










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkPhyton (Buenos Aires)
On-line version ISSN 1851-5657
Phyton (B. Aires) vol.85 no.1 Vicente López June 2016
ARTÍCULOS ORIGINALES
Diferenciación geográfica de poblaciones de chile silvestre (Capsicum annuum L. var. glabriusculum) del noroeste de México
Geographical diferentiation of wild pepper (Capsicum annuum L. var. glabriusculum) populations from northwestern Mexico
López-España RG1, S Hernández-Verdugo1, S Parra-Terraza1, F Porras1, Antonio Pacheco-Olvera1, A Valdez-Ortiz2, T Osuna-Enciso3, MD Muy-Rangel3
1 Facultad de Agronomía, Universidad Autónoma de Sinaloa (UAS), Carr. Culiacán-El Dorado Km 17.5. Tel. y Fax 01 (667) 846-1084. Apdo. Postal 726. Culiacán Sinaloa, México.
2 Facultad de Ciencias Químico Biológicas de la Universidad Autónoma de Sinaloa.
3 Centro de Investigación en Alimentación y Desarrollo A. C. Unidad Culiacán. Culiacán, Sinaloa, México.
Address correspondence to: Sergio Hernández Verdugo, e-mail: sergioh2002mx@yahoo.com.mx
Received 8.IV.2015.
Accepted 14.VII.2015.
Resumen. El chile silvestre (Capsicum annuum L. var. glabriusculum) es un valioso recurso genético para la agricultura y la alimentación. Los patrones de variación de 12 caracteres morfo-demográficos de 17 poblaciones de chile silvestre colectadas a lo largo de un gradiente latitudinal y climático en el noroeste de México fueron analizados en condiciones uniformes de invernadero. Los datos de dichas características se examinaron con análisis univariados y multivariados. La relación entre la variación fenotípica y la latitud de origen de las poblaciones se probó mediante análisis de regresión lineal. Se encontró elevada variación dentro y entre las poblaciones en todas las características medidas. Los análisis de varianza univariados y de componentes principales diferenciaron claramente a las poblaciones. Debido a que las plantas crecieron en un ambiente uniforme, estos resultados indican que las diferencias entre las poblaciones se deben a diferencias genéticas entre ellas. Las características altura de la planta, número de frutos, peso del fruto, número de semillas por fruto y número de semillas por planta mostraron un patrón de variación clinal a lo largo del gradiente latitudinal. Las plantas de las poblaciones del norte del gradiente fueron de menor altura, produjeron más frutos, frutos de mayor peso, más semillas por fruto y más semillas por planta que las plantas de las poblaciones del sur. El patrón de variación observado en estas características sugiere adaptación a las condiciones climáticas locales. La variación de las demás características (diámetro del tallo, largo y ancho de la hoja, número de ramas, largo y ancho del fruto y peso de la semilla) no se relacionó significativamente con la latitud de origen. Es posible que la diferenciación observada en estas características sea un resultado de regímenes de selección diferentes (y no clinales) en cada población, o una consecuencia de la deriva génica.
Palabras clave: Capsicum annuum silvestre; Recursos genéticos; Gradiente latitudinal; Diferenciación entre poblaciones; Variación genética.
Abstract. Wild pepper (Capsicum annuum L. var. glabriusculum) is a valuable genetic resource for food and agriculture. The patterns of variation of 12 morphological traits of 17 populations of wild pepper collected across a latitudinal and climate gradients in northwestern Mexico were analyzed in a uniform greenhouse experiment. The morphological traits data were axamined by univariate and multivarite analysis. The relationship between phenotypic variation and latitude of population origen was tested using linear regression analysis. We found high variation within and among populations for most of the measured traits. Populations were clearly differentiated by univariate analysis of variance and principal component analysis. Because plants grew in a uniform environment, these results indicate that the differences among the populations were due to genetic differences among them. Plant height, number of fruits, fruit weight, number of seeds per fruit, number of seeds per plant showed a clinal pattern of variation across latitudinal gradients. Plants from northern populations were shorter, produced more fruits, fruits were heavier, more seeds per fruit and more seeds per plant than plants from southern populations. The variation pattern observed in these traits suggests adaptation to local climate conditions. The variation of the other traits (stem diameter, leaf length and width, number of branches, fruit length and width and seed weight) was not significantly related to latitude de origin. It is possible that the observed differentiation in these traits was the result of different regimens of selection (and nonclinal) in each population, or a consequence of genetic drift.
Keywords: Wild Capsicum annuum; Genetic resource; Latitudinal gradient; Population differentiation; Genetic variation.
INTRODUCCIÓN
La mayoría de las especies vegetales mantienen variación genética intraespecífica en caracteres morfológicos, demográficos y fisiológicos que les permite adaptarse a las condiciones fluctuantes del ambiente. Esta variación genética dentro de las especies es la base de su sobrevivencia y evolución en condiciones naturales, y para el mejoramiento de las plantas cultivadas por el ser humano. La conservación de esta variación genética es de gran importancia para las estrategias de mantenimiento, uso y manejo de los recursos genéticos presentes en las especies vegetales para la agricultura y la alimentación (Food and Agriculture Organization of the United Nations, 2010).
