










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista del Museo de Antropología
Print version ISSN 1852-060XOn-line version ISSN 1852-4826
Rev. Mus. Antropol. vol.7 no.1 Córdoba 2014
ANTROPOLOGÍA BIOLÓGICA
Asimetría direccional del postcráneo en poblaciones prehispánicas del sur de Sudamérica
Postcranial directional asymmetry in southern-South America populations
Federico Lotto* y Marien Béguelin**
* División Antropología, Museo de La Plata, Universidad Nacional de La Plata, La Plata, Argentina. federicolotto@fcnym.unlp.edu.ar ;
** CONICET - Instituto de Investigación en Paleobiología y Geología Universidad Nacional de Río Negro, General Roca, Río Negro, Argentina.
Recibido 28-06-2013
Recibido con correcciones 24-02-2014
Aceptado 12-03-2014
Resumen
El objetivo de este trabajo es estudiar los patrones de asimetría direccional en el tamaño de los huesos largos en poblaciones aborígenes de cazadores-recolectores y agricultores del sur de Sudamérica. Se midieron variables del postcráneo de 272 individuos adultos de ambos sexos correspondientes a cinco muestras de las regiones pampeana, patagónica y Noroeste de Argentina y de la Costa y Valle de Azapa, en el Norte de Chile. Asimismo, se recolectaron datos de 139 individuos adultos de ambos sexos correspondientes a tres muestras del Norte, Sudeste y Sudoeste de Norteamérica. Los datos obtenidos se analizaron en términos de lateralidad y magnitud, dimorfismo sexual y diferencias por modo de subsistencia. Los resultados muestran diferencias significativas en la asimetría direccional en el miembro superior, particularmente en el diámetro de la diáfisis del húmero. A su vez, estos resultados sugieren que el modo de subsistencia, al menos en las categorías consideradas en este trabajo, no es una variable ambiental adecuada para explicar los patrones de asimetría direccional entre las poblaciones estudiadas.
Palabras claves: Lateralidad; Plasticidad; Pautas de actividad; Dimorfismo sexual.
Abstract
The purpose of this study is to analyze directional asymmetry patterns in the size of long bones of hunter-gatherer aboriginal populations of southern South América. Postcranial variables of 272 adult individuals of both sexes were measured, belonging to five samples from Pampa, Patagonia and Norwestern Argentina, and from the northern coast of Chile. Data from 139 adult individuals of both sexes, belonging to three samples from northern, southeastern and southwestern North América were also collected for reference. The entire data set was analyzed in terms of laterality, magnitude, sexual dimorphism and subsistence patterns. Results show significant differences in directional asymmetry of the upper limb, particularly in the diaphyseal diameter of the humerus. In turn, these results suggest that subsistence patterns, at least as they were considered in this study, are not an adequate environmental variable to explain asymmetry patterns in the examined populations.
Keywords: Laterality; Plasticity; Activity patterns; Sexual dimorphism.
La asimetría direccional (AD) se ha definido como el desarrollo diferencial de un rasgo en uno de los planos de simetría respecto al otro (Van Valen 1962; Ruff y Jones 1981). El estudio de los patrones de AD en el esqueleto postcraneal constituye un área de particular interés dentro de las investigaciones bioarqueológicas. La AD ha sido empleada principalmente para realizar inferencias acerca del comportamiento y estilo de vida, en particular del tipo y nivel de movilidad y actividad de las poblaciones prehistóricas (Steele y Mays 1995; Mays 1999; Schoenwolf y Larsen 2009). La AD resulta de la acción diferencial sobre uno de los lados, de los factores sistémicos (e.g. regulación hormonal, disponibilidad de nutrientes, variación genética) y/o locales (e.g. estrés mecánico) que regulan el crecimiento de los huesos largos (Ruff et al. 2006). Aunque ambos tipos de factores podrían contribuir a la AD, la evidencia disponible sugiere el rol de las fuerzas biomecánicas como la principal causa de las diferencias en tamaño y forma entre los miembros derecho e izquierdo (Ruff et al. 2006; Trinkaus et al. 1994; Blackburn et al. 2006). En este sentido, el mayor grado de asimetría observado en la diáfisis del húmero, con respecto a las epífisis y la longitud, es atribuido al efecto del remodelado óseo estimulado por las fuerzas biomecánicas, y reflejaría diferentes demandas funcionales sobre ambos miembros superiores (Jones et al. 1977; Trinkaus et al. 1994).
Numerosos trabajos han señalado que las poblaciones humanas presentan rasgos comunes en la expresión de la AD, y que los mismos se habrían establecido tempranamente en el linaje humano (Trinkaus et al. 1994; Auerbach y Ruff 2006). En particular, el miembro superior se caracteriza por un grado mayor de asimetría que el inferior y se observa un fuerte sesgo hacia el lado derecho en el miembro superior. Esto reflejaría el hecho de que en las poblaciones actuales alrededor del 80% de los individuos utiliza preferentemente el lado derecho, mientras que menos de un 15% emplea el lado izquierdo (Perelle Ehrman 1994; Steele y Mays 1995). Las causas de tales preferencias no son completamente conocidas aunque se han vinculado a la lateralización de las funciones del cerebro humano (Steele 1998). Patrones similares han sido registrados en otros primates no humanos (Falk et al. 1988; Sarringhaus et al. 2005). Por ejemplo, en los chimpancés se observa un mayor uso de la mano derecha y una ausencia de AD en el fémur (Sarringhaus et al. 2005).
