










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista del Museo de Antropología
versão impressa ISSN 1852-060Xversão On-line ISSN 1852-4826
Rev. Mus. Antropol. vol.12 no.1 Córdoba abr. 2019
ARQUEOLOGÍA
Elementos para explorar el uso de gramíneas silvestres de ambientes áridos de los Andes Centro Sur: primeras aproximaciones desde los conjuntos fitolíticos de inflorescencias e infrutescencias
Elements to explore the use of wild grasses from arid environments of the South Central Andes: first approaching from phytolith assemblages in inflorescences and infrutescences
Maria Gabriela Musaubach*, María del Pilar Babot**
*Maria Gabriela Musaubach, Facultad de Humanidades y Ciencias Sociales, Universidad Nacional de Jujuy (FHyCS, UNJu) Otero N° 262, (4600) San Salvador de Jujuy, Argentina. gabrielamusa@gmail.com
**María del Pilar Babot, Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán (IAM, FCN e IML, UNT) - Instituto Superior de Estudios Sociales, Consejo Nacional de Investigaciones Científicas y Técnicas/UNT (ISES, CONICET/UNT). San Martín 1545, (4000) San Miguel de Tucumán, Argentina. pilarbabot@yahoo.com
Recibido 17-07-2018.
Recibido con correcciones 03-01-2019.
Aceptado 06-02-2019
Resumen
En este trabajo se analiza el estado del arte del conocimiento sobre los usos pasados y presentes de las gramíneas en el desierto de altura puneño. A partir de ello se establece la pertinencia de explorar la utilización de estas plantas mediante el estudio de microfósiles. Se presentan los primeros resultados obtenidos en relación a la caracterización fitolítica de inflorescencias e infrutescencias de especies silvestres de Poaceae de esta área y sobre esta base, se introduce una discusión sobre aspectos metodológicos y conceptuales que afectan la interpretación del registro de microindicadores modernos y fósiles, tales como continuo morfológico y conjuntos fitolíticos redundantes.
Palabras clave: arqueobotánica; desierto de altura puneño; Poaceae; continuo morfológico; conjunto de fitolitos
Abstract
The state of the art of knowledge about the past and present uses of grasses in the Punean high desert, is analyzed. Then, the relevance of exploring the use of these plants through the study of microfossils is established. The first results obtained on the phytolithic characterization of inflorescences and infrutescences of wild Poaceae species in this area are presented. On this basis, a discussion is introduced on some methodological and conceptual aspects that affect the interpretation of the modern and fossil micro indicators' record, such as morphological continuum and redundant phytolith assemblages.
Keywords: archaeobotany; Punean high desert; Poaceae; morphological continuum; phytolith assemblage
Este trabajo forma parte de una investigación en desarrollo en relación a las Poáceas nativas silvestres de ambientes áridos y semi-áridos del actual territorio argentino. Se realiza desde una perspectiva arqueobotánica orientada a identificar las interacciones entre las personas y estas plantas en el pasado prehispánico del área sobre la base del registro microfósil arqueológico.
Aquí nos enfocamos en la Puna, un desierto de altura sobre los 3000 m.s.n.m. que caracteriza a los Andes Centro Sur del Noroeste argentino y chileno. Fitogeográficamente, en la Puna predomina la estepa arbustiva xerófila baja y muy abierta, con escasa cobertura del suelo. También se desarrollan la estepa herbácea, psammófila halófila y las vegas. Las gramíneas crecen en las vegas a la vera de ojos de agua y cursos permanentes y típicamente conforman la unidad vegetacional del pajonal (Burkart et al., 1983; Cabrera, 1976). En la Puna Salada, sector al que corresponde el caso que analizamos, el pajonal conforma un pastizal en el que abundan especies de Festuca, Stipa y Deyeuxia (Poaceae), otras herbáceas como Trichocline (Asteraceae) y arbustivas de los géneros Adesmia, Parastrephia, Baccharis (Asteraceae) y Fabiana (Solanaceae). Esta unidad de vegetación se halla sobre ondulaciones a gran altura (entre los 3800 y 4700 m.s.n.m.), conformando un área sometida a fuertes vientos y temporales. Las vegas pueden situarse a menor altura; en ellas se encuentran los géneros Festuca y Distichlis (Haber, 1992).
Los estudios paleoambientales disponibles para cuencas de la Puna Salada indican una alternancia de fases que implicaron variaciones en la disponibilidad de agua a lo largo del Holoceno, con momentos de mayor humedad entre comienzos del Holoceno y ca. 8500 AP y entre ca. 4500-1600 AP, alternados con momentos de menor humedad entre los ca. 8500 y ca. 4500 AP y desde ca. 1600 AP hasta la conformación del clima actual. A su vez, estas variaciones habrían afectado de manera diferencial a distintas cuencas (Grana, Tchilinguirian, Hocsman, Escola y Maidana, 2016; Tchilinguirian y Olivera, 2014). Tales circunstancias habrían impactado en la distribución espacial y altitudinal de las comunidades vegetales respecto de su actual emplazamiento y, posiblemente, también producirían variaciones en la cobertura vegetal (densidad), pero sin alterar mayormente la riqueza florística del área. Por esto, los estudios de gramíneas modernas de la zona tienen directa aplicación para el conocimiento de la vida en el pasado de este lugar.
Dentro de la Puna Salada, Antofagasta de la Sierra ha proporcionado información contundente sobre el uso prehispánico de las gramíneas silvestres en distintos contextos y a lo largo de toda la secuencia ocupacional arqueológica. Esto tiene un referente moderno en los usos tradicionales de estas plantas, que aún se verifican localmente y en distintos sectores de los Andes Centro-Sur (Cuello, 2006; Lund, 2016; Olivera, 2006; Pérez, 2006; Villagrán y Castro, 2003, entre otros).
El principal registro arqueológico del uso de gramíneas se ha sustentado en la identificación de macrorrestos vegetales, preservados en condiciones excepcionales en esta área, que han sido caracterizados en numerosas publicaciones realizadas por C. Pérez de Micou y E. Ancíbor (Pérez de Micou y Ancíbor, 1994) y por F. Rodríguez y colaboradores (Mondini, Martínez, Pintar y Reigadas, 2013; Rodríguez 2013a, 2013b; Rodríguez y Martínez, 2001; Rodríguez y Rúgolo de Agrasar, 1999; Rodríguez, Rúgolo de Agrasar y Aschero, 2003, 2006). Estos trabajos inicialmente demostraron la utilización de estas plantas en el pasado y pudieron vincularlas principalmente al acondicionamiento de los pisos de ocupación, la construcción de viviendas y la manufactura de instrumentos.
Las gramíneas silvestres también se encuentran representadas en el registro de microfósiles (Babot, 2004, 2011). Sin embargo, en el estado actual del conocimiento no ha sido posible establecer claramente si este registro corresponde a una señal del contexto natural y cultural que incluye a determinadas gramíneas y/o al resultado del procesamiento alimenticio o con fines artesanales. Esto se debe a dos factores interdependientes: por un lado, este grupo de plantas aún no está representado en las colecciones de referencia regionales de micropartículas o microindicadores vegetales, y por otro lado, recientemente se ha retomado la pregunta del papel de las plantas silvestres útiles en el consumo, entre ellas, las gramíneas (Hocsman y Babot, 2018; Lund, 2016), como un aspecto relevante para la arqueología del área, luego de que fuera planteada inicialmente hace unos años (Babot, 2004). Así, la revisión de conjuntos graminoides compuestos por células cortas, largas, apéndices dérmicos silicificados y granos de almidón, entre otros, procedentes de residuos de uso en el instrumental arqueológico se encuentra en proceso (Hocsman y Babot, 2018), motivado por un incremento en el trabajo etnobotánico realizado con las comunidades locales (Babot, Lund y Olmos, 2016; Lund, 2016; Lund y Babot, 2014) y por la conformación de las primeras bases de referencia de microindicadores de gramíneas en inflorescencias e infrutescencias en otros ambientes áridos y semi-áridos del Cono Sur desde perspectivas arqueológicas (Babot, Musaubach y Plos, 2017; Musaubach, 2014; Musaubach Babot y Plos, 2013a; Musaubach, Plos y Babot, 2013b).
La confección de colecciones de referencia es parte fundamental del estudio arqueobotánico para caracterizar las partículas biosilíceas que son producidas por las plantas, focalizando en morfotipos que remiten a diferentes partes útiles (Korstanje y Babot, 2007). En el caso de las preguntas referidas al uso de las gramíneas, esto es central ya que, en términos generales, se esperan usos disímiles para las cariopses (alimento), inflorescencias (ornamento) y las hojas, cañas y cañas floríferas (manufacturas e instalaciones), según los registros arqueológicos y etnográficos locales -ver infra-. De este modo, es deseable poder establecer diferentes conjuntos de morfotipos de referencia para las distintas partes útiles.