Los factores que afectan la distribución y abundancia de las poblaciones a escala geográfica incluye la manera en que los organismos se relacionan con el ambiente. El clima es considerado uno de los principales factores que explican los patrones de distribución y variación de muchas especies vegetales, ya sea que actúe directamente sobre los procesos fisiológicos durante el crecimiento y la reproducción o indirectamente a través de otros factores ecológicos tales como la competencia por recursos (Nooryazdan et al., 2010).
La latitud representa un gradiente ambiental a lo largo del cual varían en una manera continua varios factores del clima como la temperatura, la precipitación y la radiación solar (Li et al., 1998; De Frenne et al., 2013). La variación en características morfológicas, demográficas, fenológicas y fisiológicas a lo largo de gradientes latitudinales ha sido investigada extensivamente entre poblaciones de plantas (Li et al., 1998; Jonas y Geber, 1999; Olsson y Ågren, 2002; Hall et al., 2007; De Frenne et al., 2013). Cuando la variación fenotípica o genética se correlaciona con los factores geográficos y climáticos de los sitios de origen de las poblaciones, se considera que es en respuesta a las diferentes presiones de selección locales (Charlesworth y Charlesworth, 2010).
Los estudios sobre la variación en caracteres fenotípicos y sus patrones de distribución geográfica pueden proporcionar información importante sobre los procesos evolutivos y contribuir a una mejor comprensión de los mecanismos de diversificación entre y dentro de las especies (Nattero et al., 2011). La estructura de la variación genética puede diferir entre los caracteres estudiados, dependiendo si éstos están sujetos o no a la selección, y de cómo la selección varía espacialmente (Olsson y Ågren, 2002).
La diferenciación entre las poblaciones puede ser promovida principalmente por los procesos de selección natural y la deriva génica al azar (Wright, 1951). La selección natural opera en aquellas poblaciones donde sus individuos presentan diferencias entre sí, y cuando tales diferencias son heredables y tienen un valor adaptativo; en cambio, la deriva génica produce cambios aleatorios independientes de las condiciones locales (Kimura y Otha, 1971).
Los parientes silvestres de las plantas cultivadas son un recurso genético importante que constituye un acervo de genes primario que puede ayudar a resolver problemas de la agricultura actual, tales como tolerancia o resistencia a plagas y enfermedades, y aumentar la calidad y cantidad de la producción (Burdon y Jarosz, 1989). México es uno de los países con mayor diversidad vegetal y uno de los principales centros de domesticación de plantas en el mundo (Harlan, 1971). En particular el chile (Capsicum spp.) fue una de las primeras plantas domesticadas en el Continente Americano (MacNeish, 1964).
El género Capsicum (Solanacea) está conformado por alrededor de 30 especies distribuidas desde el sur de los Estados Unidos, hasta el norte de Argentina (Pickersgill, 1984; Hernández-Verdugo et al., 1999). Del género han sido domesticadas Capsicum annuum L., Capsicum chinense Jacq., Capsicumfrutescens L., Capsicum baccatum L. y Capsicum pubescens Ruiz y Pavón (Pickersgill, 1984). Se considera que C. annuum L. ha sido domesticada en México (Pickersgill, 1984), y de todas las especies domesticadas es la de mayor importancia económica y la que presenta mayor variabilidad en tamaño, forma y color de sus frutos. A ella pertenecen los chiles “de árbol” o “cola de rata”, “anchos”, “serranos”, “jalapeños” y “morrón”, entre otros.
Las plantas de chile silvestre (Capsicum annuum L. var glabriusculum (Dunal) Heiser y Pickersgill), conocidas comúnmente como chiles “chiltepines” o “piquines” son perennes, herbáceas o trepadoras, sus frutos son pequeños, rojos y picantes; son comidos por las aves que dispersan sus semillas. Capsicum annuum silvestre tiene una amplia distribución, desde el sur de los Estados Unidos hasta las regiones da baja altitud del Perú (Pickergill, 1971). En México, las poblaciones de chile silvestre (C. annuum) se distribuyen ampliamente en todo el territorio nacional. Se localizan principalmente en sitios no perturbados de la selva baja caducifolia, pero es posible encontrarlas a orillas de los caminos, huertos, potreros y bajo la vegetación remanente de los campos de cultivo (Hernández-Verdugo et al., 1999). En el noroeste de México, esta especie florece en los meses de julio y agosto, y sus frutos maduran en octubre y noviembre.
Estudios previos con isoenzimas, marcadores moleculares RAPDs y microsatélites han mostrado que las poblaciones de C. annuum silvestre del noroeste de México mantienen altos niveles de variación genética dentro y entre sus poblaciones (Hernández-Verdugo et al., 2001a; Oyama et al., 2006; Pacheco-Olvera et al., 2012). Estas poblaciones también variaron significativamente en la resistencia a geminivirus (Hernández-Verdugo et al., 2001b) y en la capacidad de germinación de las semillas (Hernández-Verdugo et al., 2001c, 2010). En un estudio reciente, Hernández-Verdugo et al. (2012) encontraron que las poblaciones de C. annuum silvestre del noroeste de México variaron significativamente en caracteres morfológicos medidos en condiciones naturales. Sin embargo, los estudios sobre la variación fenotípica en poblaciones de plantas que crecen en condiciones naturales no permiten distinguir los efectos genotípicos de los efectos del ambiente. Para ello se requiere la comparación en un ambiente uniforme de las poblaciones de C. annuum silvestre colectadas en sitios diferentes. Por lo anterior, el objetivo del presente estudio fue determinar los patrones de diferenciación fenotípica en un ambiente uniforme de invernadero de poblaciones de Capsicum annuum silvestre colectadas a lo largo de un gradiente latitudinal en el noroeste de México.