Dado que la AD se vincula a los patrones de actividad, sería esperable encontrar diferencias poblacionales vinculadas a distintos modos de vida y sistemas de subsistencia, entre otros. En uno de los estudios más exhaustivos realizados hasta el presente sobre poblaciones humanas pasadas, Auerbach y Ruff (2006) analizaron diversas muestras de diferente cronología y patrones de actividad. Sus resultados indican la existencia de diferencias en la magnitud de asimetría en el miembro superior entre poblaciones del Holoceno temprano y grupos modernos, aunque las frecuencias de individuos con sesgos hacia el lado derecho fueron similares en todas las poblaciones. Esto sugiere que si bien el grado de uso de los miembros puede cambiar de acuerdo con los patrones específicos de comportamiento y sistemas de subsistencia característicos de cada población, la dominancia derecha en el miembro superior habría permanecido constante a través del tiempo.
Sin embargo, estas observaciones se basan principalmente en muestras procedentes de Norte América y Europa, resultando la variación en otras regiones como Sudamérica, en gran medida desconocida (Trinkaus et al. 1994; Churchill et al. 1997; Auerbach et al. 2006). Las poblaciones humanas prehistóricas de América se caracterizan por una gran diversidad morfológica tanto a nivel craneal como postcraneal (Cocilovo y Di Rienzo, 1984; Rothhammer y Silva, 1990) a pesar de tener un origen común reciente (Goebel et al., 2008; Steele y Politis, 2009). Los datos disponibles para el sur de Sudamérica se limitan a un número reducido de muestras de cazadores-recolectores y por lo tanto no es posible evaluar una hipótesis acerca de la relación entre AD y distintos tipos de subsistencia (Auerbach y Ruff 2006; Cocilovo et al, 2006; Varela et al, 2009), por lo que, de manera comparativa se analizan tres poblaciones de cazadores y agricultores de Nrte América. En este trabajo se estudia la asimetría direccional en el tamaño de los huesos largos en poblaciones aborígenes de cazadores-recolectores y agricultores del sur de Sudamérica. El objetivo es abordar de manera preliminar el análisis de los patrones de AD, en términos de lateralidad y magnitud, en los huesos largos de los miembros superior e inferior, y evaluar la existencia de dimorfismo sexual, de diferencias poblacionales y por subsistencia en los patrones de AD. Las hipótesis que implican relaciones filogenéticas entre poblaciones no se analizarán en este trabajo debido a las características de las muestras utilizadas, que no permitirían controlar estos factores en forma adecuada. Sobre la base de lo discutido previamente, se espera que los grupos con un mismo sistema de subsistencia presenten mayores similitudes en la AD de los huesos largos.
Materiales y Métodos
Se midieron variables del postcráneo de 272 individuos adultos de ambos sexos correspondientes a cinco muestras de las regiones pampeana, patagónica y Noroeste de Argentina, y de la costa Norte de Chile (800-500 años AP). Estas muestras se encuentran depositadas en la División Antropología del Museo de La Plata, en el Museo Etnográfico ‘J. B. Ambrosetti’ de la Universidad de Buenos Aires y en el Museo Arqueológico San Miguel de Azapa de la Universidad de Tarapacá, Arica, Chile, y fueron relevadas por uno de los autores (MB).
La muestra de la costa de Chubut (Ch), Patagonia argentina, comprende 26 individuos adultos. Los fechados radiocarbónicos disponibles cubren el rango entre 2600 y 200 años AP para la muestra considerada (Béguelin 2011). Los análisis de isótopos estables del C y N sobre restos humanos así como las evidencias arqueológicas indican que las poblaciones del área se habrían basado en la caza de guanaco complementada con una menor proporción de otros recursos terrestres tales como frutos y raíces y recursos marinos de la zona del estuario y valle inferior del río Chubut (Gomez Otero 2007). La muestra de San Juan (SJ), Noroeste de Argentina, comprende 37 individuos adultos. Si bien no se dispone aún de fechados radiocarbónicos directos, la cronología inferida para la muestra es de 600 a 400 años AP (Gambier et al. 1988). La evidencia arqueológica y de isótopos estables de C y N obtenidos sobre restos humanos señala una dieta centrada en recursos C4, principalmente maíz, y complementada con caza y recolección (Gil et al. 2010). La muestra del SO de Buenos Aires, región pampeana (PP), se compone de 18 individuos adultos correspondientes a momentos históricos (Lehmann Nitsche 1910). La información etnohistórica disponible indica una amplia utilización de recursos basada en la horticultura (Zeballos 1881).
