bajo estrés hídrico el tricepiro se comportó de manera similar al triticale, y ambos genotipos fueron superiores al trigopiro. Entre los tricepiros, L62 y la línea RC50 tuvieron un comportamiento superior. Esto sugiere que existe la posibilidad de utilizar a tricepiro para diversificar los sistemas de cultivo en regiones semiáridas.
Palabras clave: Tricepiro; Cereal sintético; Sequía; Ambiente semiárido.
INTRODUCTION
In semiarid regions, water is the most limiting factor for crop production. For this reason, successful agricultural and grazing systems in these areas must make an efcient use of precipitation. Forage systems (e.g., corn, millet and triticale) have higher water use efficiency than high frequency rotations of small grain and/or oilseed crops (Nielsen et al., 2006). Grazing livestock systems in most semiarid regions around the world are largely based on cultivated and native pastures (Entz et al., 2002; Clapham & Fedders, 2008). In cultivated pastures, perennial forages provide some of the livestock feed. In addition, small grains cereals are used regularly as forages in many areas to extend the grazing season during winter, fall and spring, and to complement established mixed pastures (McColoy et al., 1971; Brown & Almodares, 1976; Juskiw et al., 1999). Although there are associated costs with planting annual forages, spring-planted winter annuals establish readily, and the yield distribution of spring-planted winter annuals provides forage for grazing in late summer and fall in the Northern hemisphere (Jedel & Salmon, 1995), and in late winter and spring in the Southern hemisphere (Amigone et al., 2005). Moreover, frequent drought periods in semiarid regions result in a shortage of feed, increasing the demand for alternative feed sources to complement pastures that many times can not keep up the demand of cattle, which is gaining size and weight (Entz et al., 2002). In this context, exploration of new plant genotypes that may have the potential to grow as forage in dryland systems is valuable.
Annual forages are promoted as a source of feed supply because they can extend the grazing period or increase hay and silage production. However, annual crops differ in phenology, growth habit and forage quality and their selection should be based on intended, fnal use. Traditional cool-season winter and spring forage crops include wheat, rye, oats and barley. Synthetic cereals, such as triticale (X Triticosecale Wittmack) and tricepiro (X Triticosecale Wittmack x X Agrotriticum Ciferri & Giacom) can be added to this list. Triticale has outperformed traditional cereals in wide-ranging semiarid environments (McLeod et al., 1998; Stallknecht & Wichman, 1998), being well adapted to extreme cold, drought and acidic soils (Brown & Almodares, 1976; Yagmur & Kaydan, 2008). On the other hand, tricepiro, a newer cereal original from Argentina, has been studied locally in its agronomic traits, showing a valuable performance in marginal areas (Covas, 1989, 1995).
Tricepiro was obtained with the purpose to develop a new crop adapted to marginal areas by crossing a hexaploid triticale (2n=6x=42, AABBRR) with an octoploid trigopiro [(X Arotriticum Ciferri & Giacom); 2n=8x=56, AABBDDJJ] (Covas, 1989; 1995; Ferrari et al., 2005). Several years of self pollination in tricepiro resulted in a line that was registered as a cultivar under the name Tricepiro "Don René INTA" (Covas, 1989, 1995). This cultivar possesses high potential as a parent line in breeding programs due to its notable disease resistance and freezing and drought tolerance, combining desirable agronomic traits from its origin species (Covas, 1989, 1995). Tricepiro has a longer growing period than triticale, and better performance for tillering, dry matter yield, resistance to grain shattering, resistance to black rust, and salt tolerance. Furthermore, in comparison with trigopiro, tricepiro has greater initial growth rate, more active re-growth, higher dry matter yield, and it is easier to thresh (Covas, 1995).
Among the forage winter cereals, tricepiro present the longest cycle of production and high biomass yield either in semiarid (Frecentese & Covas, 1985; Ferri et al., 1995), sub-humid and humid environments (Gonella & Hernández, 1987, 1990; Brizuela & Cseh, 1996; Coraglio et al., 1998). According to Coraglio et al. (1998), tricepiro is stable in its biomass production, with a minimum yield of 3500 kg dry matter per hectare. Concomitantly, studies of biomass quality demonstrate high values of dry matter digestibility throughout its life cycle (75-80%), pointing out this cereal as a good candidate to be used as a forage reserve (Brizuela & Cseb, 1996).
Tricepiro was originally conceived as winter forage (Frecentese & Covas, 1985). However, its feature as a grain producer for human and animal feeding was also tested (García & Torresi, 1996; Gros et al., 1995). Preliminary studies of protein composition show a high level of lysine, suggesting a good value as a food, and open the avenue for further studies concerning amino acid composition and biological value of its proteins (Gros et al., 1995). Tricepiro grains have also been tested as a livestock food. When diary diet of pigs was primarily based on tricepiro grain, the dorsal fat was thinner compared to that based on diets rich in grains other than tricepiro (García & Torresi, 1996). Taken as a whole, tricepiro has a potential to be used in the food industry in addition to its current use as a forage cereal (Bertoni et al., 1995 a and b).
The use of physiological traits, in addition to agronomics ones, to evaluate the drought tolerance in winter cereals is well documented (Araus et al., 2002). Variables such as stomatal resistance (SR) (Santivelli Morata, 1993; Morant-Avice et al., 1994; Nogueira et al., 2000; Frank & Berdahl, 2001); water potential (WP) (Hanson et al., 1977 cited by García et al., 2002; Frank & Berdhal, 2001; Wery 2005); leaf area (LA) (Araus et al., 2002); growth of leaves (Shonfed et al., 1988; Van Loo, 1992; Cabuslay et al., 2002; García et al., 2002; Guenni et al., 2002), and roots (Grzesiak et al., 2001; Zhang et al., 2001; Araus et al., 2002) have been extensively used to better understand their response to water stress. For instance, genotypes of triticale have been discriminated by their response to drought using WP (Grzesiak et al., 2003, Grzesiak et al., 2006), and root growth (Grzesiak et al., 2001; Grzesiak et al., 2007) among others.
In cereals, from the beginning of agriculture, drought has been the main yield-limiting factor (Araus et al., 2002). Background information in triticale and cereals in general shows that it is possible to fnd water stress tolerance in these species (Oskan et al., 1999; Barary et al., 2003; Grzesiak et al., 2003). Performance of tricepiro in the field allows us to infer a good adaptation to water deficit conditions (Covas, 1989; Dreussi, 1998). However, the performance of tricepiro under controlled water stress conditions has not been evaluated to date. Therefore, in this study, we investigated the effect of water defcit in tricepiro cultivar "Don Rene" and the experimental lines L54, L62 and LFRC50 in comparison with their progenitors (triticale and trigopiro) in field and under greenhouse conditions using physiological (i.e. elongation and leaf area, water potential, stomatal resistance) and agronomical variables (i.e. biomass production, grain yield, and yield components).