Además, la frecuencia de morfotipos silíceos de células cortas tiene un valor en la asignación taxonómica de las gramíneas al nivel de subfamilias (Twiss, 1987; Twiss, Suess, Smith, 1969; Mulholand y Rapp, 1989; Piperno, 2008). No obstante, al establecer los conjuntos fitolíticos en el material de referencia se verifica un continuo morfológico (Babot et al., 2017) que se aplica a morfotipos de morfologías próximas y sus formas intermedias. Esta situación tiene relevancia al momento de realizar las interpretaciones del registro arqueológico porque condiciona los recuentos de los microrestos y, por lo tanto, las interpretaciones que se basan en ellos. El conteo de tipos de células cortas se vuelve crucial en muestras con un bajo porcentaje de fitolitos, como es el caso de las muestras arqueológicas de residuos de uso y tártaro dental.
El propósito de este trabajo es analizar el estado del arte sobre los usos pasados y presentes de las gramíneas puneñas, realizar una presentación de los primeros resultados obtenidos en relación a la caracterización fitolítica de inflorescencias e infrutescencias, como partes de las gramíneas silvestres de esta área potencialmente útiles en la alimentación y sobre esta base, introducir una discusión sobre los aspectos metodológicos e interpretativos señalados en los párrafos precedentes.
Los usos pasados y presentes de las gramíneas silvestres en la Puna
Las gramíneas silvestres en los registros etnobotánicos de la Puna
Existen numerosos registros etnobotánicos1 acerca de la utilidad de las gramíneas silvestres nativas de la Puna argentina y chilena. La raíz de Panicum urvilleanum Kunth (jaboncillo) constituye un sustituto del jabón en el lavado de la ropa (Pérez, 2006). El empleo con fines medicinales se registra en el caso de la raíz de Cortaderia speciosa (Nees & Meyen) Stapf (raíz de cortadera), Distichlis spicata (L.) Greene y D. humilis Phil. (gramillas), y de la raíz y mata de Festuca orthophylla Pilg (iro) (Olivera, 2006; Pérez, 2006). Las hojas y raíces de Cortaderia atacamensis (Phil) Pilger (cortadera, cola de zorro), Dielsiochloa floribunda (Pilger) Pilger (peludilla) y Distichlis sp. (brama) también se refieren como medicinales en el norte de Chile (Villagrán y Castro, 2003). Allí, la caña de C. atacamensis, Stipa chrysophylla E. Desv. (paja blanca) y Deyeuxia eminens J. Presl (waylla) se empleaban en artesanías como canastos, sombreros, recipientes y aparejos para los burros.
Otras referencias de Villagrán y Castro (2003) procedentes del área circumpuneña chilena indican usos vinculados a la alimentación humana. Con los tallos más gruesos de D. eminens se tuesta la quinua ("hacer pisi"). Nassella rupestris (Phil.) Torres (paja de chojlla, ñawara), Anatherostipa venusta (Phil.) Peñail (paja amarilla, k'isi) y Stipa leptostachya Griseb. (paja de chuño) sirven como saborizantes para la cocción de un pan de harina de quinua denominado mukuna. Como su nombre vernacular lo indica, esta última también se utiliza como soporte para colocar las papas en el proceso de deshidratación por congelamiento de los mismos. Los tallos de Pennisetum chilense (E. Desv.) B. D. (esporal), en cambio, se emplean en el tostado de maíz. En la cuenca de Antofagasta de la Sierra, la raíz de F. orthophylla se usa como alimento ocasional en el campo, mientras se pastorea (Lund, 2016; Pérez, 2006).2
En el área circumpuneña de Argentina y Chile, especies de los géneros Festuca, Bromus, Deyeuxia, Distichlis, Hordeun, Munroa, Bouteloua, Eragrostis, Puccinenellia, Stipa, Pappostipa, Nasella, Jarava, Cynodon, Aristida, Dielsiochloa y Enneapogon son ampliamente reconocidos como forraje para la fauna nativa silvestre y domesticada (Camelidae) (Olivera, 2006; Pérez, 2006; Villagrán y Castro, 2003). De S. chrysophylla y Festuca chrysophila Phil. (iro, paja vizcachera) se dice que los llamos y alpacos en el primer caso y los burros, llamas, guanacos, vicuñas y vizcachas, en el segundo caso, consumen preferentemente su espiga denominada parwalla (Villagrán y Castro, 2003).
Festuca chrysophila, F. orthophylla, F. deserticola (Phill) (guajilla, guaia), S. chrysophylla, Deyeuxia eminens, D. nardifolia (Griseb.) Phil. (wailaka) y C. atacamensis se utilizan en techumbres (Aldunate, Armesto, Castro y Villagrán, 1981; Olivera, 2006; Villagrán y Castro, 2003). En el caso de F. chrysophylla esto ocurría antes de la floración, en cuya ocasión se chancaba o golpeaba la parte de la raíz para dejarla limpia. Los tallos de C. atacamensis se disponían debajo de la paja brava; esta planta y D. eminens se usaban como primera capa del techo; actualmente la primera se usa para hacer ramadas y la segunda para techar la cocina. F. hypsophila Phil. (paja waya) desprovista de su raíz, machacada y mezclada con barro, servía a modo de aislante de techos. S. chrysophylla, A. venusta y N. rupestris se aplicaban molidas y mezcladas con barro en los "cielos" o "torta" de techos. Con A. venusta también se hacen camas y con P. chilense, rellenos para ensillar a los animales. D. eminens, A. venusta, N. rupestris y S. leptostachya se señalan como especies útiles en la confección de adobes; y D. eminens mezclada con barro, para cerrar los canales de riego (Villagrán y Castro, 2003).
El nombre vernacular kafle o sáflu que se aplica a Eragrostis virescens J. Presl., Polypogon interruptus Kunth. y Hordeum murinum L. se registra en cantos regionales de Atacama que se entonan durante la limpieza de canales de riego. Jarava frigida (Phil.) F. Rojas (paja fina) sirve para espantar a los espíritus malos. Las espigas de C. atacamensis, llamadas kuchucho, a veces teñidas de colores, se utilizan con fines ornamentales en los hogares y cementerios. También tendría usos referidos a las almas Deyeuxia cabrerae (Parodi) Parodi (paja sikuya) (Villagrán y Castro, 2003).
Las gramíneas silvestres en el registro arqueológico de la Puna meridional
Como señaláramos, debido a las condiciones excepcionales de preservación orgánica del desierto de altura, han sido recuperados diferentes ecofactos y artefactos manufacturados sobre gramíneas silvestres locales y no locales3 que corresponden a las sociedades prehispánicas de la Puna argentina. Estos constituyen numerosos antecedentes arqueológicos que se refieren a los usos no alimenticios de las hojas, cañas, cañas floríferas y raíces de estas plantas en contextos residenciales, funerarios y otros rituales.
A lo largo de todo el Holoceno, las gramíneas silvestres de los géneros Deyeuxia, Festuca, Pappostipa, Stipa y Puccinellia se emplearon en la cuenca de Antofagasta de la Sierra (Puna Salada) en el acondicionamiento de los espacios residenciales de varios sitios, siendo dispuestas en "camadas de paja" para mullir los pisos de ocupación (López Campeny, Romano, Rodríguez, Martel y Corbalán, 2014; Pintar y Rodríguez, 2015; Rodríguez, 2013a; entre otros). Las camadas estaban integradas por hojas, cañas floríferas y portaban, ocasionalmente, órganos reproductivos (panojas y espiguillas). De manera similar, también se acondicionaron los espacios funerarios como en los sitios Peñas de las Trampas 1.1 (8440±40 y 8210±50 AP; Martínez, 2012), Quebrada Seca 3 (ca. 4510±100 AP, 4410±60 AP, Gonzalez Baroni, Hocsman y Aschero, 2017), Punta de la Peña 11A y 11B (Aschero, 2010) y Punta de la Peña 9.III (López Campeny, 2006-2007), mediante el uso de haces de D. eminens var. eminens y D. rigescens (Rodríguez, 2013b; Rodríguez y Rúgolo de Agrasar, 1999; Rodríguez et al., 2003), al igual que los espacios de ofrenda, como en el sitio Punta de la Peña 9.III (530±50 AP, Aschero, 2010). Otras modalidades de uso de gramíneas corresponden a techumbres de las viviendas del primer milenio D.C., según el registro del recinto E3 de Punta de la Peña 9.I (1430±60 AP, Babot, 2016).