MATERIALES Y MÉTODOS
Material vegetal y sitios de colecta. Se recolectaron frutos maduros de 20 a 40 plantas de 17 poblaciones (Tabla 1) de chile silvestre distribuidas en un gradiente latitudinal en el noroeste de México (Fig. 1). La latitud cubrió un intervalo de 23° 00’ 41” hasta 29° 55’ 59” latitud Norte (Tabla 1, Fig. 1). Las longitudes estuvieron en un intervalo de 105° 47’ 41” a 109° 49’ 48” longitud Oeste. La altitud varió entre 87 y 582 m.s.n.m. La precipitación y la temperatura media anual estuvieron entre 384 y 1138 mm, y 21,1 y 25,7 °C, respectivamente (Tabla 1). La temperatura y la precipitación media anual disminuyeron significativamente con el aumento de la latitud (y = 38,133 – 0,530 x; R2 = 0,526, F = 16,6554, P<0,001 para la temperatura media anual; y = 2367,04 – 62,46 x; R2 = 0,306, F = 6,6035, P=0,0213, para la precipitación media anual) (Fig. 2).
Tabla 1. Datos geográficos y climáticos de 17 poblaciones de Capsicum annuum silvestre estudiadas en México.
Table 1. Geographic and climatic data of the collection sites of the 17 wild Capsicum annuum populations studied in Mexico.
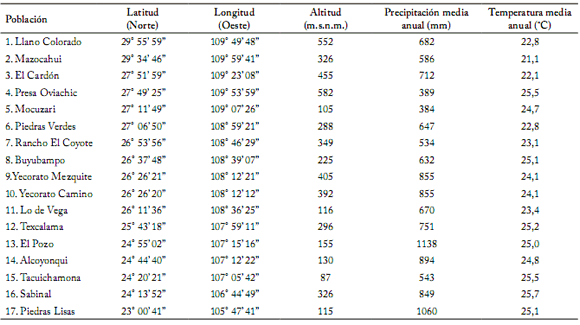

Fig. 1. Localización de las 17 poblaciones de Capsicum annuum silvestre recolectadas en el noroeste de México. Los números corresponden al número de población de la Tabla 1.
Fig. 1. Location of the 17 wild Capsicum annuum populations collected in northwestern Mexico. Numbers correspond to the population number from Table 1.

Fig. 2. Relación entre la latitud y la precipitación media anual (A) y la temperatura media anual (B) de los sitios de origen de las poblaciones de Capsicum annuum silvestre estudiadas.
Fig. 2. Relationships between latitude and mean annual precipitation (A) and mean annual temperature (B) of the origin sites of the wild Capsicum annuum studied populations.
Diseño experimental. Las semillas de las diferentes plantas de cada población se trataron con 1000 ppm de ácido giberélico durante 24 h y se pusieron a germinar en sustratoórgano-mineral (“tierralite”) en bandejas de poliestireno. Después de 30 días de la germinación se trasplantaron en promedio 35 plantas por población en macetas con 12 kg de suelo de aluvión en un diseño experimental completamente al azar con respecto a la población. Los trasplantes se dispusieron en un invernadero bajo malla-sombra que permitió el paso de 60% de luz solar (con malla-sombra fabricada por Textiles Agrícolas, Celaya, Guanajuato, México). Las plantas fueron regadas mediante riego por goteo y no fueron fertilizadas. Las plantas de cada población fueron rotadas cada 30 días durante los primeros cuatro meses del experimento para evitar la heterogeneidad del ambiente en el interior del invernadero (Fornoni y Núñez-Farfán, 2002).
Características medidas. Seis meses después del trasplante se evaluaron los siguientes caracteres: (1) altura de la planta (cm), (2) diámetro en la base del tallo (mm), (3) longitud y (4) ancho de la hoja (mm, tres hojas por planta en la parte media de la planta), (5) número de ramas sobre el tallo principal, (6) número de frutos (número total de frutos al final de la estación), (7) longitud y (8) ancho del fruto (mm), (9) peso del fruto (mg), (10) número de semillas por fruto, (11) número de semillas por planta y (12) peso de la semilla (mg). Las características longitud y ancho del fruto, peso y número de semillas por fruto se obtuvieron de una muestra de diez frutos por planta. El peso de la semilla se obtuvo pesando individualmente cinco semillas de cinco frutos de cada planta. Los frutos y las semillas se pesaron en una balanza analítica (Sartorius), con un margen de precisión de 0,1 mg.