MATERIALS AND METHODS
Tricepiro cultivar Don Rene INTA (DR), and the experimental lines 62, 54 and LF RC50 (L62, L54 and RC50), were tested in comparison with their progenitor's species trigopiro cultivar Don Noe INTA (DN) and triticale cultivar Don Santiago INTA (DS). Genotypes were provided by the Universidad Nacional de La Pampa (UNLPam), Argentina (L62, L54, DR, DN) and the Universidad Nacional de Río Cuarto (UNRC), Argentina (RC50). Two different trials were conducted, one in the greenhouse and the other in the field.
Greenhouse trial. Cultivars and experimental lines were grown in PVC pots (50 cm deep and 16 cm diameter with 9 kg of soil) in the greenhouse of the Faculty of Agronomy of UNLPam, Santa Rosa, La Pampa, Argentina (36° 34´ S, 64° 16´, 210 m.a.s.l.) from March 2004 to October 2004. Soil was composed by 59% sand, 34% silt and 7% clay, and it had 1.4% organic matter, 19.7 ppm phosphorous, pH 6.00, 12.1% of field capacity and 7.83% permanent wilting point (PWP). Figure 1 shows the relative humidity and average temperature values registered in the greenhouse during the study period. The photosynthetic photon flux density ranged from 500 to 1200 µm/m2/s.

Fig. 1. Average temperature (°C) and relative humidity (%) inside the greenhouse from seedling emergence onwards. Days from emergence (DFE).
Fig. 1. Temperatura media (°C) y humedad relativa (%) dentro del invernáculo desde la emergencia de la semilla. Días desde la emergencia (DDE).
A water stress treatment was imposed. Watering was withheld at the beginning of stem elongation (57 days after emergence) until water stress symptoms were evident (average leaf water potentials were -3 MPa and -0.4 MPa in the water stress and control treatments, respectively). After reaching this point, irrigated pots were re-watered to measure plant recovery to water stress. Controls were watered to field capacity every day. Pots were randomly arranged into a 2 x 6 (water x genotype) factorial design with 5 replicates.
After withholding water, stomatal resistance (SR; Delta T Porometer, AP4-UM-2, 2.28 Version, 1991) and leaf water potential (LWP; Scholander pressure chamber) were measured twice a week until the end of the experiment. At the same time, leaf growth on the youngest leaf of the main stem (L) from every plant was registered. L was used to calculate the average relative growth rate (RGR), RGR= [(lnL2 - lnL1) / (t2 - t1)], where L1 and L2 were leaf length at times 1 and 2 (t1 and t2) (Fitter & Hay, 1983). At the end of the experiment [55 days after withholding water (DAWW)], leaf area was measured using a LICOR LI 3000 leaf area meter. Shoot and root dry weights were determined and used to calculate specifc leaf area (SLA, leaf area/leaf weight), root/shoot ratio and wilted/green leaf weight (W/G).
Field trial. The study was conducted in the Agricultural Experiment Station Anguil "Ing. Agr. Guillermo Covas", La Pampa Argentina ( EEA Anguil, 36° 30´ S, 63° 59´ W), during 2005 and 2006; the soil was an entic haplustol with 41.86 ppm phosphorous and 0.12% nitrogen, field capacity 15.7% and PWP 11.6%. Weed control was applied both years (500 cc/ha 2.4D + 600 cc/ha Brominal). The experimental design was 2 x 6 (water x genotype) factorial design with 5 blocks in a split plot arrangement, where water treatment was the main plot and the genotypes the subplots. Sowing was performed on May 10, 2005 and April 21, 2006 at a density of 93 kg/ha. Plots (5 x 1.4 m) had row interspaces of 0.2 m. Plots had two water conditions, rain-feed and water irrigation (in addition to rainfall). Irrigation (35.7 mm each) was applied every 15 days; a total of 285.6 mm and 214.2 mm were added in 2005 and 2006, respectively. Rainfall during 2004 was 814 mm/year; monthly averages during 2005 and 2006 are shown in Table 1 (Meteorological Service, INTA EEA Anguil). Rainfall during the experimental period (highlighted in Table 1) was 199 and 178 mm in 2005 and 2006, respectively.
Table 1. Historical and 2005-2006 rainfall, number of days with frost (5 cm), mean and minimum temperatures during 2005 and 2006 in the site under study: National Institute of Agricultural Technology - Agricultural Experiment Station Anguil "Ing. Agr. Guillermo Covas", La Pampa, Argentina.
Tabla 1. Precipitaciones históricas y anuales, número de días con heladas (5 cm), temperaturas medias y mínimas durante 2005 y 2006 en el sitio bajo estudio: Instituto Nacional de Tecnología Agropecuaria, EEA Anguil -Ing. Agr. G. Covas, La Pampa, Argentina.

Distance between meteorological station and experimental site was 1 km. Months corresponding to the growing period of the experiment are shown in bold.
La distancia entre la estación meteorológica y el sitio experimental fue de 1 km. Los meses correspondientes al período de crecimiento en este estudio se muestran en negrita.
Measurements of plant height, SR and WP were made during the experiment as described earlier. Leaf area was determined (LICOR LI 3000) on leaves of plants from 0.5 m2 subplots during stem elongation. Measurements were conducted on October 11, 2005 (1544 DD) and September 27 (1453 DD in 2006), and leaf area index was also determined.
Afterwards, leaves were oven-dried at 60 °C to constant weight to calculate dry weight and specific leaf area (SLA; cm/g). The following forage quality determinations were made: acid detergent fiber (ADF, automatic analyzer Ankow, Goering & Van Soest, 1970), dry matter digestibility (DMD), metabolized energy (Mcal/kgDM), and crude protein (CP, Khjeldal, AOAC, 1990). Seed harvest was made when the grain moisture was 13%. A selection index was calculated using forage yield under control and water stress conditions in both years [(Ig1 =Yg1 -Ym/Ym)*100] where Ig1 is the index for each genotype, Yg1 is forage yield for each genotype, and Ym is the average forage yield of all genotypes (Badiali, 2008).