Cordeles, sogas y nudos manufacturados con diferentes especies del género Deyeuxia, han sido registrados, por ejemplo, en los sitios Quebrada Seca 3 y Punta de la Peña 4 (ca. 2500-9000 AP) (Rodríguez y Rúgolo de Agrasar, 1999; Rodríguez et al., 2003). En el primero de ellos también se recuperó cestería en C. speciosa (2480±60 AP y 4510±100 / 4410±60 AP) (Aschero, 2010; Pérez de Micou y Ancíbor, 1994; Rodríguez, 1999) al igual que en Real Grande 1 y 9 (ambos sitios sin datar, Olivera, 1988; Podestá, 1991), habiéndose utilizado la lámina de la hoja, el nervio medio, la raíz y el culm o pedúnculo floral en diferentes sectores de los contenedores (Pérez de Micou y Ancíbor, 1994). Dos cestas más del mismo material -Cortaderia sp.- proceden del contexto funerario del sitio Punta de la Peña 11A (3680±50 AP, Aschero, 2010). Los fragmentos de una red de lazos anudados que envolvían una urna funeraria en Punta de la Peña 13 indicaron la presencia de un complejo ensamble de fibras de la caña florífera de Deyeuxia aff. deserticola, bromeliáceas y arecáceas, junto a un tallo de Schoenoplectus asper Presl. rodeado por cañas floríferas de Festuca sp. (1280±60 AP, López Campeny et al., 2014). De importancia, en el sitio Peñas Chicas 1.5 se recuperó un fragmento de un recipiente de barro con alma de fibra vegetal de Cortaderia speciosa sin cocción (ca. 3800 AP) asociado a restos de esa planta (Aschero y Hocsman, 2011). Este constituye el antecedente más temprano de alfarería en Argentina y uno de los más tempranos en los Andes Centro Sur. Otros usos de las gramíneas silvestres del género Deyeuxia incluyen la confección de paquetes de cuero rellenos con vegetales, dispuestos como parte de un acompañamiento mortuorio en Punta de la Peña 9.III (1480±40 AP, López Campeny, 2006-2007).
El único registro macrobotánico que ha sido vinculado al uso culinario de Poáceas silvestres puneñas corresponde al empleo de Deyeuxia eminens var. eminens y Jarava vaginata (Rodríguez, 2013b). Gran cantidad de espigas y algunas hojas se encontraron en asociación con granos de quínoa (Chenopodium quinua Willd.), probablemente en un contexto de tostado del grano andino en el sitio Punta de la Peña 4 (760±40 AP) en el que las gramíneas se utilizarían como un medio instrumental para tal fin (Rodríguez et al., 2006).
En lo que respecta a la información procedente del estudio de microfósiles en residuos de uso, Babot (2009, 2011, 2016) ha reportado la presencia de fitolitos graminoides en artefactos de molienda de varios sitios arqueológicos de las localidades Quebrada Seca, Punta de la Peña y Peñas Chicas, en la cuenca de Antofagasta de la Sierra entre ca.7500-400 AP. Debido a la presencia de Poáceas en el entorno natural y de camas y artefactos manufacturados con hojas y tallos de esas plantas en los niveles arqueológicos de algunos de los sitios, no ha sido posible especificar si esos fitolitos procedían del procesamiento de los granos o fibras o bien, de la depositación natural contemporánea o postdepositacional (contaminación cruzada). Morfotipos fitolíticos graminoides también han sido registrados en grandes cuchillos líticos de los sitios Punta de la Peña 9, Punta de la Peña 12 y Casa Chávez Montículos que fueron empleados en la cosecha de la panoja de quinua (Chenopodium quinoa) entre otras labores agrícolas ca. 1600-1200 AP (Escola, Hocsman y Babot, 2013), y en puntas de proyectil reutilizadas como cuchillos de Quebrada Seca 3 que se usaron para el corte de carne y tubérculos asados ca. 5500-4500 años AP (Babot, Cattáneo y Hocsman, 2013). Otros hallazgos se refieren a la presencia de almidón y silicofitolitos de Poáceas en tártaro dental humano de individuos procedentes de Punta de la Peña 9 (ca. 1388±45 AP / 1306±44 AP), interpretados como consumo de cariopses debido al registro de almidón (Gonzalez Baroni y Babot, 2013).
En lo que respecta a la Puna Seca, un manojo de gramíneas atado con un cordel, un atado hecho con una gramínea y un ramillete de plumas y pelos atado con gramíneas en la base del raquis proceden del sitio Inca Cueva cueva 7, ca. 4100-3900 AP (Pérez de Micou, López Campeny y Lessa Costa, 2016).
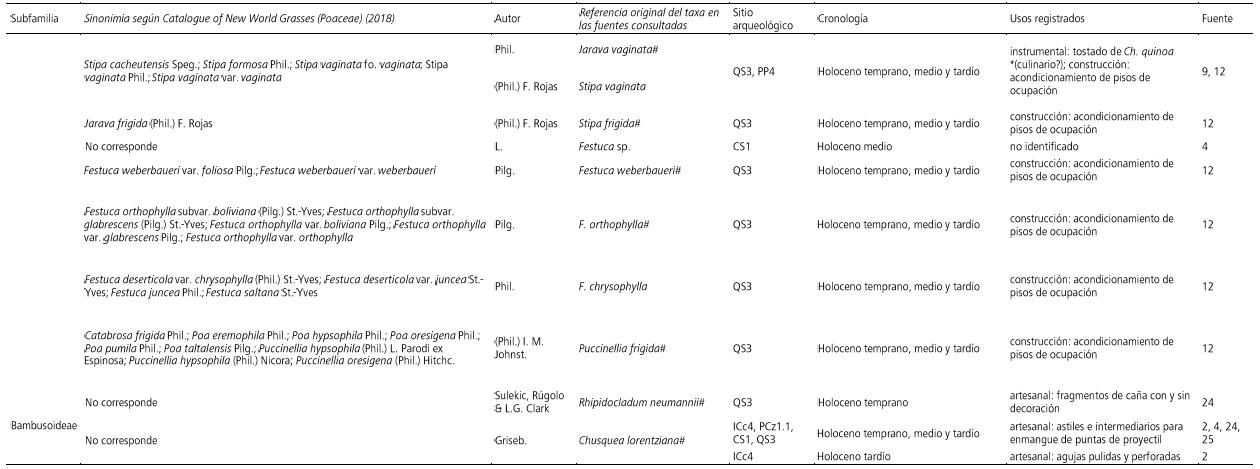
Por su parte, las gramíneas con cañas, no locales, del género Chusquea lorentziana Griseb., se emplearon en la manufactura de astiles para el enmangue de puntas de proyectil en sistemas de caza tanto en la Puna Seca (sitio Inca Cueva, cueva 4, provincia de Jujuy) como en la Puna Salada (sitios Peñas de la Cruz 1, Cueva Salamanca 1 y Quebrada Seca 3, provincia de Catamarca), entre ca. 10600-4080 AP (Aschero, 2010; Mondini et al., 2013; Pintar y Rodríguez, 2015; Rodríguez y Martínez, 2001). Con esta caña también se manufacturaron agujas pulidas y perforadas en Inca Cueva, cueva 4 (4080±80 AP). Además, fragmentos de caña de Rhipidocladum neumannii Sulekic, Rúgolo & L.G. Clark con y sin decoración se hallaron en Quebrada Seca 3 (ca. 8600 AP) (Rodríguez y Martínez, 2001). Todos estos casos corresponden a especies de ambientes húmedos del Noroeste argentino, distantes de los sitios arqueológicos en los que fueron identificados.
Aproximación metodológica y conceptual para el estudio de microindicadores
A partir de los registros de gramíneas que se distribuyen en Antofagasta de la Sierra se realizó una primera selección de especies que forman parte de la cobertura natural, contando, algunas de ellas, con usos documentados. Los registros de especies fueron realizados por distintos autores en diferentes épocas utilizando el nombre científico vigente en ese momento. Al trabajar con material de herbario en esta investigación fue necesario establecer las sinonimias que permiten relacionar los taxones mencionados en la bibliografía con los que se encuentran depositados en los reservorios. Conocer las sinonimias de las plantas relevadas para una región nos permite conocer el universo real compuesto por los taxones englobados en la problemática bajo estudio y establecer comparaciones entre los resultados de las investigaciones precedentes.