Análisis estadístico. Los datos morfo-demográficos se analizaron mediante análisis de varianza univariados y análisis multivariados de componentes principales. Cuando las diferencias de los análisis univariados fueron significativas (P<0,05) se hicieron pruebas múltiples de medias (Tukey-Kramer). Se estimaron la media, desviación estándar y coeficiente de variación. Para mejorar la normalidad de los datos, las características número de frutos y número de semillas por planta se transformaron en sus valores logarítmicos. Los patrones de diferenciación morfológica entre las poblaciones, se examinaron mediante análisis de regresión lineal entre las medias de las poblaciones para cada característica y las latitudes de sus sitios de origen. Los análisis se efectuaron con el paquete estadístico JMP 6.0 de SAS.
RESULTADOS
Estadísticos descriptivos. Considerando todas las poblaciones juntas, los estadísticos descriptivos mostraron elevada variación en la mayoría de las características analizadas (Tabla 2). La altura de la planta varió de 53 a 249 cm. El diámetro del tallo estuvo en un intervalo de 5,9 hasta 20,7 mm. La longitud y ancho de la hoja variaron de 12,5 a 79,3 y 5,2 a 58,1 mm, respectivamente. Los valores mínimos y máximos del número de ramas fueron 2 y 5, respectivamente. El número de frutos varió ampliamente, desde 1 hasta 199. En los frutos, la longitud y el ancho tuvieron valores mínimos y máximos de 2,7 a 9,4 mm y 2,4 a 7,2 mm, respectivamente. El peso de fruto varió ampliamente, desde 2,8 hasta 130,6 mg. El número de semillas por fruto varió ampliamente, desde 1 hasta 25. El peso individual de las semillas varío desde 0,9 hasta 5,5 mg. La característica número de semillas por planta presentó una amplia variación, desde 2,0 hasta 1377 (Tabla 2).
Tabla 2. Estadísticos descriptivos de 12 características analizadas en 17 poblaciones de Capsicum annuum silvestre de México.
Table 2. Descriptive statistics of 12 traits measured on 17 populations of wild Capsicum annuum from Mexico.
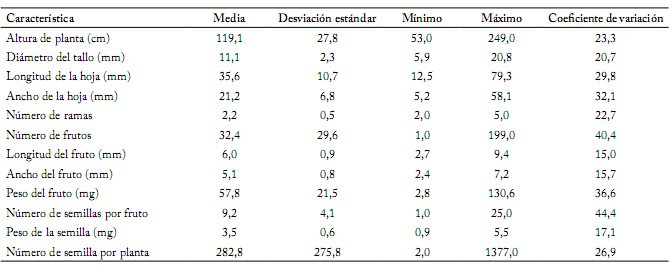
Los coeficientes de variación (CV) variaron de 15,0% (longitud del fruto) a 44,4% (número de semillas por fruto) con una media de 27,1%. Las características reproductivas número de semillas por fruto y número de frutos mostraron CV superiores al 40%, mientras que el peso del fruto, ancho de la hoja y longitud de la hoja tuvieron CV cercanos o superiores al 30%. Sólo las características longitud del fruto, ancho del fruto y peso de la semilla tuvieron CV alrededor de 15% de variación (Tabla 2).
Diferenciación morfológica entre poblaciones. El análisis de varianza de una vía mostró que las poblaciones de Capsicum annuum silvestre se diferenciaron significativamente en todas las características evaluadas (Fig. 3).
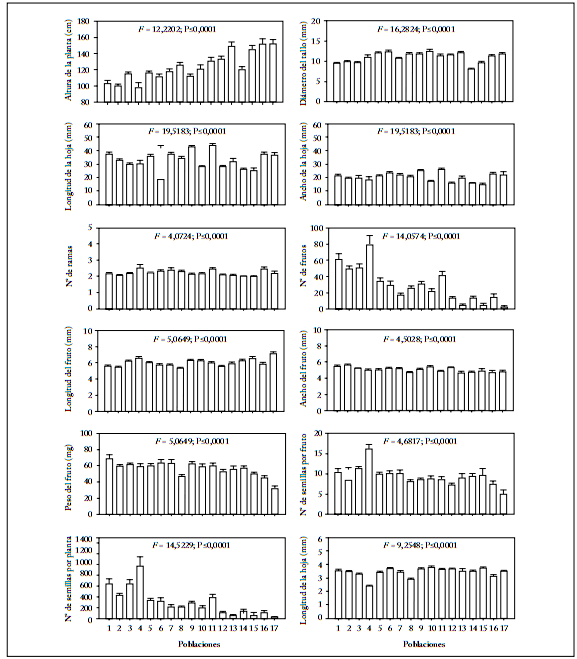
Fig. 3. Medias (±1 error estándar) de 12 características analizadas en 17 poblaciones silvestres de Capsicum annuum del noroeste de México. Se indica los resultados del análisis de varianza de una vía (F y P). Los números de cada población corresponden a los nombres de la Tabla 1.
Fig. 3. Means (±1 standard error) for the 12 traits measured in 17 wild populations of Capsicum annuum from northwestern Mexico. Results of one way ANOVAs are indicated (F and P). Numbers of each population correspond to names given in Table 1.