Data were statistically analyzed by ANOVA and LSD test (p<0.05). Pearson correlation analysis between yield and morpho-physiological variables, both under field and greenhouse conditions, was performed using InfoStat (2003).
RESULTS
In this study we evaluated morphological, physiological and agronomical variables under water stress in tricepiro cultivar "Don Rene" and the experimental lines L54, L62 and RC50 in comparison with their progenitors (triticale and trigopiro) under greenhouse and field conditions (2005 and 2006). In this section, results are organized per variable, and results from both greenhouse and feld experiments are presented jointly. In field experiments, variance analysis was performed. Triple and double interactions are presented in Table 2.
Table 2. Analysis of Variance in field experiments (2005 and 2006).
Tabla 2. Análisis de la Varianza en experimentos de campo (2005 y 2006).

* p<0.05, (-) Not significant. H: plant height; SR: Stomatal Resistance; LAI: Leaf area index; SLA: Specific leaf area. Y: year; G: genotype; W: water condition.
* p<0,05, (-) No significativo. H: altura de planta; SR: Resistencia Estomática; LAI: Índice de área foliar; SLA: Área foliar específica. Y: año; G: genotipo; W: condición hídrica.
Leaf water potential. In the greenhouse, differences in water potential (i.e., irrigation vs. water stress) were evident 19 days after withholding water. However, differences among genotypes during water stress and after recovering were not evident. Figure 2 shows LWP mean values for all genotypes exposed to each water condition.
Fig. 2. Water potential (MPa) of water stressed (WS) and control plants (C; irrigation). Results are the mean of the evaluated genotypes (tricepiro, triticale and trigopiro) in a greenhouse trial. Date of the rewatering after water stress treatment: 53 DAWW. DAWW: days after withholding water.
Fig. 2. Potencial agua (MPa) en plantas expuestas a estrés hídrico (WS) y control (C, regadas). Resultados representan la media de los genotipos evaluados (tricepiro, triticale and trigopiro) en un ensayo en invernáculo. Días luego de haber suspendido el suministro de agua.
Stomatal resistance. In the greenhouse experiment, well watered plants showed significant differences compared to stressed plants from 14 DAWW onwards in all the evaluated genotypes. The interaction W x G was significant (p<0.05) and differences among genotypes were also significant between 30 and 45 days. In this period, Trigopiro DN showed the greatest stomatal resistance (p<0.05), while the other cultivars did not differ (Table 3 shows results at 35 DAWW). Significant differences among cultivars were found the first day after re-watering plants (Table 3). On the first day after re-watering, there was a G x W interaction and differences among genotypes were registered. Trigopiro DN showed the greatest values of stomatal resistance, and tricepiro DR the lowest (p<0.05). Besides, RC50 showed the slower recovering after withholding water. On the second day after re-watering, we could not detect any significant differences among cultivars (data not shown).
Table 3. Stomatal resistance (SR, s/cm) of one cultivar (DR) and experimental lines of tricepiro (L62, L54 y RC 50), triticale (DS) and trigopiro (DN) on irrigated (C) and water stress (WS) plants.
Tabla 3. Resistencia estomática (SR, s/cm) de un cultivar (DR) y líneas experimentales de tricepiro (L62, L54 y RC 50), triticale (DS) y trigopiro (DN) en plantas con riego (C) y con estrés hídrico (WS).

*Data collected at 35 days DAWW.
In a column, means with the same letter are not statistically different (LSD, p<0.05). ns: not significant.
*Datos recolectados 35 días después de haber suspendido el suministro de agua.
Promedios con la misma letra en una columna no son signifcativamente diferentes (LSD, p<0,05). ns: no significativo.
In field experiments under irrigation, genotypes did not show differences among them either 2005 or 2006. However, in rainfeed plots, trigopiro and tricepiro RC50 showed the highest SR value compared with the other cultivars (p<0.05; Table 3).
Leaf area, leaf area index and leaf relative growth rate. In the greenhouse experiment, water stress reduced leaf area (p<0.01; Table 4) and differences among genotypes were also evident (p<0.05). However, a signifficant interaction W x G was detected. This reduction was quantifed as leaf area reduction (LAR: Leaf area water stress / Leaf area irrigation; Table 4), and it showed signifcant differences among genotypes. Trigopiro DN showed the highest reduction, as well as triticale and tricepiro L 54. Remaining tricepiro genotypes showed higher leaf area than their progenitors under water stress.
Table 4. Leaf area per plant (LA, cm2) and Leaf area reduction (WS/C, %), Specific leaf area (SLA, cm2/g), specific leaf area reduction (WS/C, %) and Wilted/Green Leaf proportion of one cultivar (DR) and experimental lines of tricepiro (L62, L54 y RC 50), triticale (DS) and trigopiro (DN) under water stress (WS) and control (C). Results from a representative experiment are shown.
Tabla 4. Área foliar por planta (LA, cm2) y reducción del área foliar (WS/C, %), Área foliar específica (SLA, cm2/g), reducción del área foliar específica (WS/C, %) y proporción de hoja verde/hoja seca en un cultivar (DR) y líneas experimentales (L62, L54 y RC 50), triticale (DS) y trigopiro (DN) bajo estrés hídrico (WS) y controles (C). Se muestran resultados de un experimento representativo.

Within columns, values with the same letter are not statistically different (LSD, p<0.05). ns: not significant.
Dentro de columnas, valores con la misma letra no son estadísticamente significativos (LSD, p<0,05). ns: no significativo.
Water deficit significantly reduced the specific leaf area (SLA; p<0.01; Table 4) and differences were found among cultivars (p<0.01); differences were not evident in watered plants. Under water stress DR showed the greatest SLA but did not differ from L62, L54 and DN.
Water stress induced a significant increase in senescence, measured as wilted/green (W/G) leaves proportion (p<0.01). This effect was more evident in Trigopiro DN and Triticale DS, while tricepiro L62 was the less afected (Table 4). In addition, the interaction W x G was not significant.
Regarding the leaf relative growth rate (LRGR), cultivars decreased this parameter after 42 days under water stress. Triticale and trigopiro highly decreased LRGR under water stress (i.e. WS/C = 37 and 51%, respectively) in comparison with tricepiro (Fig. 3).

Fig. 3. Leaf Relative growth rate (LRGR) of one cultivar (DR) and experimental lines of tricepiro (L62, L54 and RC50), triticale (DS) and trigopiro (DN), cumulative in a 42-day-period (LSD, p<0.05 performed on WS/C). WS: water stress, C: control.