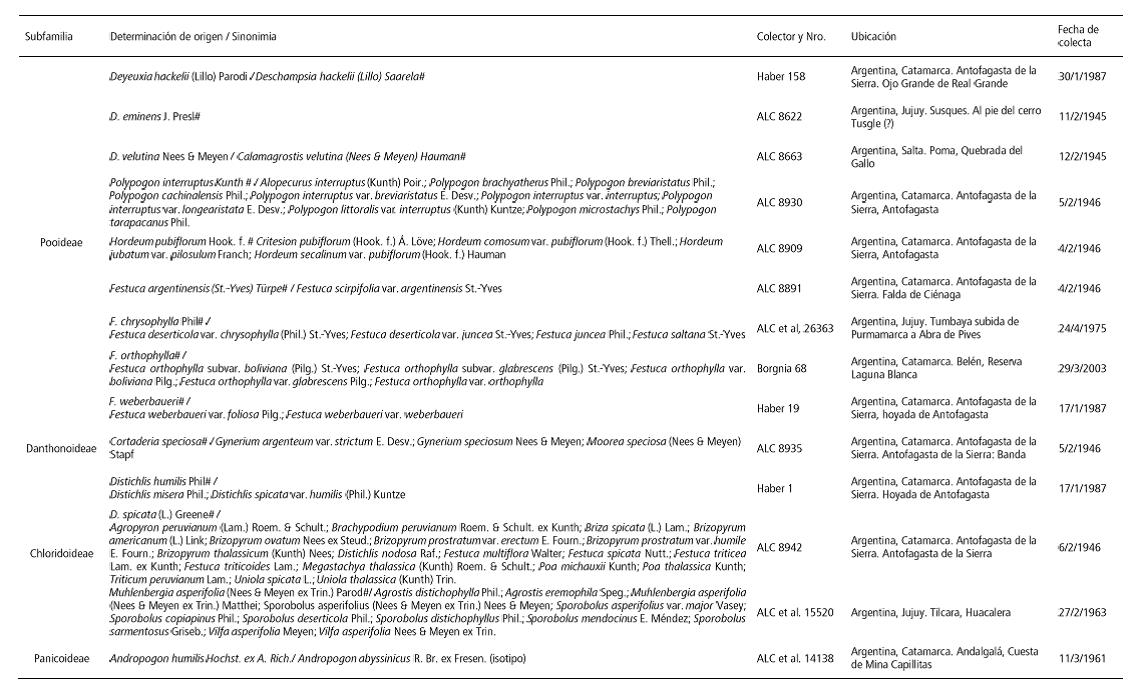
Los especímenes muestreados (tabla 1) se encuentran catalogados en el Herbario del Museo de La Plata y fueron colectados en el área de estudio o áreas ambientalmente similares. Esto último fue contemplado en la selección de los ejemplares a los fines de controlar la variable ambiental en la producción fitolítica. Para este trabajo se seleccionaron muestras de las inflorescencias/ infrutescencias (incluyendo ejes y cariopses o flores), dependiendo del estado vegetativo de cada ejemplar. Previo al muestreo y considerando la antigüedad de la fecha de colecta de los especímenes, en algunos casos, de más de 70 años, se procedió a su rehidratación en agua destilada por el término de 24 horas. Se recurrió a la técnica de calcinado en seco (dry ashing) de las dos secciones disectadas por separado, disponiendo en mufla a 600ºC durante 5 horas. Para procurar la recuperación de fitolitos articulados, se evitaron posteriores tratamientos del material calcinado, según lineamientos de bajo impacto en colecciones de referencia aplicados en casos de estudio anteriores. Estos evitan la desagregación durante el procesamiento de colecciones de referencia y muestras arqueológicas (Babot, Lund y Olmos, 2014, Babot et al., 2017; Musaubach, 2012, 2014, 2015). Posteriormente, llevamos adelante las primeras observaciones microscópicas de los conjuntos fitolíticos según parámetros arqueobotánicos estándar consignando la presencia/ausencia de los morfotipos incluidos en la Nomina conservanda del ICPN (Madella, Alexandre y Ball, 2005), es decir, los tipos cross (cruciforme), saddle y rondel, así como bilobate (bilobados). Estas constituyen las morfologías clásicas en la descripción de las asociaciones fitolíticas presentes en Poáceas y la base sobre la cual se construyó la clasificación de Twiss et al. (1969) a nivel de subfamilia y trabajos posteriores de otros autores como los de Mulholland y Rapp (1989), los cuales luego son retomados para construir los índices de reconstrucciones paleoambientales. Los otros morfotipos consignados fueron elongates (elongados) y trichome (tricomas) y trichome base (base de tricomas). Agrupamos los conjuntos presentes en nuestra muestra de calcinados de inflorescencias e infrutescencia según sus subfamilias de origen, a saber: Pooideae, Danthonoideae, Chloridoideae y Panicoideae.
Tabla 1. Exicata de los especímenes muestreados # Nombre científico aceptado actualmente.
Table 1. Exicata of sampled specimens # Scientific name currently accepted.
Siguiendo nuestro protocolo de trabajo también definimos y registramos la presencia de tres tipos de formas intermedias en continuos morfológicos: rondel/ saddle; bilobado/rondel, rondel/trapeziform y saddle normal/rondel. El concepto de continuo morfológico hace referencia a una serie de morfologías muy próximas entre sí que ocurren entre dos límites definidos, los cuales en este caso estarían dados por dos clases establecidas de fitolitos o morfotipos (adaptado de Babot et al., 2017). Al aplicar una técnica de bajo impacto para extracción de los fitolitos, pudimos describir los tipos de fitolitos articulados pluricelulares (Zucol, 2014) presentes que conforman fragmentos de tejidos silicificados (tejidos vasculares, de estructura granular indiferenciado, subepidérmicos y complejos estomáticos).
Resultados
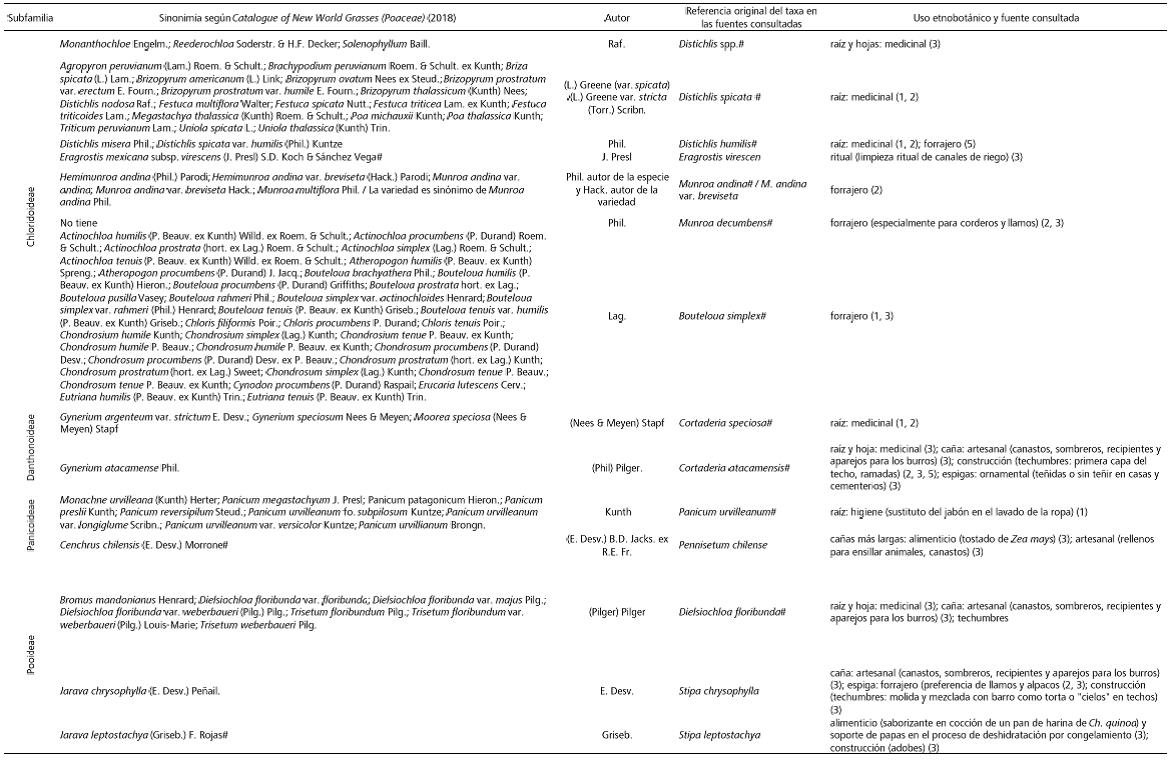
En la Tabla 2 se sintetizan los datos etnobotánicos relevados para la Puna argentina y Norte de Chile acerca de los usos tradicionales de las gramíneas silvestres, proveyendo las sinonimias correspondientes. Las clases de usos registrados para 31 especies incluyen los higiénicos, medicinales, artesanales, instrumentales, alimenticios, forrajeros, rituales, ornamentales y en la construcción. Las partes de la planta implicadas en ellos varían desde el uso de la mata completa o despojada de algunas secciones, hasta las hojas, cañas, pedúnculos, raíces o espigas por separado.