El análisis de componentes principales mostró que los primeros tres componentes principales explicaron 75,97% de la variación (Tabla 3). El primer componente principal explicó 36,34% de la variación y fue definido principalmente por las características número de semillas por planta, número de semillas por fruto, número de frutos y peso de los frutos, con signo positivo y altura de la planta con signo negativo. El segundo componente principal explicó 23,11% y estuvo determinado por las características longitud de la hoja, ancho de la hoja y número de ramas, todas con signo positivo. El tercer componente principal explicó 16,56% de la variación y estuvo determinado principalmente por las características peso de la semilla y ancho del fruto, con signo positivo (Tabla 3). Las poblaciones se diferenciaron claramente en el espacio bidimensional de los componentes principales 1 y 2 (Fig. 4). El componente principal 1 distinguió a las poblaciones 4 (Presa Oviachic), 1 (Llano Colorado) y 3 (El Cardón) en la región de plantas con menor altura, mayor número de frutos, mayor número de semillas por fruto, mayor número de semillas planta y mayor peso del fruto, mientras que las poblaciones 17 (Piedras Lisas), 16 (Sabinal) y 13 (El Pozo) ocuparon la región opuesta (Fig. 4, Tabla 3). El componente principal 2 colocó a la población 11 (Lo de Vega) en la región de plantas con mayor longitud y ancho de la hoja y mayor número de ramas. En cambio, la población 15 (Tacuichamona) ocupó la región opuesta (Fig. 4, Tabla 3).
Tabla 3. Resultado del análisis de componentes principales de 12 características analizadas en 17 poblaciones de Capsicum annuum silvestre. Las variables con mayor peso están en negritas.
Table 3. Results of the principal component analysis of 12 traits measured on 17 wild Capsicum annuum populations. Most important variables are in boldface.


Fig. 4. Diferenciación de 17 poblaciones de Capsicum annuum silvestre sobre los componentes principales 1 y 2. Los números corresponden a las poblaciones de la Tabla 1.
Fig. 4. Differentiation of the 17 wild Capsicum annuum populations on the principal components 1 and 2. Numbers correspond to the populations from Table 1.
Relación entre la variación morfo-demográfica y la latitud de los sitios de recolecta. Los análisis de regresión indicaron que las características altura de la planta, número de frutos, peso de los frutos, número de semillas por fruto y número de semillas por planta variaron clinalmente con la latitud de los sitios de origen de las poblaciones (Fig. 5). Las plantas fueron de menor altura (intervalo de variación de 92,3 cm en la población 4 a 167,5 cm en la población 17) (Fig. 5A), con menor número de frutos (intervalo de variación de 3,0 en la población 17 a 79,1 en la población 4) (Fig. 5B), frutos con menor peso (intervalo de variación de 31,2 mg en la población 17 a 68,4 mg en la población 1) (Fig. 5C), menor número de semillas por fruto (intervalo de variación de 5,0 en la población 17 a 16,1 en la población 4) (Fig. 5D), y menor número de semillas por planta (intervalo de variación de 18,7 en la población 17 a 966,6 en la población 4) en la parte norte del gradiente (Fig. 5E). Las demás características no se correlacionaron significativamente con la latitud.

Fig. 5. Variación latitudinal en las características altura de la planta (A), número de frutos (B), peso del fruto (C), número de semillas por fruto (D) y número de semillas por planta (E) en 17 poblaciones de Capsicum annuum silvestre ( media ± 1 error estándar).
Fig. 5. Latitudinal variation in the traits plant height (A), fruit number (B), fruit weight (C), number of seeds per fruit (C), number of seeds per plant (D) on 17 wild Capsicum annuum populations (mean ± 1 standard error).
DISCUSIÓN
Los resultados de este estudio indican que las poblaciones silvestres de Capsicum annuum del noroeste de México mantienen elevados niveles de variación entre y dentro de sus poblaciones. De las 12 características medidas, dos mostraron CV superiores a 40% y otras dos presentaron CV mayores a 30%. El CV promedio de las doce características fue 27,1% y sólo tres características (longitud del fruto, ancho del fruto y peso de la semilla) mostraron CV menores a 20%. Otros investigadores han considerado que CV superiores a 20% indican la presencia de variación elevada dentro de poblaciones o especies vegetales útiles para la agricultura y la alimentación. Adebola y Morakinyo (2006), Lewu et al. (2007) y Nooryazdan et al. (2010) interpretaron los CV mayores a 20% como la presencia de una amplia variación dentro de poblaciones y especies de los géneros Cola, Polorgonium y Heliantus, respectivamente.
Los análisis de varianza de una vía y el multivariado de componentes principales diferenciaron claramente a las poblaciones estudiadas. Estos resultados coinciden con la variación genética elevada entre y dentro de las poblaciones de C. annuum silvestre del noroeste de México estimada con isoenzimas, RAPDs y microsatélites (Hernández-Verdugo et al., 2001a; Oyama et al., 2006; Pacheco-Olvera et al., 2012) y en caracteres cuantitativos medidos en condiciones comunes de invernadero (Hernández-Verdugo et al., 2008). Estos resultados coinciden también con los estudios isoenzimáticos (Hamrick y Godt, 1997), con RAPDs (Bussell, 1999) y con caracteres fenotípicos (Rice y Mack, 1991), que han indicado que las especies vegetales con autofecundación muestran diferenciación significativa entre sus poblaciones. Debido a que las plantas de este experimento crecieron en un ambiente uniforme, los resultados indican que las diferencias entre las poblaciones de C. annuum silvestre estudiadas son debido a diferencias genéticas entre ellas.