La muestra Chinchorro (Chi), procedente de la Costa del Valle de Azapa (Norte de Chile), comprende 20 individuos adultos del sitio Morro de Arica. Los fechados radiocarbónicos disponibles para el período Chinchorro ubican a la muestra entre 5500 y 4700 años AP (Vera 1981; Varela et al. 2002). Los análisis de isótopos estables de C y N sobre muestras humanas indican el aprovechamiento de recursos costeros, con una fracción marina predominante (Aufderheide 1993). La segunda muestra de Chile proviene del sitio Azapa 71 cerca de San Miguel de Azapa (SM), emplazado a unos 20 kilómetros de la costa sobre el mismo valle, y está conformada por 20 individuos adultos. Los fechados radiocarbónicos para el período correspondiente a esta muestra la ubican entre 960 y 805 años AP (Moraga et al. 2005). La evidencia arqueológica indica una subsistencia basada en la agricultura y la ganadería, y el período comprendido coincide con el Período Expansivo de Tiahuanaco. La historia biológica de la población en los últimos 5000 años en la región presentaría cierto grado de complejidad asociada al flujo génico extrapoblacional (Sutter y Mertz 2004, Varela y Cocilovo 2002).
Asimismo, se recolectaron datos de 139 individuos adultos de ambos sexos correspondientes a tres muestras del Norte, Sudeste y Sudoeste de Norteamérica. El acceso a estas muestras se realizó a través de la base de datos denominada Goldman Data Set (Auerbach y Ruff 2006; http://web.utk.edu/~auerbach/index.htm), de la que se seleccionaron los individuos que presentaban el menor número de datos perdidos para los pares de variables correspondientes al lado izquierdo y derecho de cada estructura anatómica y que no tenían patologías visibles que afectaran las mediciones. La muestra de Tigara (Ti) comprende 48 individuos adultos y corresponde al cementerio de un poblado de la fase Tigara, en la península de Point Hope, Alaska (Estados Unidos). Los fechados radiocarbónicos disponibles ubican esta muestra entre ca. 800 y 300 años AP, y la evidencia arqueológica, así como los estudios de desgaste, indican el aprovechamiento de recursos marinos y costeros (Larsen y Rainey 1948). La muestra procedente del sitio Indian Knoll (IK) Kentucky, Estados Unidos, está constituida por 48 individuos adultos. Los fechados radiocarbónicos para el período considerado sitúan esta muestra entre 5500 y 3700 años AP (Herrmann y Fenton 2000). La muestra corresponde a una población semi sedentaria, basada en recursos fluviales y costeros del Green River superior, y con una mínima inversión en horticultura (Webb y Winters 1974). Finalmente, se incluyó una muestra de Nuevo México, Estados Unidos (Hawikuh, Ha), que comprende 43 individuos adultos y corresponde al período Pueblo IV, entre 500 y 300 años AP. Este período se caracterizó por poblaciones sedentarias y agricultura intensiva en las terrazas del río Zuñi, mientras que la caza no habría representado un aporte significativo a la dieta ((Smith y Woodbury 1966).
A fin de caracterizar los modos de subsistencia de las poblaciones analizadas se tomó en consideración la literatura arqueológica y etnohistórica disponible. Sobre la base de esta información se establecieron tres categorías (cazador-recolector, agricultor y horticultor) que, si bien simplifican la variación real existente entre las poblaciones, constituyen un modelo útil como herramienta analítica y resumen las principales características de cada estrategia de subsistencia con relación al grado de movilidad y composición de la dieta (Auerbach y Raxter 2008). Estos datos son resumidos en la Tabla 1.
Tabla 1. Procedencia geográfica, cronología y número de individuos por sexo de las muestras analizadas.
Table 1. Geographical procedence, chronology and number of male and female individuals in the analyzed samples1: Auerbach y Raxter 2008
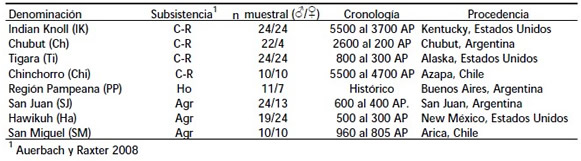
Para las muestras sudamericanas se efectuó la estimación del sexo mediante el análisis de indicadores de la pelvis y el cráneo. En el primer caso se consideraron los criterios propuestos por Phenice (1969) para la región subpúbica (i.e. arco ventral, concavidad subpúbica, rama isquiopúbica) y el ángulo de la escotadura ciática mayor del ilion (Buikstra y Ubelaker 1994). En el cráneo se registraron las características morfológicas de los procesos mastoideos, el margen supraorbital, la glabela, y los arcos superciliares, que en conjunto permiten estimar el sexo con un elevado grado de exactitud (Buikstra y Ubelaker 1994). Para las muestras de norteamérica los datos sobre el sexo fueron obtenidos de la base de datos Goldman Data Set (Auerbach et al. 2006). Las muestras disponibles para este tipo de análisis resultan sumamente limitadas, ya que las condiciones requeridas (presencia bilateral de los elementos óseos de los miembros y buenas condiciones de preservación) no son frecuentes en restos de procedencia arqueológica. La necesidad de contar con muestras representativas de modos de subsistencia suficientemente discretos para los análisis implica, por lo tanto, amplios rangos temporales y geográficos.