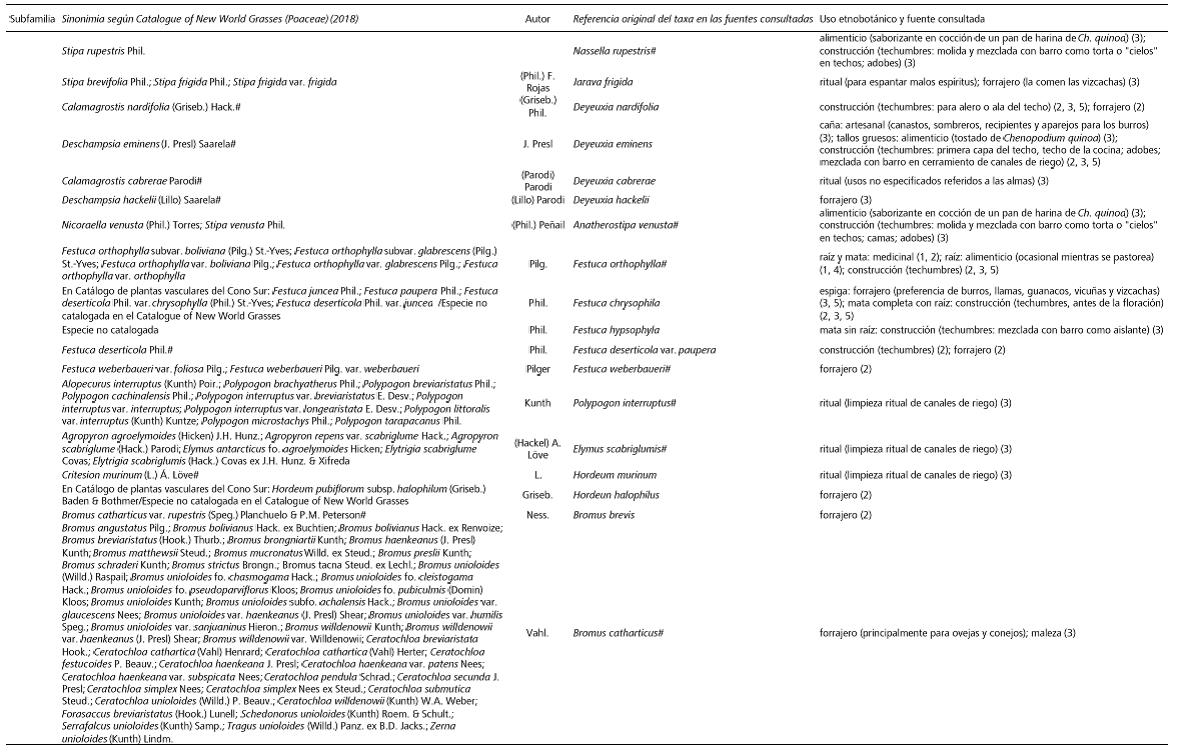
Tabla 2 a y b . Registros etnobotánicos acerca de la utilidad de las gramíneas silvestres nativas de la Puna argentina y chilena. Referencias: (1) Pérez, 2006, (2) Olivera, 2006, (3) Villagrán y Castro, 2003, (4) Lund, 2016, (5) Aldunate et al., 1981. # Nombre científico aceptado actualmente.
Table 2 a y b. Ethnobotanical records about the usefulness of the native wild grasses of the Argentinian and Chilean Puna. References: (1) Pérez, 2006, (2) Olivera, 2006, (3) Villagrán & Castro, 2003, (4) Lund, 2016, (5) Aldunate et al., 1981. # Cientific name currently acepted.
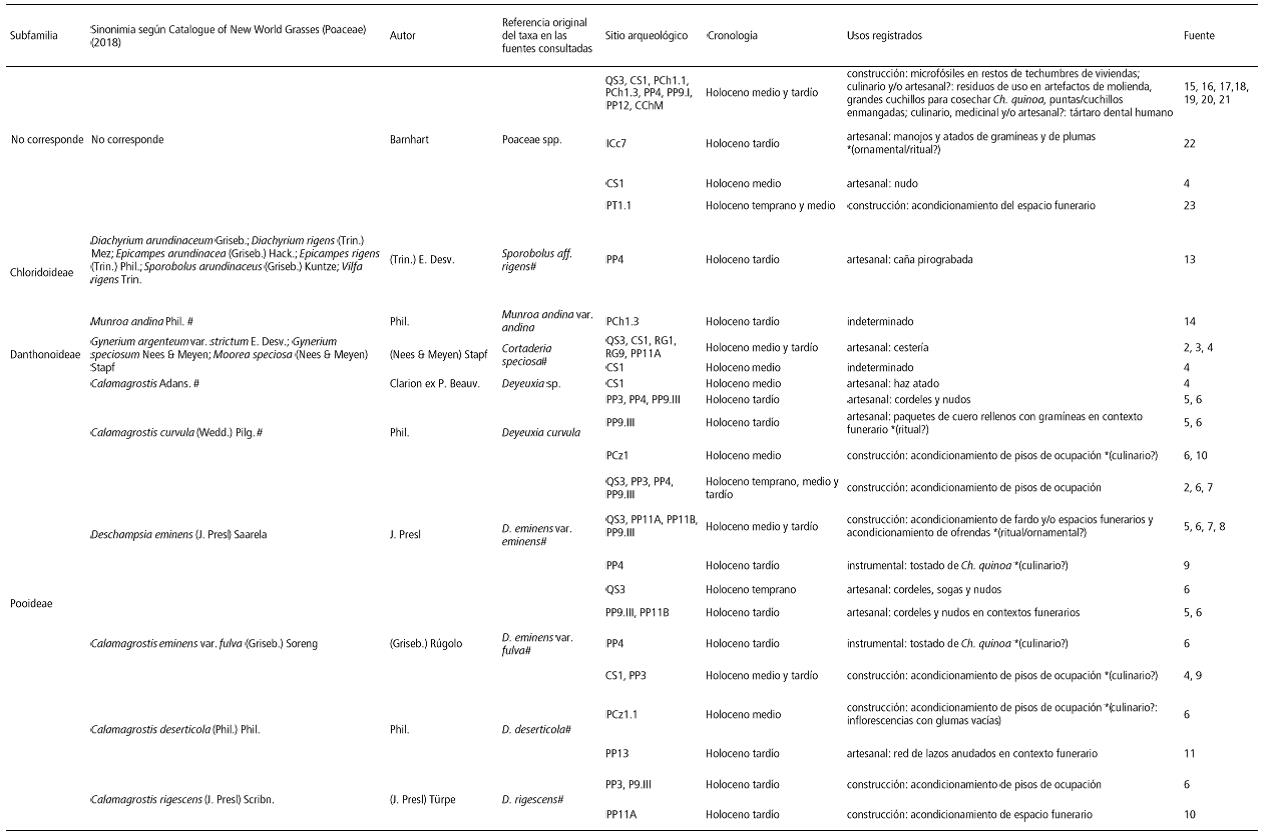
Por su parte, el registro arqueológico de la Puna argentina que proporciona evidencias del uso de las gramíneas silvestres a lo largo de todo el Holoceno se resume en la Tabla 3. Este incluye 14 especies locales, una de ellas identificada al nivel de dos variedades y dos especies exóticas, todas con su nomenclatura original publicada en los antecedentes revisados y la correspondiente sinonimia. Los usos asignados a ellas varían entre artesanales, constructivos e instrumentales. Sobre la base de los registros etnográficos y según la parte de la parte preservada en los contextos, podrían proponerse usos alternativos en la ingesta alimenticia o no alimenticia y ritual en algunos casos indicados (marcados con * en la Tabla 3).
Tabla 3 a y b. Poáceas silvestres en el registro arqueológico puneño. Abreviaturas de los sitios: Peñas Chicas 1.1 (PCh1.1), Peñas Chicas 1.5 (PCh1.5), Quebrada Seca 3 (QS3), Cueva Salamanca 1 (CS1), Real Grande 1 (RG1), Real Grande 9 (RG9), Punta de la Peña 3 (PP3), Punta de la Peña 4 (PP4), Punta de la Peña 9.I (PP9.I), Punta de la Peña 9.III (PP9.III), Punta de la Peña 11A (PP11A), Punta de la Peña 11B (PP11B), Punta de la Peña 12 (PP12), Punta de la Peña 13 (PP13), Peñas de la Cruz 1.1 (PCz1.1), Peñas de las Trampas 1.1 (PT1), Casa Chávez Montículos (CChM1), Inca Cueva cueva 4 (ICc4), Inca Cueva cueva 7 (ICc7). Holoceno temprano: ca. 10000-8000 AP, Holoceno medio ca. 8000-4500 AP, Holoceno temprano ca. 4500 AP-presente. Referencias: (1) Aschero y Hocsman, 2011, (2) Aschero, 2010, (3) Pérez de Micou y Ancíbor, 1994 (4) Pintar y Rodríguez, 2015, (5) López Campeny, 2006-2007, (6) Rodríguez et al., 2003, (7) Rodríguez y Rúgolo de Agrasar, 1999, (8) Gonzalez Baroni et al., 2017, (9) Rodríguez et al., 2006, (10) Rodríguez, 2013b, (11) López Campeny et al., 2014, (12) Rodríguez, 1999, (13) Rodríguez, 2008, (14) Aguirre, 2005, (15) Babot et al., 2012, (16) Babot, 2009, (17) Babot, 2011, (18) Babot, 2016, (19) Escola et al., 2013, (20) Babot et al., 2013, (21) Gonzalez Baroni y Babot, 2013, (22) Pérez de Micou, López Campeny y Lessa Costa, 2016, (23) Martínez, 2013, (24) Rodríguez y Martínez, 2001, (25) Mondini et al., 2013. # Nombre científico aceptado actualmente. * Posible uso alternativo propuesto, afín a referencias etnográficas y según la parte de la planta representada, no considerado en la fuente arqueológica original.