La variación de las características altura de la planta, número de frutos, peso del fruto, número de semillas por fruto y número de semillas por planta se correlacionó significativamente con la latitud. Las poblaciones colectadas en los sitios de mayores latitudes, presentaron las plantas de menor altura, con mayor número de frutos, frutos de mayor peso, mayor número de semillas por fruto y mayor número de semillas por planta. El patrón de variación observado en estas características sugiere adaptación a las condiciones locales. La latitud se ha correlacionado con la temperatura, precipitación y con la radiación solar (Li et al., 1998; Jonas y Geber, 1999; De Frenne et al., 2013). En este estudio la temperatura media anual y la precipitación media anual disminuyeron aproximadamente de manera lineal con el aumento de la latitud (Fig. 2), sugiriendo que el patrón de diferenciación geográfico observado es una expresión indirecta de un gradiente climático de los sitios de origen de las poblaciones de C. annuum silvestre.
Otros investigadores han propuesto que las diferencias fenotípicas entre poblaciones de plantas de la misma especie que crecen en un ambiente uniforme y muestran patrones clinales de diferenciación geográfica, se deben a la selección natural que actúa a lo largo de gradientes ambientales (Hall, et al., 2007; Nooryazdan et al., 2010). En un experimento de invernadero, Hall et al. (2007) encontraron que la diferenciación en varias características fenotípicas de 12 poblaciones de Populus tremula siguieron un patrón clinal latitudinal. Ellos concluyeron que la diferenciación entre las poblaciones de Populus tremula son adaptaciones dirigidas por las condiciones climáticas locales. En un estudio en condiciones de invernadero, Nooryazdan et al. (2010) reportaron que la diferenciación en características morfológicas de 77 poblaciones de Heliantus annus silvestre colectadas en los Estados Unidos se correlacionó con las variables climáticas de los sitios de origen de las poblaciones, sugiriendo adaptación a las condiciones locales.
En nuestro estudio, las plantas de las poblaciones del norte del gradiente latitudinal fueron de menor altura. Este patrón de variación en el tamaño de las plantas a lo largo de un gradiente latitudinal coincide con el reportado en varias especies vegetales, como en Verbascum thapsus (Reinartz, 1984) Arabidopsis thaliana (Li et al., 1998), Lythrum salicaria (Olsson y Ågren, 2002). El patrón de variación en altura de la planta en las poblaciones de C. annuum silvestre relacionado con la latitud, presumiblemente es un resultado de la diferenciación genética en respuesta a las condiciones locales. Por ejemplo, la temperatura local es determinada en gran parte por la latitud (Li et al., 1998; De Frenne et al., 2013), por lo que las áreas de bajas latitudes, más cercanas al ecuador, tienden a tener mayores temperaturas (Fig. 2), las cuales aumentan las actividades metabólicas, el crecimiento celular, la fotosíntesis, y en consecuencia el crecimiento de la planta (Li et al., 1998).
En este estudio las plantas de las poblaciones del norte del gradiente latitudinal fueron de menor altura, pero produjeron un mayor número de frutos, mayor número de semillas por fruto y número de semillas por planta. Esto puede ser debido a la existencia de un conflicto en la asignación de recursos entre la reproducción y el crecimiento vegetativo en las plantas (Snow y Whigham, 1989; Olsson y Ågren, 2002). Estos resultados coinciden con los reportados en otras especies de plantas como Podophylum peltatum (Sohn y Policansky, 1977), Poa annua (Law, 1979), Asclepias cuadrifolia (Chaplin y Walker, 1982), Tipularia discolor (Snow y Whigham, 1989) y Lythrum salicaria (Olsson y Ågren, 2002), donde el aumento en la producción de flores y frutos condujo a una reducción significativa en el tamaño de las plantas. En nuestro estudio la altura de la planta se correlacionó negativa y significativamente con número de frutos, peso del fruto, número de semillas por fruto y número de semillas por planta (r promedio = -0,684, P<0,01). Esta relación inversa entre estas característica puede cancelar la selección que actúa independientemente sobre ellas, constituyendo una limitación potencial para alcanzar valores óptimos (Scheiner y Berrigan 1998).
En contraste con los patrones de variación entre las poblaciones en las características altura de planta, número de frutos, peso del fruto, número de semillas por fruto y número de semillas por planta, la variación de las características diámetro del tallo, longitud y ancho de la hoja, número de ramas, longitud y ancho del fruto, y peso de la semilla no mostraron patrones latitudinales. Es posible que la variación observada en estas características sea un resultado de regímenes de selección diferentes (y no clinales) que ha conducido a cada población a su óptimo local (Wright, 1951), o sea producto de la deriva génica que genera cambios aleatorios independientes de las condiciones locales (Kimura y Otha, 1971).