Variables y análisis estadístico
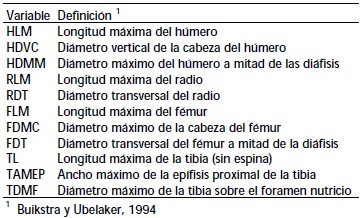
Se relevaron 11 variables morfométricas postcraneanas (Tabla 2) utilizando métodos de la morfometría tradicional, incluyendo longitudes, diámetros y perímetros, siguiendo la metodología de Buikstra y Ubelaker (1994). El criterio de selección de las variables fue que resultaran más representativas de las dimensiones de cada elemento óseo, y que además fueran comparables con variables definidas por otros autores (Auerbach 2007; Béguelin 2011). Las 11 variables fueron utilizadas en la comparación de los patrones de asimetría bilateral en las muestras descriptas previamente.
Tabla 2. Variables postcranealaes analizadas.
Table 2. Postcranial variables analized.
Se obtuvieron índices de asimetría direccional (AD) entre el lado izquierdo (I) y derecho (D) según la siguiente expresión:
AD = (D - I)/ ((D + I) / 2)
De esta manera las mediciones se estandarizaron con respecto al tamaño de las variables originales y se expresan en proporciones. A partir de estos índices se obtuvieron estadísticas descriptivas de cada muestra. Con el fin de determinar la existencia de AD se efectuó también una prueba de t con los índices de cada variable, considerando como hipótesis nula una media cero, de acuerdo a la definición aceptada de AD como distribución normal con media significativamente distinta de cero (Palmer y Strobeck, 1992). Para evaluar la existencia de dimorfismo sexual en los patrones de AD, así como posibles diferencias poblacionales en éstos, se realizó un ANOVA de dos vías utilizando sexo y población como factores fijos. Asimismo, se efectuó un ANOVA modelo mixto empleando las variables subsistencia y sexo como factores fijos y la población como factor aleatorio. Este último análisis permite evaluar la existencia de diferencias en los patrones de AD atribuibles al dimorfismo sexual y las prácticas de subsistencia controlando el efecto de las poblaciones. En ambos análisis las variables dependientes fueron los índices de asimetría direccional obtenidos para cada medida lineal.
Por último se realizó una prueba de Tukey para establecer entre cuáles grupos existían diferencias significativas en todos los ANOVA (resultados no mostrados). Todos los análisis se realizaron en R versión 2.8.0 (R Development Core Team 2011), utilizando el paquete base.
Resultados
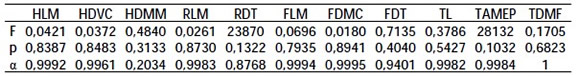
Los resultados del ANOVA de medidas repetidas se pueden observar en la tabla 3. El α obtenido en el CCI es elevado para casi todas las variables, excepto HDMM. El ANOVA, sin embargo, no muestra diferencias significativas entre las medias de las series (F y p están dentro del rango de aceptación en todos los casos), y los valores bajos de CCI pueden ser explicados por la presencia de error al azar. En conjunto, estos resultados indican un bajo grado del error intraobservador en el registro de las variables.
Tabla 3. Análisis del error intraobservador. Resultados del ANOVA de medidas repetidas (F y p) y del CCI (α).
Table 3. Intraobserver error analysis. Repeated measures ANOVA results (F and p) and ICC (α).
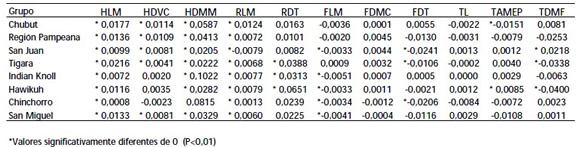
Los resultados obtenidos para el postcráneo en la prueba de t indican que el miembro superior se caracteriza por un mayor grado de AD con un sesgo positivo hacia el lado derecho, mientras en el inferior, el grado de asimetría es menor y ésta es sesgada hacia el lado izquierdo (Tabla 4). Las variables longitud del húmero y radio y diámetro máximo del radio presentaron AD en todos los casos, con una sola excepción, marginal, para el grupo Chinchorro en HDMM, mientras que la cabeza del húmero (HDVC) presentó AD significativa en cinco de los ocho grupos. Por el contrario, las muestras analizadas no presentaron AD significativa en la mayoría de las variables del fémur y la tibia.
Tabla 4. Valor medio de AD por variable y por grupo.
Table 4. Sample mean AD value for each variable by group.
El modelo de ANOVA de dos factores fijos mostró diferencias significativas por población para cinco variables (Tabla 5), y sólo tres variables se diferenciaron por sexo, correspondientes a la longitud del radio y del húmero (RLM y HLM) y el diámetro de la cabeza del húmero (HDVC). La interacción resultó significativa sólo para un diámetro del húmero (HDVC) y uno del fémur (FDT). La prueba post-hoc mostró, para la variable HLM, diferencias significativas entre pares que incluían a las muestras Tigara y Chubut. Para la variable HDMM, los pares que difieren incluyeron a las muestras Chinchorro e Indian Knoll. Para RDT sólo se diferenciaron pares que incluyeran a la muestra Hawikuh. Para TDMF se diferenciaron pares que incluyeron las muestras Hawikuh y Tigara, y para TAMEP sólo se diferenció el par Hawikuh-Chinchorro. Estos resultados se reflejan en la figura 1 que muestra las distribuciones por muestra de las variables con diferencias significativas para el factor población.
Tabla 5. Modelo ANOVA de dos factores, por grupo y sexo.
Table 5. Two-way ANOVA using sex and group as factors.