Fig. 3. Tasa de crecimiento relativa (LRGR) de un cultivar (DR) y líneas experimentales de tricepiro (L62, L54 and RC50), triticale (DS) y trigopiro (DN), acumulado en un periodo de 42 días (LSD, p<0,05 realizado en WS/C). WS: estrés hídrico, C: control.
In field experiments (2005 and 2006), leaf area index (LAI) did not show significant differences in rainfeed versus irrigated cultivars (WS/C), indicating that genotypes were affected by water stress to a similar extent. The decrease of LAI, as an average of genotypes, was 36 in 2005 and 39% in 2006. In 2005, the specific leaf area was significantly reduced between water levels (p<0.05, mean values were 186 and 173 cm2/g for C and WS, respectively), although no differences were found among genotypes; the interaction W x G was not significant. In 2006, no signifcant differences were detected either between water levels or among genotypes.
Plant height. In field experiments, Trigopiro DN showed small plant height (p<0.05) in both years and water conditions. Analyzing plant height for each cultivar compared with the control (WS/C; %), no significant differences were found in 2005, suggesting a similar response of all genotypes under rainfeed conditions. However, significant differences were found in 2006 (p<0.05) where plant height of tricepiros RC50, DR and trigopiro DN were less affected compared to the control (p<0.05, Fig. 4).

Fig. 4. Plant height (cm) under irrigation (C), water deficit (WS) and WS/C of one cultivar (DR) and experimental lines of tricepiro (L62, L54 y RC50), triticale (DS) and trigopiro (DN), during 2005 and 2006. Differences in 2005 were not significant. LSD p<0.05.
Fig.4. Altura de planta (cm) bajo riego (C), sin riego (WS) y WS/C de un cultivar (DR) y líneas experimentales de tricepiro (L62, L54 y RC50), triticale (DS) y trigopiro (DN), durante 2005 y 2006. Diferencias en el 2005 no fueron significativas. LSD p<0,05.
Aerial, underground biomass, aerial/underground ratio. In greenhouse experiments, water stress significantly reduced (p<0.01) biomass. The interaction W x G was not significant. Genotypes did not differ among them either under water stress or under well watered conditions (Table 5). However, biomass yield (i.e. WS/C, %) showed significant differences among cultivars under water stress in comparison with controls (p<0.05). Tricepiro RC50 and L62 showed a small decrease in aerial biomass due to water stress while tricepiro DR was the most affected cultivar.
Table 5. Above and underground biomass in greenhouse and field experiments of one cultivar (DR) and experimental lines of tricepiro (DR, L62, L54 y RC 50), triticale (DS) and trigopiro (DN), exposed to irrigation (C) and water stress conditions (WS).
Tabla 5. Biomasa aérea y radicular en experimentos de campo y de invernáculo de un cultivar (DR) y líneas experimentales (L62, L54 y RC 50), triticale (DS) y trigopiro (DN) y trigopiro (DN) bajo riego (C) y estrés hídrico (WS).

* Plots were rainfeed.
Within columns, means with the same letter are not statistically different (LSD, p<0.05).
* Las parcelas fueron expuestas a lluvia natural.
Dentro de las columnas, promedios con la misma letra no son estadísticamente diferentes (LSD, p<0,05).
In field experiments, trigopiro showed the lowest biomass production under irrigation and rainfeed conditions in 2005 and 2006. In contrast, triticale showed the greatest biomass yield, while tricepiro lines had an intermediate performance (Table 5). Reduction in dry matter production due to water stress was, on average, 31 and 49% in 2005 and 2006 respectively, but there were no differences among cultivars.
Regarding root biomass, the interaction W x G and the genotype effect were both significant (p<0.05 and 0.01, respectively). Stressed plants had a lower root biomass than control plants, except for tricepiro RC50 which did not show differences (p<0.01). Under the evaluated conditions, parental genotypes triticale DS and trigopiro DN had smaller root dry matter and WS/C than tricepiro materials (Table 5).
Forage quality. In field experiments, water stress did not affect forage quality variables; mean values are presented in Table 6. In turn, differences in crude protein (p<0.05) among genotypes were detected. Tricepiro RC50 had the highest protein percentage under watering and rainfeed conditions (i.e. 14.32 and 12.97%, respectively), while triticale had the lowest (i.e., C = 9.33 and WS = 8.52%). Tricepiro DR had higher protein content under rainfeed than watering conditions (i.e., 12.57 vs 9.52%); this could be the result of a dilution efect that did not show up in other genotypes.
Table 6. Average forage quality of one cultivar and experimental lines of tricepiro, triticale and trigopiro under irrigated (C) and rainfeed (WS) conditions.
Tabla 6. Calidad de forraje promedio de un cultivar y líneas experimentales de tricepiro, triticale y trigopiro con riego (C) y sin riego (WS).

NDF: neutral detergent fiber; ADF: acid detergent fiber; DMD: dry matter digestibility; ME: metabolic energy; CP: crude protein.
NDF: fibra detergente neutro; ADF: fibra detergente ácido; DMD: digestibilidad de la material seca; ME: energía metabólica; CP: proteína cruda.
Grain yield. Seed production in field plots did not show a W x G interaction either in 2005 or 2006. Water stress had a significant effect, showing on average a yield reduction of 37% and 48% in 2006 and 2005, respectively (p<0.01). In 2005, Triticale had a superior seed production (Table 7). In 2006, although variance analysis did not show significant differences among varieties, triticale had a superior seed production.
Table 7. Seed production (SP, kg/ha) of one cultivar and experimental lines of tricepiro (DR, L62, L54 y RC 50), triticale (DS) and trigopiro (DN) under irrigated (C) and rainfeed (WS) conditions.
Tabla 7. Producción (SP, kg/ha) de un cultivar y líneas experimentales de tricepiro (DR, L62, L54 y RC 50) triticale (DS) y trigopiro con riego (C) y sin riego (WS).
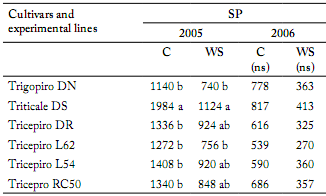
Within columns, means with the same letter are not statistically different (LSD, p<0.05); ns: not significant.
Dentro de las columnas, promedios con la misma letra no son significativamente diferentes (LSD, p<0,05); ns: no significativo.