Los resultados de este estudio difirieron de los reportados previamente en estas poblaciones creciendo en condiciones naturales (Hernández-Verdugo et al., 2012), quienes no encontraron relación entre la altura de la planta y el gradiente latitudinal, mientras que las características peso del fruto y número de semillas por fruto se relacionaron positivamente con la precipitación y la temperatura media anual. Las diferencias en los resultados obtenidos en este estudio con los de Hernández-Verdugo et al. (2012) pueden ser debido a que en condiciones uniformes de crecimiento de las plantas, la diferenciación observada entre las poblaciones es considerada principalmente adaptativa, mientras que en condiciones naturales, la diferenciación entre poblaciones de la misma especie puede ser producto de una respuesta plástica a la heterogeneidad del ambiente (De Frenne et al., 2013; Castillo et. al., 2014). Hernández-Verdugo et al. (2015) encontraron elevados niveles de plasticidad fenotípica en poblaciones de C. annuum silvestre del noroeste de México en respuesta a diferentes niveles de luz. La elevada variación genética, junto con la elevada plasticidad fenotípica y en la germinación de sus semillas (Hernández-Verdugo et al., 2001a, 2001c, 2010), pueden ser mecanismos que le permiten a C. annuum silvestre vivir en diferentes hábitats, tales como sitios perturbados y no perturbados de la selva baja caducifolia, y tener la amplia distribución geográfica actual en el Continente Americano.
AGRADECIMIENTOS
A la Universidad Autónoma de Sinaloa (proyecto PROFAPI 2014/087) y al CONACYT (proyecto 106129) por el apoyo financiero otorgado a este estudio. A Heriberto Luna Barraza por su colaboración en la colecta de material biológico.
REFERENCIAS
1. Adebola, P.O. y J.A. Morakinyo (2006). Evaluation of morpho-agronomic variability of wild and cultivated kola (Cola species Schott et Endl.) in South Western Nigeria. Genetic Resources and Crop Evolution 53: 687-694. [ Links ]
2. Bussell, D. (1999) The distribution of random amplified polymorphic DNA (RAPD) diversity amongst populations of Isotomapetreae (Lobeliaceae). Molecular Ecology 88: 775-89. [ Links ]
3. Burdon, J.J. y A.M. Jarosz. (1989). Wild relatives as sources of disease resistance. En: Brown A.H.D., O.H. Frankel, D. R. Marshall y J.T. Williams (eds.) pp. 280-296. The use of plant genetic resources, Cambridge University Press. [ Links ]
4. Castillo, J.M., B.J. Grewell, A. Pickart, A. Bortolus, C. Peña, E. Figueroa y M. Sytsma (2014). Phenotypic plasticity of invasive Spartina densiflora (Poaceae) along a broad latitudinal gradient on the Pacific Coast of North America. American Journal of Botany 101: 448-458. [ Links ]
5. Chaplin, S.J. y J.L. Walker (1982). Energy constraints and adaptive significance of the floral display of a forest milkweed. Ecology 63: 1857-1870. [ Links ]
6. Charlesworth, B. y D. Charlesworth (2010). Elements of Evolutionary Genetics. Roberts and Company Publishers. Colorado, USA. [ Links ]
7. De Frenne, P., B.J. Graae, F. Rodríguez-Sánchez, A. Kolb, O. Chabrerie, G. Decocq, H. De Kort y A.D. Schrijver (2013). Latitudinal Gradients as natural laboratories to infer species’ responses to temperature. Journal of Ecology 101: 784-795.
8. Food and Agriculture Organization of the United Nations (2010). The second report on the state of the world’s plant genetic resources for food and agriculture. Commission of the genetic resources for food and agriculture, Roma, Italia 370 p.
9. Fornoni, J. y J. Núnez-Farfán (2002). Evolutionary ecology of Datura stramonium: genetic variation and costs for tolerance to defoliation. Evolution 54: 789-797. [ Links ]
10. Hall, D., V. Luquez, V.M. Garcia, K.R. St Onge, S. Jansson y P.K. Ingvarsson (2007). Adaptive population diferentiation in phenology across latitudinal gradient in European aspen (Populus tremula, L.): a comparison of neutral markers, candidate genes and phenotypic traits. Evolution 61: 2849-2860. [ Links ]
11. Hamrick, J.T. y M.J.W. Godt (1997). Allozyme diversity in cultivated crops. Crop Sciences 37: 26-30. [ Links ]
12. Harlan J. R. (1971). Agricultural origins: centers and no centers. Science 174: 468-474. [ Links ]
13. Hernández-Verdugo, S., P. Dávila-Aranda y K. Oyama (1999). Síntesis del conocimiento taxonómico, origen y domesticación del género Capsicum. Boletín de la Sociedad Botánica de México 64: 65-84. [ Links ]
14. Hernández-Verdugo, S., R. Luna-Reyes, y K. Oyama (2001a). Genetic structure and diferentiation of wild and domesticated populations of Capsicum annuum from Mexico. Plant Systematics and evolution 226: 129-142. [ Links ]
15. Hernández-Verdugo S., R. G. Guevara-González, R.F. Rivera-Bustamante y K. Oyama (2001b). Screening wild plants of Capsicum annuum for resistance to pepper huasteco virus: presence of viral DNA and diferentiation among populations. Euphytica 122: 31-36. [ Links ]