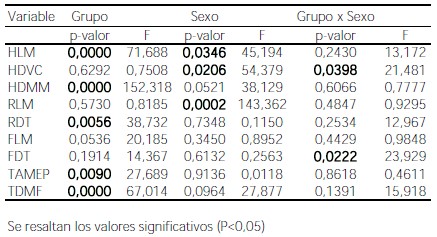
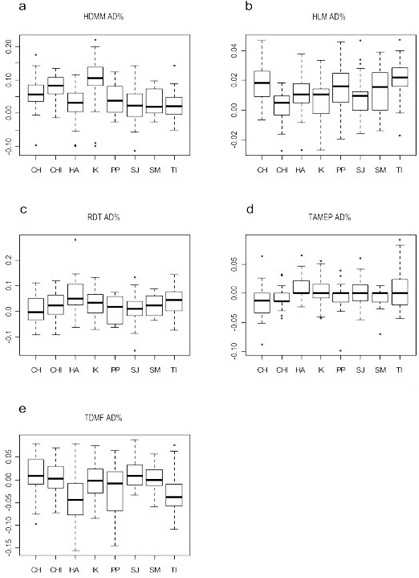
Figura 1. Valores medios y dispersión de AD% de las variables diámetro máximo del húmero a mitad de las diáfisis (a), longitud máxima del húmero (b), diámetro transversal del radio (c), ancho máximo de la epífisis proximal de la tibia (d) y diámetro máximo de la tibia sobre el foramen nutricio (e) para las muestras de Chubut (CH), Chinchorro (CHI), Hawikuh (HA), Indian Knoll (IK), región pampeana (PP), San Juan (SJ), San Miguel (SM), y Tigara (TI).
Figure 1. Average and dispersion AD% values of humerus maximum diameter at midshaft (a), humerus maximum length (b), radius transverse diameter (c), tibia maximum proximal epiphyseal breadth (d) and tibia maximum diameter at the nutrient foramen (e) variables for Chubut (CH), Chinchorro (CHI), Hawikuh (HA), Indian Knoll (IK), Pampa (PP), San Juan (SJ), San Miguel (SM), and Tigara (TI) samples.
El modelo de ANOVA mixto (Tabla 6) no mostró diferencias significativas para el factor subsistencia, aunque se encontraron diferencias para el sexo en las mismas tres variables que fueron significativas en el ANOVA de dos factores fijos. La interacción, a su vez, resultó significativa para los diámetros del miembro inferior, FDT y TDMF. La AD en estas variables, sin embargo, sólo presenta dimorfismo sexual en la muestra de la región pampeana (Figura 2).
Tabla 6. Modelo ANOVA mixto de dos factores fijos (subsistencia y sexo).
Table 6. Mixed ANOVA model with subsistence and sex as fixed factors.
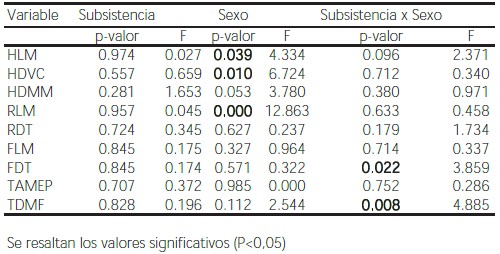
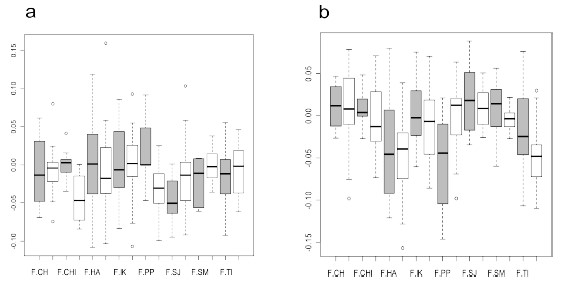
Figura 2. Valores medios y dispersión de AD% de las variables diámetro transversal del fémur a mitad de la diáfisis (a) y diámetro máximo de la tibia sobre el foramen nutricio (b) para las muestras de Chubut (CH), Chinchorro (CHI), Hawikuh (HA), Indian Knoll (IK), región pampeana (PP), San Juan (SJ), San Miguel (SM), y Tigara (TI), separadas en individuos femeninos (gris) y masculinos (blanco).
Figure 2. Average and dispersion of AD% values of femur transverse midshaft diameter (a) and tibia maximum diameter at the nutrient foramen (b) variables for Chubut (CH), Chinchorro (CHI), Hawikuh (HA), Indian Knoll (IK), Pampa (PP), San Juan (SJ), San Miguel (SM), and Tigara (TI) samples, divided into female (grey) and male (white) individuals.
Discusión
Los resultados obtenidos para la AD en los huesos largos de las poblaciones Sudamericanas reflejan, en general, los patrones de asimetría bilateral documentados previamente para diversas poblaciones humanas (Tabla 4). En conjunto, estos resultados indican que los factores ambientales a los que estuvieron sometidos los individuos habrían tenido un efecto similar en ambos miembros inferiores, dada la ausencia de AD significativa en la mayoría de las variables de la tibia y el fémur, mientras difirieron significativamente en el miembro superior. El mayor grado de asimetría observado en la diáfisis del húmero, con respecto a la epífisis proximal y la longitud, seria atribuible al efecto del remodelado óseo estimulado por las fuerzas biomecánicas, y reflejaría diferentes demandas funcionales sobre ambos miembros superiores. El sesgo hacia el lado derecho es consistente con la mayor proporción de individuos que utilizan preferentemente el lado derecho en las poblaciones de humanos modernos (Perelle y Ehrman 1994; Steele y Mays 1995; Auerbach y Ruff 2006).