Correlation between forage yield and morpho-physiological variables. Pearson correlation analysis was performed between biomass and the morpho-physiological variables, both in the greenhouse and at feld, to identify variables which showed a close relationship with yield. Biomass was associated significantly to water potential (r = -0.46; p<0.01), stomatal resistance (r = 0.42; p<0.02), root biomass (r = 0.40; p<0.03) and wilted/green ratio (r = 0.41; p<0.02). Under field conditions, forage yield was highly correlated to leaf area (r = 0.64 and 0.68; p<0.001) and plant height (r = 0.37; p<0.05) in the two study years. This last variable is of relatively simple determination; hence this could be useful for screening genotipes in breeding programs.
DISCUSSION
We studied the water stress response of one cultivar and three experimental lines of tricepiro in comparison with their progenitors triticale and trigopiro. We found that genotypes of tricepiro and triticale performed similarly under drought both in the greenhouse and at the field; however, tricepiro was superior to trigopiro. Overall, among tricepiros, L62 and the line RC50 were the best performing genotypes. We evaluated morpho-physiological variables that might be related to yield under water deficit, which could be useful to select superior genotypes in breeding programs (Araus et al., 2002, 2008). In the greenhouse, forage yield had a high correlation with root biomass (p<0.001) both under control and water-stress conditions. Under water stress, WP and RE significantly correlated with forage yield (p<0.01 and 0.02, respectively) as well as the ratio wilted/green (p<0.05). In the field, forage yield had a fne association (p<0.05) with plant height both under control and water-stress conditions in 2005 and 2006. Leaf area, on the other hand, was highly related to yield during stress (p<0.01) but not under well irrigated conditions.
There is currently a strong support toward an approach that the understanding of the crop at the physiological and molecular biology level might help discover the key traits that are currently limiting yield. Such approach may therefore complement the conventional breeding programs and accelerate yield improvements. Analyzing physiological determinants of yield responses to water availability may help in breeding for higher yield and stability under drought conditions (Araus et al., 2002). The identification of one or more physiological parameters as indices of drought resistance has been the subject of extensive research works (Karamanos, 1984; Karamanos & Papatheohari, 1999). Today, several variables have been proposed to screening genotypes (Araus et al., 2008).
The relevance of water to maintain turgor in plant tissues is well known, and, in many genotypes, a high water potential in leaves is indicative of a higher drought tolerance. This is the case in cereals such as triticale, barley, rice, wheat and other grasses (Frank & Berdhal, 2001; García et al., 2002; Grzesiak et al., 2003; Grzesiak et al., 2006). In tricepiro and its progenitors, differences in WP of plants under water stress and controls were found both in the greenhouse and field experiments, although differences among genotypes were not significant. Recovering after watering of stressed plants was similar for all genotypes even after two days of re-watering (Table 3). In wheat, Shonfeld et al. (1988) found a decrease in WP in plants under stress but they did not find differences among genotypes. In triticale accessions, Santivelli Morata (1993) did not fund diferences either in WP or relative water content. In triticale cultivars, Grezsiak et al. (2003) reported that, under drought, changes of WP, fluorescence and leaf injury, which might be suitable for screening of drought tolerance.
Stomatal resistance has been used as a criterion in different species to select drought tolerant plants (Morant-Avice et al., 1994; Nogueira et al., 2000; Frank & Berdhal, 2001; Araus et al., 2002; Wery, 2005). In our study, we found similar results in the evaluated genotypes both in field and greenhouse experiments. Trigopiro DN showed the greatest increase in stomatal resistance under water stress and lower recovering in stomatal conductance after re-watering. Divergence in stomatal conductance during recovering was found in other cereals such as wheat. In wild and cultivated genotypes of Triticum, Shimshi et al. (1982) observed that recovery was fastest in T. longissimum, an annual species of semiarid to sub-humid habitats, somewhat slower in T. kotsch and much slower in bread wheat. Fischer et al. (1998) in bread wheat found that high stomatal conductance was associated with higher yield. This plant response could explain the lower yield in trigopiro compared to the other accessions.
Leaf area index in field experiments showed that a limitation in water content affected tricepiro lines and its progenitors to similar extent. However, at the greenhouse, tricepiro showed a greater leaf area compared to trigopiro and triticale, and a lower reduction of leaf area in comparison with controls. The divergence between field and greenhouse experiments could be explained as a result of the more severe water stress at the greenhouse than at the field (i.e., -3 MPa and -1.23 MPa under greenhouse and feld conditions, respectively).
A higher leaf area per unit of mass has resulted in a high rate of potential growth under non-stress conditions, but, in turn, it has been a disadvantage under stressful conditions (Sugiyama, 2006). Under water stress, the specific leaf area was reduced both at the field and the greenhouse (p<0.05) with no significant differences among genotypes, indicating a similar response to water stress in the evaluated materials.
One of the plant responses to water deficit is the increase in leaf senescence to reduce the transpiration rate (Araus et al., 2002). However, this feature can compromise plant yield. In this study, significant differences in leaf senescence between well-watered and stressed plants were found, similar to other reports in the literature (Maroco et al., 2000; Cabuslay et al., 2002). Water stress did not affect all genotypes to the same extent. While triticale was the most affected (WS/C= 362%), L62 was the less affected (WS/C= 92%).
Reduction in leaf growth as a result of water stress is well documented in grasses (Van Loo, 1992; Cabuslay et al., 2002; García et al., 2002). In our study, leaf growth rate (LGR) was severely reduced under water stress conditions in triticale and trigopiro (WS/C = 37 and 51%, respectively) under greenhouse conditions. In tricepiro genotypes, in turn, leaf growth rate decreased more slightly under water stress (ranging from 60 to 95% WS/C). This could be associated with the higher leaf area found in tricepiro materials under water stress (Table 4).
An important root growth is a feature to be considered in semiarid environments, since plants roots can reach deeper layers of soils to contribute to plant yield stability (Pugnaire et al., 1994). In our greenhouse experiments, tricepiro RC50 showed the highest underground biomass under stress while trigopiro and triticale had the lowest root biomasses (p<0.05). Grzesiak et al. (2001, 2007) observed that, in triticale, root biomass was more affected in sensitive genotypes under water stress. The same observation was made by Araus et al. (2002) in cereals. In this matter, it is important to note the limitation of greenhouse experiments since root growth in pots does not strictly compare with root growth under field conditions.