16. Hernández-Verdugo, S., K. Oyama y C. Vázquez-Yanes (2001c). Diferentiation in seed germination among populations of Capsicum annuum along a latitudinal gradient in Mexico. Plant Ecology 155: 245-257. [ Links ]
17. Hernández-Verdugo, S., R.G. López-España, P. Sánchez-Peña, M. Villarreal-Romero, S. Parra-Terraza, F. Porras y J.L. Corrales-Madrid (2008). Variación fenotípica entre y dentro de poblaciones silvestres de chile del noroeste de México. Revista Fitotecnia Mexicana 31: 323-330. [ Links ]
18. Hernández-Verdugo, S., R.G. López-España, F. Porras, S. Parra-Terraza, M. Villarreal-Romero y T. Osuna-Enciso (2010). Variación en la germinación entre poblaciones y plantas de chile silvestre. Agrociencia 44: 667-677. [ Links ]
19. Hernández-Verdugo, S., F. Porras, R.G. López-España, M. Villarreal-Romero, S. Parra-Terraza y T. Osuna-Enciso (2012). Caracterización y variación ecogeográfica de poblaciones de chile silvestre del noroeste de México. Polibotánica 33: 175-191. [ Links ]
20. Hernández-Verdugo S., R.A. González-Sánchez, F. Porras, S. Parra-Terraza, A. Valdez-Ortiz, A. Pacheco-Olvera y R. G. López-España (2015). Plasticidad fenotípica de poblaciones de chile silvestre (Capsicum annuum var. glabriusculum) en respuesta a disponibilidad de luz. Botanical Sciences (en prensa). [ Links ]
21. Jonas, C.S. y M.A. Geber (1999). Variation among populations of Clarkia unguiculata (Onagraceae) along altitudinal and latitudinal gradients. American Journal of Botany 83: 333-343. [ Links ]
22. Kimura, M. y T. Otha (1971). Theoretical Aspects of Population Genetics. Princeton University Press. Princenton, New Jersey, USA. [ Links ]
23. Law, R. (1979). The cost of reproduction in annual meadow grass. American Naturalist 113: 3-16. [ Links ]
24. Lewu, F.B., D. S. Grierson y A.J. Afolayan (2007). Morphological diversity among accessions of Pelargonium sidoides DC. in the Eastern Cape, South Africa. Genetic Resources and Crop Evolution 54: 1-6. [ Links ]
25. Li, B., J-I. Suzuki y T. Hara (1998) Latitudinal variation in plant size and relative growth rate in Arabidopsis thaliana. Oecologia 115: 293-301. [ Links ]
26. MacNeish, R.S. (1964). Ancient Mesoamerican civilization. Science 143: 531-537. [ Links ]
27. Nattero, J., A.N. Sérsic y A.A. Cocucci (2011). Geographical variation of floral traits in Nicotiana glauca: relationships with biotic and abiotic factors. Acta oecologica 37: 503-11. [ Links ]
28. Nooryazdan, H., H Serieys, R. Baciliéri, J. David y A. Bervillé (2010). Structure of wild annual sunflower (Heliantuhus annus L.) accessions based on agro-morphological traits. Genetic Resources andCrop Evolution 57: 27-39. [ Links ]
29. Olsson, K. y J. Ågren (2002). Latitudinal population diferentiation in phenology, life history and fower morphology in the perennial herb Lythrum salicaria. Journal of Evolutionary Biology 15: 983-996. [ Links ]
30. Oyama, K., S. Hernández-Verdugo, C. Sánchez, A. González-Rodríguez, P. Sánchez-Peña, J.A. Garzón-Tiznado y A. Casas (2006). Genetic structure of wild and domesticated populations of Capsicum annuum (Solanaceae) from northwestern Mexico analized by RAPDs. Genetic Resources and Crop Evolution 53: 553-562. [ Links ]
31. Pacheco-Olvera, A., S. Hernández-Verdugo, V. Rocha-Ramírez, A. González-Rodríguez y K. Oyama K. (2012). Genetic diversity and structure of pepper (Capsicum Annuum L.) from Northwestern Mexico analyzed by microsatellite markers. Crop Science 52: 231-241. [ Links ]
32. Pickersgill, B. (1971). Relationships between weedy and cultivated forms in some species of chili peppers (genus Capsicum). Evolution 25: 683-691. [ Links ]
33. Pickersgill B. (1984). Migration of chili peppers, Capsicum spp. in the Americas. En: Stone D. (ed), pp 105-123. Pre-Columbian plant migration. Harvard University Press, Cambridge, USA. 183 p. [ Links ]
34. Reinartz, J.A. (1984). Life history variation of common mullein (Verbascum thapsus). I. latitudinal differences in population dynamics and timing of reproduction. Journal of Ecology 72: 897-912. [ Links ]
35. Rice, K.J. y R.N. Mack (1991). Ecological genetics of Bromus tectorum. I. A hierarchical analysis of phenotypic variation. Oecologia 88: 77-83. [ Links ]
36. Scheiner, S.M. y D. Berrigan (1998). The genetics of phenotypic plasticity. VIII. Cost of plasticity in Daphnia pulex. Evolution 52: 368-378. [ Links ]
37. Snow, A.A. y D.F. Wigham (1989). Costs of flower and fruit production in Tipularia discolor (Orchidaceae). Ecology 70: 1286-1293. [ Links ]
38. Sohn, J.J y D. Policansky (1977). The costs of reproduction in the mayapple Podophyllum peltatum (Berberidaceae). Ecology 58: 1366-1374. [ Links ]
39. Wright, S. (1951). The genetic structure of populations. Annals of Eugenesic 15: 323-354. [ Links ]