La presencia de diferencias significativas entre ambos sexos en la longitud del radio (RLM) y del húmero (HLM) y el diámetro de la cabeza del húmero (HDVC), sugiere la existencia de un grado importante de dimorfismo sexual en el miembro superior cuando la muestra es considerada en su conjunto (Tabla 5). En las muestras de Sudamérica los hombres presentaron una mayor AD en la cabeza del húmero, mientras que en las mujeres la longitud del radio fue más asimétrica. Este patrón se observó también en los grupos de Norteamérica con excepción de Tigara, en el cual las mujeres presentaron mayor asimetría en la variable HDVC. El dimorfismo sexual en el grado de AD es frecuentemente empleado para inferir diferencias en el tipo y/o magnitud de las actividades manuales realizadas por hombres y mujeres dentro de una población. Sin embargo, debe tenerse en cuenta que los estudios sobre poblaciones actuales sugieren que las respuestas a las fuerzas biomecánicas en el miembro superior son altamente dimórficas. Cuando son expuestos a una fuerza mecánica similar, los individuos masculinos exhiben un mayor crecimiento óseo compensatorio que las mujeres, resultando en un mayor grado de asimetría bilateral (Jones et al. 1977). Esta respuesta diferencial a un mismo estímulo externo indicaría que una menor asimetría en los individuos femeninos no puede atribuirse necesariamente a actividades diferenciales en relación con la carga soportada.
Asimismo, se detectaron diferencias entre algunas poblaciones en el grado de asimetría del miembro superior, en particular, en la longitud del húmero y los diámetros de las diáfisis del húmero y del fémur (Tabla 5). Estas diferencias son más marcadas en el diámetro de la diáfisis del húmero (Figura 1). La variación poblacional observada podría resultar tanto de factores evolutivos o filogenéticos (i.e. relaciones de parentesco) como ambientales. La importancia de estos factores para explicar el grado de asimetría en rasgos con simetría bilateral entre poblaciones humanas Sudamericanas no ha sido abordada hasta el presente, a diferencia de los estudios efectuados para el componente simétrico de variación (e.g. Cocilovo et al. 2001; González-José et al. 2005; Béguelin 2011; Gonzalez et al. 2011). Aunque no fue probado explícitamente en nuestro trabajo, los análisis sugieren que la estructura de parentesco entre las poblaciones estudiadas no es el factor más importante para explicar la variación observada en la asimentría.
Particularmente, los pares de muestras Sudamericanas con una relación de parentesco relativamente estrecha (i.e. Chinchorro y San Miguel; Pampeanos y Chubut) presentan valores medios de AD marcadamente diferentes. Este resultado sugiere la importancia de alguna variable ambiental para explicar el patrón de variación en la AD entre las poblaciones estudiadas.
El análisis de ANOVA modelo mixto indicó que las diferencias observadas anteriormente utilizando como factor la población (Tabla 5) no estarían vinculadas a las categorías de subsistencia aquí empleadas, al usar como factor la subsistencia (Tabla 6). Sin embargo, si se considera la variable que presentó el mayor grado de asimetría, el diámetro de la diáfisis del húmero, se observa que en tres de las cuatro muestras correspondientes a grupos de cazadores recolectores de Norte y Sudamérica (Ch, IK, Chi) la AD supera el 5% (Figura 1). La muestra de cazadores recolectores de Tigara se aparta de este patrón, ya que se caracteriza por valores similares a los de las muestras de agricultores. Trabajos previos también indican la existencia de diferencias en la AD dentro de cada categoría amplia de subsistencia, las cuales han sido atribuidas al grado de bimanualidad de las actividades efectuadas por los individuos (Auerbach y Raxter 2008, Weiss 2009). Esta hipótesis implica asumir que los cazadores recolectores compartirían actividades de mayores demandas mecánicas unimanuales mientras los agricultores se caracterizarían por actividades bimanuales. Sin embargo, la información disponible para estas poblaciones no permite la discusión de tal hipótesis. La obtención de datos sobre marcadores esqueletales de estrés ocupacional (patológicos y no-patológicos), por ejemplo, constituye una de las líneas de evidencia independiente que podría contribuir a establecer las demandas mecánicas involucradas en las actividades cotidianas realizadas por los individuos (Stirland 1993). Es importante además señalar que en estudios de tipo comparativo no es posible inferir relaciones de causalidad entre las variables, por lo que los resultados del presente trabajo son estrictamente correlacionales.