Table 3 a y b. Wild Poaceae in the Punean archaeologial record. Abbreviations of the sites: Peñas Chicas 1.1 (PCh1.1), Peñas Chicas 1.5 (PCh1.5), Quebrada Seca 3 (QS3), Cueva Salamanca 1 (CS1), Real Grande 1 (RG1), Real Grande 9 (RG9), Punta de la Peña 3 (PP3), Punta de la Peña 4 (PP4), Punta de la Peña 9.I (PP9.I), Punta de la Peña 9.III (PP9.III), Punta de la Peña 11A (PP11A), Punta de la Peña 11B (PP11B), Punta de la Peña 12 (PP12), Punta de la Peña 13 (PP13), Peñas de la Cruz 1.1 (PCz1.1), Peñas de las Trampas 1.1 (PT1), Casa Chávez Montículos (CChM1), Inca Cueva cueva 4 (ICc4), Inca Cueva cueva 7 (ICc7). Holoceno temprano: ca. 10000-8000 AP, Holoceno medio ca. 8000-4500 AP, Holoceno temprano ca. 4500 AP-presente. References: (1) Aschero & Hocsman, 2011, (2) Aschero, 2010, (3) Pérez de Micou & Ancíbor, 1994 (4) Pintar y Rodríguez, 2015, (5) López Campeny, 2006-2007, (6) Rodríguez et al., 2003, (7) Rodríguez & Rúgolo de Agrasar, 1999, (8) Gonzalez Baroni et al., 2017, (9) Rodríguez et al., 2006, (10) Rodríguez, 2013b, (11) López Campeny et al., 2014, (12) Rodríguez, 1999, (13) Rodríguez, 2008, (14) Aguirre, 2005, (15) Babot et al., 2012, (16) Babot, 2009, (17) Babot, 2011, (18) Babot, 2016, (19) Escola et al., 2013, (20) Babot et al., 2013, (21) Gonzalez Baroni & Babot, 2013, (22) Pérez de Micou, López Campeny & Lessa Costa, 2016, (23) Martínez, 2013, (24) Rodríguez & Martínez, 2001, (25) Mondini et al., 2013. # Cientific name currently acepted. * Possible alternative use proposed, related to ethnographic references and according to the part of the plant represented, not considered in the original archaeological source.
De manera preliminar el estudio de especies de Poáceas silvestres de la Puna agrupadas en 4 subfamilias (Chloridoideae, Danthonoideae, Panicoideae y Pooideae) (Tabla 4) muestra la presencia de un conjunto de morfologías atribuibles a la silicificación de células cortas, células largas, elementos epidérmicos y subepidérmicos y de conducción que se distribuyen de manera diferencial entre las subfamilias. Estos morfotipos se discuten aquí preliminarmente en términos de su presencia/ausencia en las especies estudiadas y agrupadas en subfamilias, pero la producción absoluta de cada subfamilia podrá ser establecida con precisión en el futuro al ampliar el número de taxones analizados.
Tabla 4. Presencia y ausencia de morfotipos fitolíticos en las inflorescencias e infrutescencias de las subfamilias analizadas.
Table 4. Presence and absence of phytolithic morphotypes in the inflorescences and infrutescences of the subfamilies analyzed.
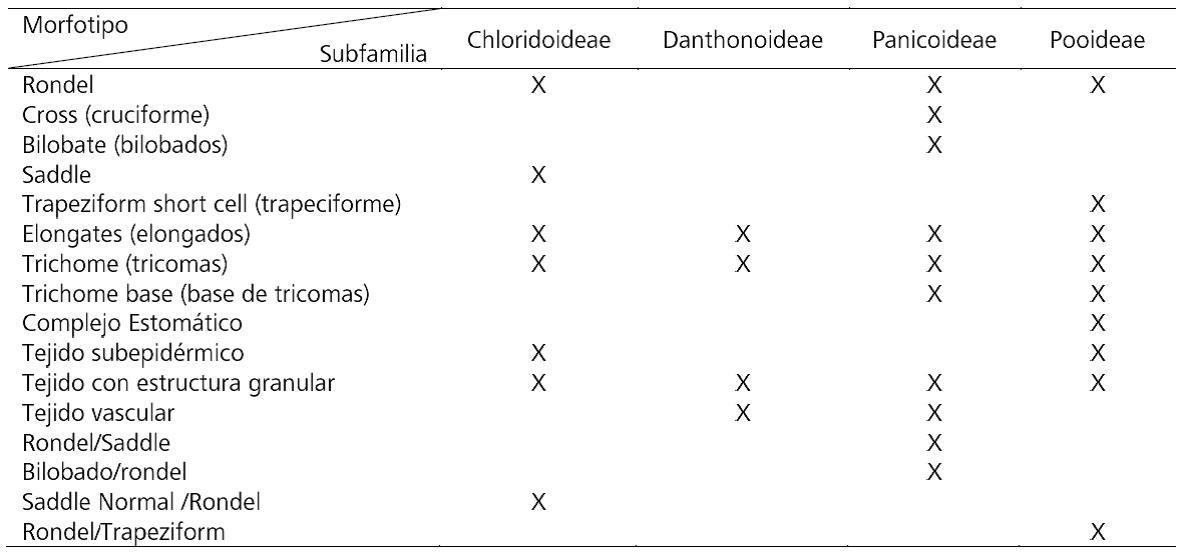
Entre los morfotipos correspondientes a células cortas, se aprecia la presencia de tres formas incluidas en la nómina conservanda ICPN (Madella et al., 2005), a saber: rondel, cross (cruciforme) y saddle, y otras dos que según la nomenclatura actual corresponden a bilobate short cell (bilobado o bilobate), y trapeziform short cell (trapeciforme o square/rectangle). En general, estas morfologías tienen un recuento bajo, aunque destaca la presencia del morfotipo rondel en especies de Pooideae y Chloridoideae y de la clase saddle en las Chloridoideae. También se encuentran formas intermedias (continuo morfológico) entre estos morfotipos: rondel/saddle y bilobado/rondel se aprecian en especies de Panicoideae, rondel/trapeziform se observa en Pooideae, mientras que la forma saddle normal/rondel está presente en la subfamilia Chloridoideae (Tabla 4 y Figura 1). Asimismo, se registran fitolitos de la clase elongate long cell (elongados), dominantemente y como forma principal en especies de Pooideae, y con buena representación en las subfamilias Chloridoideae, Panicoideae y Danthonioideae. Los elementos epidérmicos incluyen morfotipos correspondientes a trichome (tricoma) con una buena representación en la muestra, excepto en Panicoideas; trichome base (base de tricoma) en Pooideas y Panicoideas y stomate (complejo estomatocístico) en Pooideas. Finalmente, en mucha menor medida se encuentran silicificaciones en tejidos subepidérmicos (Chloridoideae y Pooideae), tejidos vasculares de la clase vascular cell or tissue (Danthonioideae y Panicoideae) y con estructura granular o granulate (en todas las subfamilias).

Figura 1. Morfotipos fitolíticos registrados en material de referencia moderno de las subfamilias Pooideae, Danthonoideae, Chloridoideae y Panicoideae. Escala 20 µm. Referencias: A. Rondel/Trapeziform, B. Saddle Normal /Rondel, C. Bilobado/rondel, D. Bilobado/rondel, E. Rondel/ Saddle, F. Tejido con estructura granular, G. Tejido subepidérmico, H. Tejido subepidérmico y Complejo Estomático, I. Tejido con estructura granular, J. Tejido vascular y tejido con estructura granular, K. Tejido subepidérmico, L. Tejido con estructura granular, M. Trapeziform short cell (trapeciforme), N. Rondel, O. Elongates (elongados), Trichome (tricomas) y Trichome base (base de tricomas), P. Trichome (tricomas), Q. Elongates (elongados), R. Trichome (tricomas), S. Saddle, T. Rondel, U. Elongates (elongados), V. Cross (cruciforme), W. Rondel, X. Trichome (tricomas), Y. Bilobate (bilobados), Z. Elongates (elongados) y Bilobate (bilobados).
Figure 1. Phytolithic morphotypes in the comparative current material of the Pooideae, Danthonoideae, Chloridoideae and Panicoideae subfamilies. Scale bar 20 µm. References: A. Rondel/Trapeziform, B. Saddle Normal /Rondel, C. Bilobado/rondel, D. Bilobado/rondel, E. Rondel/Saddle, F. Tejido con estructura granular, G. Tejido subepidérmico, H. Tejido subepidérmico y Complejo Estomático, I. Tejido con estructura granular, J. Tejido vascular y tejido con estructura granular, K. Tejido subepidérmico, L. Tejido con estructura granular, M. Trapeziform short cell (trapeciforme), N. Rondel, O. Elongates (elongados), Trichome (tricomas) y Trichome base (base de tricomas), P. Trichome (tricomas), Q. Elongates (elongados), R. Trichome (tricomas), S. Saddle, T. Rondel, U. Elongates (elongados), V. Cross (cruciforme), W. Rondel, X. Trichome (tricomas), Y. Bilobate (bilobados), Z. Elongates (elongados) y Bilobate (bilobados).