When water stress is imposed slowly, as is generally the case under field conditions, a reduction in C assimilation and utilization may occur along with restrictions in CO2 assimilation due to stomatal closure when drought progresses. Concomitantly, there is a proportional reduction in the activity of various enzymes of the Calvin cycle (Maroco et al., 2002). Under water stress conditions, forage production is also affected because the leaf middle life is shorter and pastures are less dense, with a reduced numbers of tillers and green leaves per tiller (Colabelli et al., 1998). In our field experiments, water stress affected forage production, being trigopiro DN the cultivar with lowest forage yield both in 2005 and 2006. However, differences among cultivars under water stress with respect to the control were no evident in both years. Hence, divergences among cultivars seem to be inherent to each genotype instead of an effect of water stress. Water limitation in field experiments was moderate (i.e. -1.23 MPa), and this could be the reason why differences were not evident among genotypes. In greenhouse experiments, where the water stress was severe (i.e., -3 MPa), divergences among genotypes were evident.
To better explain the performance of the genotypes, we calculated a standardized index (higher values indicate better performance) in both years and watering conditions (Badiali, 2008), and plotted control vs. stress (Fig. 5). In this plot, trigopiro located in the lower left quadrant, an indication of its poor performance under both control and stress conditions in 2005 and 2006. Contrary to this, triticale and tricepiro L62 positions in the upper right quadrant showed a superior performance in all situations; among the other tricepiro genotypes the index was intermediate.

Fig. 5. Biomass index of one cultivar and experimental lines of tricepiro (DR, L62, L54 and RC50), triticale (DS) and trigopiro (DN) in Water Stress versus Control, during 2005 and 2006.
Fig. 5. Indice de biomasa de un cultivar y líneas experimentales (DR, L62, L54 y RC 50), triticale (DS) y trigopiro (DN) expuestas a Estrés hídrico versus Control, durante 2005 y 2006.
Analyzing the effect of water stress on biomass reduction in the greenhouse experiment, we observed that it was smaller in RC50 and L62 compared to the other tricepiro genotypes (Table 5). RC50 did not show a reduction in root biomass, which could contribute to a better water capture. On the other hand, L62 showed the lower leaf senescence under water stress, maintaining a longer photosynthetic capacity. These two variables (i.e. root biomass and W/G) had a high correlation with biomass; hence they could be major contributors to yield. In the case of RC50, the biomass yield in the control was low; however, under water stress it showed a good performance. Thus, this line could be suitable for environments with frequent drought situations. On the other hand, L62 showed a good performance both under control and stress conditions, appearing as promissory among the tricepiro lines for both water availability conditions. Overall, it is noticeable the high variability of biomass reduction among tricepiro lines, opening an avenue for future selection.
Forage quality (NDF, ADF, DMD, CP, ME) of well-watered and rainfed plants did not differ, except for crude protein (CP). In this case, divergences in the genotypes were evident both under watered and rainfeed conditions. Tricepiro RC50 had the highest values and triticale had the lowest. Regarding ADF, NDF, DMD and ME, our results differ from other authors. In some perennial summer grasses, it has been shown that water stress increased DMD and nitrogen levels in leaves (Guenni et al., 2002). In Russian rye (Psathyrostachys juncea), water stress increased protein levels and decreased ADF and NDF (Karn et al., 2004). However, this increase in digestibility could be the efect of delaying the stem elongation, flowering and leaf ontogenic changes (Wilson & Ng, 1975).
Grain production of tricepiro lines was intermediate in comparison with their progenitors. Triticale had the highest crop index and grain yield only during 2005. Low production of tricepiro lines could be related to a low fertility natural from interspecific crosses and, additionally, to the lower seed weight. Decreasing in grain yield as a consequence of water stress was similar among cultivars.
Interspecific and intergeneric crosses have been shown to be successful in some aspects - such as seed production, quality, disease resistance- and triticale is a clear example (Santivelli Morata, 1993). The intergeneric cross of triticale x trigopiro showed to be superior in terms of palatability, growing cycle, regrowth capacity and disease resistance (Covas, 1995). In terms of water stress tolerance, the aim of this work, tricepiro had a good performance, similar to triticale. In addition, we found morphophysiological variables that highly correlated with biomass production. In the greenhouse, water potential, stomatal resistance, root biomass and wilted/green were close associated with forage, under a considerable water stress (- 3 MPa). In the field (mild-moderate stress: -1.23 MPa), forage yield was higly related to plant height and leaf area in the two year trials. These physiological traits could be evaluated as potential indicators that might be useful for screening tricepiro genotypes in breeding programs. There is considerable variability among the evaluated tricepiro genotypes to allow the selection of drought tolerant materials. Besides, future tricepiro breading plans might consider the use of parental lines with drought tolerance background in order to improve water stress tolerance.
REFERENCES
1. Amigone, M.A., A.M. Kloster, C. Navarro & N. Bertram (2005). Elección de cultivares e implantación de verdeos de invierno. In: Verdeos de alta producción para optimizar la cadena forrajera. Información para Extensión N° 96, pp. 5-14. EEA Marcos Juárez, INTA. [ Links ]
2. AOAC (1990). Official methods of analysis (15th Ed.) Association of Official Analytical Chemists. Washington, DC. [ Links ]