En síntesis, las poblaciones analizadas muestran diferencias importantes en la asimetría direccional en el miembro superior, particularmente en el diámetro de la diáfisis del húmero. El modo de subsistencia, al menos en las categorías consideradas en este trabajo, no parece ser una variable ambiental adecuada para explicar los patrones de asimetría direccional entre las poblaciones estudiadas. Observaciones similares fueron realizadas por Auerbach y Raxter (2008). Las pautas de actividad particulares de cada grupo podrían ser importantes para comprender las diferencias poblacionales en los valores de asimetría. Futuros estudios con datos independientes acerca de las pautas de actividad, como análisis de los marcadores de estrés musculo-esqueléticos (Hawkey et al. 1995; Kennedy 1989) y las relaciones de parentesco entre las poblaciones estudiadas, son necesarios para explorar esta hipótesis en las poblaciones americanas.
La Plata, 24 de Junio de 2013
Agradecimientos
Agradecemos al personal de la División Antropología del Museo de La Plata, del Museo Etnográfico ‘J. B. Ambrosetti’ de Buenos Aires y del Museo Arqueológico San Miguel de Azapa de Arica, Chile, por permitir y facilitar el acceso a las muestras utilizadas en este trabajo. Agradecemos también a Paula González, Iván Pérez y Valeria Bernal por la revisión crítica del manuscrito.
1. Auerbach, B.M. 2007. “Human skeletal variation in the New World during the Holocene: Effects of climate and subsistence across geography and time”. Baltimore: The Johns Hopkins University. Tesis de posgrado. Disponible en: http://gradworks.umi.com/32/95/3295793.html.
2. Auerbach, B.M., Raxter, M.H. 2008. Patterns of clavicular bilateral asymmetry in relation to the humerus: variation among humans. Journal of human evolution 54(5): p.663–74.
3. Auerbach, B.M., Ruff, C.B. 2006. Limb bone bilateral asymmetry: variability and commonality among modern humans. Journal of human evolution 50(2): p.203–18.
4. Aufderheide, A. 1993. Reconstrucción química de la dieta del hombre de Acha. En I. Muñoz, B. T. Arriaza, A. Aufderheide (eds) Acha 2 y los orígenes del poblamiento humano en Arica, Arica: Universidad de Tarapacá [ Links ]
5. Béguelin, M. 2011. “Variación geográfica en la morfología del esqueleto postcraneal de las poblaciones humanas de pampa y patagonia durante el holoceno tardío: una aproximación morfométrica”. Izeta, A (Editor). 87 pp. B.A.R. International S2253. Oxford
6. Béguelin, M. 2011. Stature estimation in a central Patagonian prehispanic population: development of new models considering specific body proportions. International Journal of Osteoarchaeology 21(2): p.150–158.
7. Blackburn, A., Knüsel, C.J. 2006. Hand dominance and bilateral asymmetry of the epicondylar breadth of the humerus: A test in a living sample. Current anthropology 47(2): p.377–382.
8. Buikstra, J.E., Ubelaker, D. 1994. Standards for data collection from human skeletal remains. En Proceedings of a seminar at the field Museum of Natural History, Arkansas: Arkansas Archaeological Survey Research [ Links ]
9. Churchill, S.E., Formicola, V. 1997. A case of marked bilateral asymmetry in the upper limbs of an upper palaeolithic male from Barma Grande (Liguria), Italy. International Journal of Osteoarchaeology 7(1): p.18–38.
10. Cocilovo JA, Di Rienzo J. 1984. Un modelo biológico para el estudio del poblamiento prehispánico del territorio argentino. Correlación fenético-espacial. Relaciones 16:119-135. [ Links ]
11. Cocilovo, J.A., Varela, H. y Espoueys, O. 2001. El proceso microevolutivo de la poblacion nativa antigua de Arica. Chungará (Arica) 33(1): p.13–20.
12. Falk, D., Pyne, L., Helmkamp, R.C. y DeRousseau, C.J. 1988. Directional asymmetry in the forelimb of Macaca mulatta. American journal of physical anthropology 77(1): p.1–6.
13. Gambier, M., Michieli, C.T. y Iantanos, N. 1988. La fase cultural Punta del Barro. San Juan: Instituto de Investigaciones Arqueológicas y Museo, Facultad de Filosofía, Humanidades y Artes, Universidad Nacional de San Juan. [ Links ]
14. Gil, A.F., Neme, G.A. y Tykof, R.H. 2010. Isótopos estables y consumo de maíz en el centro occidente argentino: tendencias temporales y espaciales. Chungará (Arica) 42(2): p.497–513.
15. Goebel T, Waters MR, O’Rourke DH. 2008. The late Pleistocene dispersal of modern humans in the Americas. Science 319:1497-1502.
16. Gomez Otero, J.E. 2007. Dieta, uso del espacio y evolución en poblaciones cazadoras-recolectoras de la costa centro – septentrional de Patagonia durante el Holoceno medio y Tardío.
17. Gonzalez, P.N., Perez, S.I. y Bernal, V. 2011. Ontogenetic Allometry and Cranial Shape Diversification Among Human Populations From South America. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 294(11): p.1864–1874.
18. González-José, R. Ramírez-Rozzi, F., Sardi, M., Martínez-Abadías, N., Hernández, M. y Pucciarelli, H. 2005. Functional-cranial approach to the influence of economic strategy on skull morphology. American journal of physical anthropology 128(4): p.757–771.
19. Hawkey, D.E., Merbs, C.F. 1995. Activity-induced musculoskeletal stress markers (MSM) and subsistence strategy changes among ancient Hudson Bay Eskimos. International Journal of Osteoarchaeology 5(4): 324–338.