Discusiones y consideraciones finales
Analizando el registro etnobotánico y arqueológico en la Puna Numerosas referencias etnobotánicas de la Puna argentina y chilena indican la persistencia del uso de las gramíneas nativas de este ambiente árido hasta tiempos modernos. La mayoría de las aplicaciones actuales se refieren a la selección de las partes vegetativas (hojas y tallos o cañas), principalmente, y de las raíces en menor medida. Las partes reproductivas (espigas y panojas) se mencionan sólo eventualmente en relación al forrajeo animal y a los usos ornamentales o festivos. Mientras que algunos usos marcan fuertes tradiciones entre el pasado arqueológico y el presente, otros registros arqueológicos no se verifican en la actualidad, constituyendo conocimientos y elecciones que se perdieron con el paso del tiempo, o que no forman parte de los relevamientos existentes. Entre estos, podrían encontrarse aquellos referidos al consumo humano de las semillas silvestres, eventualmente reemplazados por granos domesticados (maíz, quinua, amarantos, por ejemplo) y en tiempos más recientes, también por granos de origen no americano (trigo, cebada, arroz, por ejemplo), dado que las referencias etnobotánicas a estas prácticas no se registran en el área. No obstante, sí se verifican usos alimenticios indirectos, tales como el empleo de los tallos o cañas y raíces de ciertas gramíneas como saborizantes (Villagrán y Castro, 2003).
Ciertos hallazgos arqueológicos macrobotánicos de Poáceas de esta área llaman la atención pues podrían vincularse a restos de colectas con fines alimenticios en espacios residenciales, debido específicamente a la presencia de espiguillas y panojas. Estos podrían ser los casos de D. eminens J. Presl. var. eminens (pasto huailla, huaya, iru, sora sora) (Rodríguez y Rúgolo de Agrasar, 1999) y de D. eminens var. fulva (pasto de vega, cebadilla de vicuña), en distintos sitios en que se recuperaron inflorescencias incompletas donde se aprecian ramificaciones laterales con espiguillas dispuestas en su parte distal (Rodríguez, 2013b). También destaca el registro de grandes acumulaciones de D. deserticola sin órganos reproductivos (presentando las glumas vacías) (Rodríguez, 2013b).
De igual modo, pueden proponerse hipótesis referidas a los usos ornamentales o rituales de estas plantas como en los casos que involucran el uso de taxones que actualmente intervienen en actos propiciatorios en los Andes, a los que hemos aludido previamente. Entre ellos, podrían incluirse los atados y manojos de gramíneas y plumas (Pérez de Micou, López Campeny y Lessa Costa, 2016), los paquetes de cuero rellenos con gramíneas como acompañamiento mortuorio (López Campeny 2006-2007) y la propia selección de determinadas especies en el acondicionamiento del espacio funerario.
El análisis de estos registros permite visibilizar y problematizar el rol potencial de las Poáceas en distintas funciones previamente no consideradas, en particular, en la alimentación antigua de esta parte de Sudamérica. El tratamiento de las sinonimias potencia la comparabilidad entre los distintos cuerpos de datos, mientras que la vía del análisis de microindicadores abre una puerta para nuevas indagaciones sobre ellos.
Primeras implicancias de la aproximación de microindicadores
A partir del estudio de un conjunto de especies de Poáceas silvestres de la Puna agrupadas en 4 subfamilias (Chloridoideae, Danthonoideae, Panicoideae y Pooideae) destacamos los siguientes aspectos relevantes. El primero de ellos se refiere a que los tipos globales de las células cortas, es decir, los morfotipos rondel, bilobate, cross, saddle y trapeziform, identificados en inflorescencias, infrutescencias y ejes, coinciden a nivel de subfamilia con los tipos esperados y mencionados en hojas de especies de la subfamilia (ver por ejemplo Twiss et al., 1969; Mulholland y Rapp, 1989; Gallego, Distel, Camina y Rodríguez Iglesias, 2004; Fernandez Honaine, Bernava Laborde y Zucol, 2008). Lo mismo ocurre con los morfotipos menos diagnósticos de las clases elongate long cell, trichome, trichome base, stomate y vascular cello o tissue, entre otros. Por lo tanto, consideramos necesario realizar un estudio que detalle las variaciones presentes dentro de las morfologías globales para establecer si es posible diferenciar estos órganos a partir de subclases de cada uno de los morfotipos registrados. Esto pone en evidencia el fenómeno de redundancia que caracteriza a los fitolitos, definido para indicar la recurrencia de morfologías similares en diferentes taxones (Rovner, 1971; Piperno, 2008) y en este caso, también en diferentes partes anatómicas de la planta. A la vez, podemos apelar al término conjuntos fitolíticos redundantes para referirnos a la indiferenciación entre las clases fitolíticas descriptas en diferentes órganos del mismo taxón agregando el apelativo aparentes cuando no existe un estudio morfométrico y/o estadístico detallado de la variación de formas. Por lo tanto, mientras esto no se establezca, la identificación a nivel de órgano de la planta (hoja vs. inflorescencia/infrutescencia) debe ser realizada con cautela. Incluso, sostenemos que en la caracterización del conjunto de fitolitos de un taxón o de una parte anatómica de ese taxón, es preciso considerar a los morfotipos que son típicos en tales conjuntos sean éstos diagnósticos o solo característicos. Ello permite obtener una visión más completa de la producción de fitolitos de cada taxón al trabajar con muestras modernas (Korstanje y Babot, 2007) y a su vez, fortalece la asignación de los fitolitos en registros fragmentarios como los que corresponden a contextos antiguos.
Si bien en el estado actual de esta investigación la discriminación de partes de la planta aún no es posible, los resultados obtenidos permiten visibilizar y comenzar a discutir el rol potencial de las Poáceas en distintas funciones, en particular, en la alimentación antigua de esta parte de Sudamérica. A su vez, la diferenciación de los morfotipos originados en distintos taxones de esta familia, permitirá a futuro una identificación de aquellos taxones elegidos para el consumo alimenticio de sus cariopses, o bien para otros usos de las inflorescencias.
En este trabajo también abordamos la variación morfológica que puede ser observada como resultado de la silicificación de células contiguas de un mismo origen anatómico, la que queda tipificada a través del concepto de continuo morfológico (Babot, Musaubach y Gonzalez, 2016). Al describir las morfologías de células cortas propuesta por el ICPN (Madella et al., 2005) se identificaron formas intermedias que corresponden a un continuo originado en la variación biológica natural. Esto había sido registrado previamente en el estudio de inflorescencias/infrutescencias de gramíneas de ambientes semiáridos pampeanos (Babot et al., 2017). El fenómeno queda especialmente retratado al observar la variación de morfotipos correspondientes a células cortas que pueden ser recuperadas como fitolitos articulados pluricelulares. Estos incluyen dos o más elementos celulares vinculados o en conexión orgánica (Zucol, 2014) y, por lo tanto, la variación entre dos o más formas contiguas en el espacio puede ser claramente registrada. No obstante, tal variación también se verifica en las mismas muestras en los fitolitos unicelulares. De este modo, la situación descripta es esperable en los contextos antiguos y, por lo tanto, debe ser considerada, correctamente registrada e informada.
Recientemente, Zucol (2014) revisó las designaciones e implicancias descriptivas y clasificatorias de los fitolitos articulados pluricelulares. éstos remiten a un nivel de organización diferente al del fitolito unicelular que corresponde a células individuales, pues se originan en la silicificación de una parte del tejido vegetal (espacio intra y/o intercelular). Según el autor, los fitolitos unicelulares corresponden a los denominados fitolitos no articulados (Zucol, 1995) o single phytoliths (PHYTCORE 2016), mientras que los fitolitos articulados (Zucol, 1995) constituyen los así llamados multicellular structures (PHYTCORE, 2016), silicified tissue (Runge, 1999) y plate o pavement plate (Wallis, 2003). Los fitolitos articulados pueden resultar de la biomineralización de células de igual o diferente origen anatómico y, por lo tanto, contener uno o más morfotipos vinculados. Según Zucol (2014) los fitolitos de origen pluricelular que han experimentado las fases fosildiagenéticas pueden ser considerados como microfósiles y en tal sentido se les puede aplicar los principios, reglas y recomendaciones regidas por el International Code of Nomenclature for Algae, Fungi, and Plants (Turland et al. 2018), así como las provisiones del ICPN (Madella et al., 2005), tal como se observa en el ejemplo proporcionado por Prasad et al. (2011). Finalmente, como Zucol (2014) ha sugerido, es importante considerar y describir a los fitolitos pluricelulares que corresponden a fragmentos de tejidos y por lo tanto representan un nivel diferente de organización que los fitolitos unicelulares. Así, sería interesante que la variación morfológica pudiera quedar reflejada en estos casos excepcionales de silicificación, al constituir un dato relevante que caracteriza al taxón.