3. Araus, J.L., G.A. Slafer, C. Royo & M.D. Serret (2008). Breeding for Yield Potential and Stress Adaptation in Cereals. Critical Reviews in Plant Science 27: 377-412. [ Links ]
4. Araus, J.L., G.A. Slafer, M.P. Reynolds & C. Royo (2002). Plant breeding and drought in C3 cereals: what should we breed for? Annals of Botany 89: 925-940. [ Links ]
5. Badiali, O.J. (2008) Selección de genotipos de triticales hexaploide. Congreso Nacional de Trigo. Santa Rosa, La Pampa. [ Links ]
6. Barary, M., N.W.M. Warwick, R.S. Jessop & A.M. Taji (2003). Variation for osmotic adjustment in Autralian triticale cultivars. Proceedings of Australian Agronomy Conference, Australian Society of Agronomy. 4 p. [ Links ]
7. Bertoni, M.H., A. Pereyra González, P. Cattaneo & G. Covas (1995a). Composición química de especies naturales y sintéticas de Triticeas (Gramineae). Tricepiros I. Características de los granos y de los aceites crudos de extracción. Anales de la Asociación Química Argentina 83: 153-158. [ Links ]
8. Bertoni, M.H., A. Pereyra González, P. Cattaneo & G. Covas (1995b). Composición química de especies naturales y sintéticas de Triticeas (Gramineae). Tricepiros II. Harinas de extracción de cereales sintéticos. Anales de la Asociación Química Argentina 83:159-163. [ Links ]
9. Brizuela, M.A. & S.B. Cseh (1996). Niveles de macrominerales en tricepiro bajo dos frecuencias de defoliación. Revista Argentina de Producción Animal 16 Sup. 1: 237-238. [ Links ]
10. Brown, A.R. & A. Almodares (1976). Quantity and quality of triticale forage compared to other small grains. Agronomy Journal 68: 264-266. [ Links ]
11. Cabuslay, G.S., O. Ito & A.A. Alejar (2002). Physiological evaluation of responses of rice (Oriza sativa L.) to water deficit. Plant Science 163: 815-827. [ Links ]
12. Clapham, W.M. & J.M. Fedders (2008). Effect of sowing date of triticale on seasonal herbage production in the central Appalachian Highlands of the United States. Grass Forage Science 63: 447-457. [ Links ]
13. Colabelli M., M. Agnusdei, A. Mazzanti & M. Labreveux (1998). El proceso de crecimiento y desarrollo de gramíneas forrajeras como base para el manejo de la defoliación. Boletín Técnico Nº 148. INTA EEA Balcarce. Centro regional Buenos Aires Sur. 21 p. [ Links ]
14. Coraglio, J.C., C.A. Vieyra, F. Casanoves & L. Capón (1998). Estabilidad de la producción de triticale, trigopiro y tricepiro en el centro de la provincia de Córdoba. Revista Argentina de Producción Animal 18 Sup.1: 132. [ Links ]
15. Covas, G. (1989). Pampa semiárida. Nuevos cultivos. Ciencia Hoy 1: 75-77. [ Links ]
16. Covas, G. 1995. Tricepiro Don René INTA: un verdeo muy productivo. Horizonte Agropecuario 35: 6-7. [ Links ]
17. Dreussi, L.W. (1998). Características de algunos cultivares obtenidos en la Estación Experimental Agropecuaria Anguil. Boletín de Divulgación Técnica Nº 54, 18-21. [ Links ]
18. Entz, M.H., V.S. Baron, P.M. Carrc, D.W. Meyerd, S.R. Smith Jr. & W.P. McCaughey (2002). Potential of forages to diversify cropping systems in the Northern Great Plains. Agronomy Journal 94: 240-250. [ Links ]
19. Ferrari, M.R., E.J. Greizerstein, H.A. Paccapelo, C.A. Naranjo, A. Cuadrado, N. Jouve & L. Poggio (2005). The genomic composition of Tricepiro, a synthetic forage crop. Genome 48: 154-159. [ Links ]
20. Ferri, C.M., O.A. Hernández & M.A. Frecentese (1995). Comportamiento de verdeos invernales en santa Rosa la Pampa I. Distribución estacional y rendimientos acumulados de materia seca. Revista de la Facultad de Agronomía UNLPam 8: 1-9. [ Links ]
21. Fische, R.A. & R. Maurer (1978). Drought resistance in spring wheat cultivars. I. Grain yield responses. Australian Journal of Agronomy Research 29: 897-912. [ Links ]
22. Fischer, R.A., D. Rees, K.D. Sayre, Z.M. Lu, A.G. Condon & A. Larque-Saavedra (1998). Wheat yield progress associated with higher stomatal conductance and photosynthetic rate and cooler canopies. Crop Science 38: 1467-1475. [ Links ]
23. Fitter, A.H. & R.K.M. Hay (1983). Environmental physiology of plants. Academic Press, London, UK. 355 p. [ Links ]
24. Frecentese, M. & G. Covas (1985). Comportamiento de nuevos verdeos en la región semiárida pampeana. Informativo de Tecnología Agropecuaria para la Región Semiárida Pampeana 84: 6-7. [ Links ]
25. Frank, A.B. & J.D. Berdhal (2001). Gas exchange and water relations in diploid and tetraploid russian wildrye. Crop Science 41: 87-92. [ Links ]
26. García, G. & L. Torresi (1996). Utilización de tricepiro en dietas de crecimiento y engorde de cerdos. Trabajo Final de Graduación para la obtención del título de Ingeniero Agrónomo. Revista de la Facultad de Agronomia UNLPam. 16 p. [ Links ]
27. García, M.G., C.A. Busso, P. Polci, N.L. García Girou & V. Echenique (2002). Water relations and leaf growth rate of tree Agro- pyron genotypes under water stress. Biocell 26: 309-317. [ Links ]
28. Goering, H.K. & P.J. Van Soest (1970). Forage fiber analysis (apparatus, regents, procedures and some applications). In: Agriculture Handbook, USDA Washington, DC. 379 pp. 1-20. [ Links ]
29. Gonella, C.A. & R.A. Hernández (1987). Resultados técnicoeconómico de la unidad modelo demostrativa de invernada. Boletín Técnico Informativo para la Región Pampeana Subhúmeda. EEA INTA Gral. Villegas 5: 1-7. [ Links ]
30. Gonella, C.A. & R.A. Hernández (1990). Evaluación de verdeos invernales. Boletín Técnico Informativo para la Región Pampeana Subhúmeda. EEA INTA Gral. Villegas 8: 1-4. [ Links ]
31. Gros, E.G., G.M. Caballero, M.H. Bertoni, P. Cattaneo & G. Covas (1995). Composición química de especies naturales y sintéticas de Triticeae (Gramínea). Tricepiros III. Estudio comparativo de esteroides en aceites puros de extracción. Anales de la Asociación Química Argentina 83: 165-169. [ Links ]
32. Grzesiak, S., M.J. Grzesiak, W. Filek, J. Stabyla, T. Hura & S. Pienkowski (2001). Genotypic variation in the growth of the root system of triticosecale cultivars under water stress conditions. Abstracts of the 4th International Conference, Rackovadolina, Slovakia. September 12-14, 2000. JCEA 2(1-2). [ Links ]