20. Herrmann, N.P., Fenton, J. 2000. A report on new radiocarbon dates and recent investigations at three Green River Archaic sites. En 7th Kentucky Heritage Council Archaeological Conference, Bowling Green, KY [ Links ]
21. Jones, H.H., Priest, J.D., Hayes, W.C., Tichenor, C.C. y Nagel, D.A. 1977. Humeral hypertrophy in response to exercise. The Journal of bone and joint surgery. American volume 59(2): p.204–208.
22. Kennedy, K.A.R. 1989. Skeletal Markers of Occupational Stress. Reconstruction of Life from the Skeleton, 129–160. Nueva York: Alan R. Liss
23. Larsen, H., Rainey, F. 1948. Ipiutak and the Arctic whale hunting culture. New York: Published by order of the Trustees of the American Museum of Natural History. Anthropological papers of the American Museum of Natural History; v. 42. Disponible en: http://www.getcited.org/pub/101176360. [ Links ]
24. Lehmann Nitsche, R. 1910. Catálogo de la sección Antropología del Museo de La Plata. Buenos Aires: Coni Hnos. [ Links ]
25. Mays, S. 1999. A biomechanical study of activity patterns in a medieval human skeletal assemblage. International Journal of Osteoarchaeology 9(1): p.68–73.
26. Moraga, M., Santoro, C.M., Standen, V.G., Carvallo, P. y Rothhammer, F. 2005. Microevolution in prehistoric Andean populations: chronologic mtDNA variation in the desert valleys of northern Chile. American journal of physical anthropology 127(2): p.170–81.
27. Perelle, I.B., Ehrman, L. 1994. An international study of human handedness: the data. Behavior genetics 24(3): p.217–27.
28. Phenice, T.W. 1969. A newly developed visual method of sexing the os pubis. American Journal of Physical Anthropology 30(2): p.297–301.
29. R Development Core Team. 2011. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. [ Links ]
30. Rothhammer F, Silva C. 1990. Craniometrical variation among South American prehistoric populations: climatic, altitudinal, chronological, and geographic contributions. American journal of physical anthropRuff, [ Links ] C.B., Holt, B. y Trinkaus, E. 2006. Who’s afraid of the big bad Wolff?: ‘Wolff’s law’ and bone functional adaptation. American journal of physical anthropology 129(4): p.484–498.
31. Ruff, C.B., Jones, H.H. 1981. Bilateral asymmetry in cortical bone of the humerus and tibia-sex and age factors. Human biology 53(1): p.69–86.
32. Sarringhaus, L.A., Stock, J.T., Marchant, L.F. y McGrew, W.C. 2005. Bilateral asymmetry in the limb bones of the chimpanzee (Pan troglodytes). American journal of physical anthropology 128(4): p.840–5.
33. Schoenwolf, G.C., Larsen, W.J. 2009. Larsen’s human embryology. Philadelphia: Churchill Livingstone. Disponible en: http://books.google.com.ar/books?id=56xqAAAAMAAJ.
34. Smith, W., Woodbury, R.B. 1966. The Excavation of Hawikuh by Frederick Webb Hodge: report of the Hendricks-Hodge Expedition, 1917-1923. Contributions from the Museum of the American Indian, Heye Foundation 20: p.336. [ Links ]
35. Steele, J. 1998. Cerebral asymmetry, cognitive laterality, and human evolution. Current Psychology of Cognition 17: p.1202–1214.
36. Steele, J., Mays, S. 1995. Handedness and directional asymmetry in the long bones of the human upper limb. International Journal of Osteoarchaeology 5(1): p.39–49.
37. Steele J, Politis G. 2009. AMS 14C dating of early human occupation of southern South America. Journal of archaeological science 36:419-429. [ Links ]
38. Stirland, A.J. 1993. Asymmetry and activity-related change in the male humerus. International Journal of Osteoarchaeology 3(2): p.105–113.
39. Trinkaus, E., Churchill, S.E. y Ruff, C.B. 1994. Postcranial robusticity in Homo. II: Humeral bilateral asymmetry and bone plasticity. American Journal of Physical Anthropology 93(1): 1–34.
40. Van Valen, L. 1962. A study of fluctuating asymmetry. Evolution 16(2): 125–142.
41. Varela, H.H., Cocilovo, J.A. 2002. Genetic drift and gene flow in a prehistoric population of the Azapa valley and coast, Chile. American Journal of Physical Anthropology 118(3): 259–67.
42. Vera, J. 1981. Momias Chinchorro de preparación complicada del Museo de Historia Natural de Valparaíso: 3290 y 3060 A.C. Anales del Museo de Historia Natural de Valparaíso 14: 5–17.
43. Webb, W.S., Winters, H.D. 1974. Indian knoll. Knoxville: University of Tennessee press. [ Links ]
44. Weiss, E. 2009. Sex differences in humeral bilateral asymmetry in two hunter-gatherer populations: California Amerinds and British Columbian Amerinds. American Journal of Physical Anthropology 140(1): 19–24.
45. Zeballos, E. 1881. Descripcion amena de la Republica Argentina. Viaje al país de los Araucanos. Buenos Aires: Peuser. [ Links ]