33. Grzesiak, S., M.T. Grzesiak, W. Filek & J. Stabryla (2003). Evaluation of physiological screening tests for breeding drought resistant triticale (x Triticosecale wittmack). Acta Physiologia Plantarum 25: 29-37. [ Links ]
34. Grzesiak, M.T., S. Grzesiak & A. Skoczowski (2006). Changes o leaf water potential and gas exchange during and after drought in triticale and maize genotypes difering in drought. Photosynthetica 44: 561-568. [ Links ]
35. Grzesiak, M.T., A. Rzepka, T. Hura, K. Hura & M. Skoczowski (2007). Changes in response to drought stress of triticale and maize genotypes differing in drought tolerance. Photosynthetica 45: 280-287. [ Links ]
36. Guenni, O., D. Marín & Z. Baruch (2002). Responses to drought of five Brachiaria species. Biomass production, leaf growth, root distribution, water use and forage quality. Plant Soil 243: 229-241. [ Links ]
37. InfoStat (2003). InfoStat versión 1.5. Manual del usuario. Grupo InfoStat, FCA, Universidad Nacional de Córdoba. 1 ed., Editorial Brujas, Argentina. 336 p. [ Links ]
38. Jedel, P.E. & D.F. Salmon (1995). Forage potential of spring and winter cereal mixtures in a short-season growing area. Agronomy Journal 87: 731-736. [ Links ]
39. Juskiw, P.E., D.F. Salmon & J.H. Helm (1999). Annual forage production from spring-planted winter cereal monocrops and mixtures with spring barley. Canadian Journal of Plant Science 79: 565-577. [ Links ]
40. Karamanos, A.J. (1984). Ways of detecting adaptive responses of cultivated plants to drought. In: N.S. Margaris et al. (ed.) Being alive on land. Tasks for vegetation science. pp. 91-101. Dr. W. Junk, The Hague. [ Links ]
41. Karamanos, A.J. & A.Y. Papatheohari (1999). Assessment of drought resistance of crop genotypes by means of the water potential index. Crop Science 39:1792-1797. [ Links ]
42. Karn, J.F., A.B. Frank, J.D. Berdahl & W.W. Poland (2004). Ploidy, water and nitrogen effects on Russian wildrye chemical composition. Rangeland Ecology and Management 57: 503-510. [ Links ]
43. Badiali, O.J. (2008). Selección de Genotipos de Triticale Hexaploide. VII Congreso Nacional de Trigo - V Simposio de Cereales de Siembra Otoño-Invernal - I Encuentro del MERCOSUR - [ Links ]
44. Maroco, J.P., J.S. Pereira & M.M. Chaves (2000). Growth, photosynthesis and water -use efficiency of two C4 Sahelian grasses subjected to water deficits. Journal of Arid Environment 45: 119- 137. [ Links ]
45. McColoy, Q.W., L.W. Sherrod, R.C. Albin & K.R. Hansen (1971). Nutritive value of triticale for ruminants. Journal of Animal Science 32: 534-539. [ Links ]
46. McLeod, J.G., Y.T. Gan, D.F. Salmon & V.S. Baron (1998). Triticale Biomass potential and quality on the Canadian prairies. In: P.E. Juskiw (ed.). Proc. Int. Triticale Symp., 4th, Red Deer, AB, Canada. 26-31 July 1998. pp. 264-267. Vol. 2. Agric. and AgricFood Can., Lacombe, AB. [ Links ]
47. Morant-Avice A., P. Jurvillies & A. Coudret (1994). Stomatal movements and gas exchanges of a triticale and its parental species in water stress conditions. Agronomie 14: 113-120. [ Links ]
48. Nielsen D.C., M.F. Vigil & J.G. Benjamin (2006). Forage yield response to water use for dryland corn, millet, and triticale in the Central Great Plains. Agronomy Journal 98: 992-998. [ Links ]
49. Nogueira, R.J.M.C., J.F. da Silva Júnior, J.E.F. Becerra, I.E. Lederman, H.A. Burity & V.F. dos Santos (2000). Comportamiento estomático y tensión de agua en el xilema de dos genotipos de pitanga (Eugenia uniflora L.) cultivados bajo estrés hídrico. Investigación Agraria en Protección Vegetal 15: 214-225. [ Links ]
50. Oskam H., I. Genc, T. Yagbasanlar & F. Toklu (1999). Stress tolerance in hexaploid spring triticale under Mediterranean environment. Short Communication. Plant Breeding 118: 365. [ Links ]
51. Pugnaire, F.I., L. Serrano Endolz & J. Pardos (1994). Constraints by water stress on plant growth. Cap. 11. In: Persakli, M. University of Arizona. Tucson, Arizona. EEUU. pp. 247-259. [ Links ]
52. Santivelli Morata, P. (1993). Influencia del hábito de crecimiento sobre el comportamiento agronómico y fisiológico del triticale hexaploide (x Triticosecale, Wittmack). Servei de Publications Universitat de Leida pp.35-54. [ Links ]
53. Schonfeld, M.A., R.C. Johnson, B.F. Carver & D.W. Mornhinweg (1988). Water relations in winter wheat as drought resistance indicators. Crop Science 28: 526-531. [ Links ]
54. Shimshi, D., M.L. Mayoral & D. Atsmon (1982). Responses to water stress in wheat and related wild species. Crop Science 22: 123-128. [ Links ]
55. Stallknecht, G.F. & D.M. Wichman (1998). The evaluation of winter and spring triticale (Triticosecale Wittmack) for grain and forage production under dryland cropping in Montana, USA. In: P.E. Juskiw (ed.) Proc. Int. Triticale Symp., 4th, Red Deer, AB Canada. 26-31 July 1998. pp. 272-278. Vol. 2. Agric. and AgricFood Can., Lacombe, AB. [ Links ]
56. Sugiyama, S. (2006). Responses of shoot growth and survival to water stress gradient in diploid and tetraploid populations of Lolium multiforum and L. perenne. Grassland Science 52:155-160. [ Links ]
57. Van Loo, E.N. (1992). Tillering, leaf expansion and growth of plants of two cultivars of perennial ryegrass grown using hydroponics at two water potentials. Annals of Botany 70: 511-518. [ Links ]
58. Wery, J (2005). Differential effects of soil water deficit on the basic plant functions and their significance to analyses crop responses to water deficit in indeterminate plants. Australian Journal of Agronomy Research 56: 1201-1209. [ Links ]
59. Wilson, J.R. & T.T. Ng (1975). Influence of water stress on parameters associated with herbage quality of Panicum maximum var. trichoglume. Australian Journal of Agronomy Research 26: 127-136. [ Links ]
60. Yagmur, M. & D. Kaydan (2008). Early seedling growth and relative water content of triticale varieties under osmotic stress of water and NaCl. Research Journal of Agriculture and Biological Science 4: 767-772. [ Links ]
61. Zhang, X.G., R.S. Jessop, F. Ellison & P. Evans (2001). Diferential responses to water regimes in triticale genotypes differing in aluminum tolerance. Proceedings of the Australian Agronomy Conference, Australian Society of Agronomy pp.1-8. [ Links ]