Las morfologías intermedias o mixtas entre las que se encuentran las que hemos registrado aquí, rondel/saddle, bilobate/rondel, rondel/trapeziform y saddle normal/ rondel, han sido reconocidas y citadas en la literatura en otras oportunidades, pero su problemática no ha recibido mayor tratamiento. En algunos casos la indefinición se expresa con el sufijo "pseudo" o con el uso de la preposición inglesa "to", en un esfuerzo descriptivo para incluirlas en algunos de los grupos más clásicos. Si bien no se refirieron al continuo morfológico dentro de un mismo taxón que puede ser seguido al observar fitolitos pluricelulares, Piperno y Pearsall (1998) sí se expresaron con respecto a las que aquí llamamos formas intermedias indicando con una "/" la transición entre dos morfotipos clásicos en las formas frecuentes en especies de la subfamilia Arundinoideae. Como se desprende de este ejemplo, las implicancias clasificatorias no son menores por cuanto es a partir de las formas y sus recuentos, que luego se discute la identificación taxonómica y que se determina la presencia de subfamilias en los conjuntos antiguos. Como indican Piperno y Pearsall (1998), el uso de categorías mixtas permite dejar expresada la ocurrencia de formas intermedias; en su defecto, estas quedan incluidas en una categoría ad hoc, denominada "Otras".
Así, las variaciones morfológicas dentro un mismo morfotipo de célula corta y las afinidades entre morfotipos similares, que están implicadas en el concepto de continuo morfológico, invitan a reflexionar sobre cuánto son forzadas a encajar las morfologías intermedias observadas en nuestras muestras antiguas y modernas, dentro de un grupo acotado de morfotipos definidos, debido a que las clasificaciones, por caso las de las Poáceas, han apelado únicamente a frecuencias de morfotipos estándar. Si bien estas clasificaciones apelan a tendencias con significado taxonómico real, en especial al promediar un conteo importante de ejemplares, de cierta manera, desdibujan la variación en la producción de formas y dejan librado a criterio del investigador la asignación de las morfologías no claras en uno u otro tipo morfológico estándar, o bien a no contabilizarlas. La principal limitante se verifica al asignar conjuntos antiguos con un insuficiente conteo de morfotipos o cuando estos son ambiguos por su indefinición.
Es así que, atendiendo a los aspectos discutidos, profundizar el estudio de la colección de referencia de Poáceas silvestres de ambientes áridos y semiáridos, permitirá abordar a futuro el consumo, la ingesta y las técnicas de procesamiento (prácticas post colecta) de las partes útiles de las gramíneas, así como otros usos de los granos e inflorescencias, a partir de la recuperación de microfósiles en residuos de la utilización de artefactos, en el tártaro dental y en sedimentos, entre otros. Tales aspectos aún constituyen incógnitas sin resolución en esos ámbitos, siendo de gran interés el uso alimenticio de estas plantas en el pasado.
1. En esta sección se mantienen los nombres científicos según la cita original de los autores a los fines de facilitar el acceso a los datos básicos en las fuentes de antecedentes que hemos consultado. La sinonimia se proporciona en la sección de resultados.
2. Si bien no se tienen referencias acerca del uso alimenticio de las cariopses de especies de los géneros Bromus y Chusquea, de interés para el caso de estudio, estos han sido mencionados en otros registros atinentes al Cono Sur americano (Babot et al., 2017).
3. En esta sección se mantienen los nombres científicos según la cita original de los autores a los fines de facilitar el acceso a los datos básicos en las fuentes de antecedentes que hemos consultado. La sinonimia se proporciona en la sección de resultados.
Agradecimientos
Agradecemos muy especialmente a la Dra. Anabela Plos del Herbario del Museo de La Plata, quien nos colaboró en la selección de los ejemplares muestreados y el control de las sinonimias y a la Lic. Mónica Burgos y Srta. Julieta Zapatiel del Laboratorio de Arqueobotánica, IAM, FCN e IML por su inestimable contribución en los procesos de laboratorio. Al Jefe de la División Plantas Vasculares, Dr. J. V. Crisci por permitir el acceso al material óptico del Herbario del Museo de la Plata. A los Proyectos PIPCONICET Nº577 (Dir. C. Aschero y P. Babot) y PICT 20131703 (Dir. S. Hocsman).
Bibliografía
1. Aguirre, M. G. (2005). Arqueobotánica de Peñas Chicas 1.3 Antofagasta de la Sierra, Catamarca (tesis de Grado inédita). Universidad Nacional de Tucumán, San Miguel de Tucumán. Argentina. [ Links ]
2. Aldunate, C., Armesto J., Castro V. y Villagrán C. (1981). Estudio etnobotánico en una comunidad precordillerana de Antofagasta: Toconce. Boletín del Museo Nacional de Historia Natural, 38, 183-223. [ Links ]
3. Aschero, C. A. (2010). Arqueologías de Puna y Patagonia centro-meridional: comentarios generales y aporte al estudio de los cazadores-recolectores puneños en los proyectos dirigidos desde el IAM (1991-2009), en P. Arenas, C. Aschero y C. Taboada (eds.), Rastros en el camino.Trayectos e Identidades de una Institución. Homenaje a los 80 años del IAM-UNT, pp. 257-293. San Miguel de Tucumán: Instituto de Arqueología y Museo, EDUNT. [ Links ]
4. Aschero, C.A. y S. Hocsman 2011. Arqueología de las ocupaciones cazadoras-recolectoras de fines del Holoceno Medio de Antofagasta de la Sierra (Puna Meridional Argentina). Chungará 43 (Volumen Especial 1): 393-411. [ Links ]
5. Babot, M. P. (2004). Tecnología y utilización de artefactos de molienda en el Noroeste Prehispánico. Tesis de Doctorado en Arqueología - Universidad Nacional de Tucumán, San Miguel de Tucumán, 2004. [ Links ]
6. Babot, M. P. (2009). La cocina, el taller y el ritual. Explorando las trayectorias del procesamiento vegetal en el Noroeste Argentino. Darwiniana, 47 (1), 7-30. [ Links ]
7. Babot, M. P. (2011). Cazadores-recolectores de los Andes Centro-Sur y procesamiento vegetal. Una discusión desde la Puna Meridional Argentina (ca. 7000-3200 años A.P.). Chungara, Revista de Antropología Chilena, 43 (1), 413-432. [ Links ]
8. Babot, M. P. (2016). Notas sobre la arqueología de las plantas en un desierto de altura. Cuadernos do Lepaarq. Textos de Antropologia, Arqueologia e Patrimônio 13(25), 333-365. [ Links ]
9. Babot, M. P., Cattáneo, G. R. y Hocsman, S. (2013). Assessing the life history of projectile points/ knives from the Middle Holocene of Argentina's Southern Puna. Quaternary International, 287, 3-19. [ Links ]
10. Babot, M. P., Hocsman, S., Piccón Figueroa, R. E. y Haros, M. C. (2012). Recetarios prehispánicos y tradiciones culinarias. Casos de la Puna argentina. En (Babot, M. del P.; M. Marschoff y F. Pazzarelli, eds.) Las manos en la masa. Arqueologías, Antropologías e Historias de la Alimentación en Suramérica, pp. 235-269. Museo de Antropología, Universidad Nacional de Córdoba), Córdoba. [ Links ]
11. Babot, M. P., Lund, J. y Olmos, A.V. (2014). Taphonomy in the kitchen: culinary practices and processing residues of native tuberous plants of the South-Central Andes. Intersecciones en Antropología, Special Issue Nº 1 Taphonomic Approaches to the Archaeological Record, 35-53. [ Links ]
12. Babot, M. P., Lund, J. y Olmos, A. V. (2016). Experiencias y relatos verdes compartidos con los vecinos antofagasteños. Un aporte reflexivo desde la etno y arqueobotánica en la puna de Catamarca. En Actas del XIX Congreso Nacional de Arqueología Argentina. Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Serie Monográfica y Didáctica 54: 1005-1010. San Miguel de Tucumán, Tucumán. [ Links ]
13. Babot, M. P., Musaubach, M. G. y Gonzalez, J. A. 2016. Continuo morfológico y fitolitos 3D. Aportes desde una perspectiva arqueobotánica para definir conjuntos fitolíticos. En Zucol, A. F., Patterer, N.I., Colobig, M.M. y E. Moya (eds.). Taller de Micropaleoetnobotánica. Relevancia de una Red Interdisciplinaria de Investigaciones en Fitolitos y Almidones, Libro de Resúmenes, 79. [ Links ]
14. Babot, M. P., Musaubach, M. G. y Plos, A. 2017. An archaeobotanical perspective in the study of inflorescence phytoliths of wild grasses from arid and semi-arid environments of Argentina. Quaternary International 434, 129-141. DOI: 10.1016/j.quaint.2016.02.045. [ Links ]
15. Burkart, R. B., Sánchez, N. O., Gómez, R. O., Sanchez, D.A., Boloy, R.C. y E Czerwenka, J. (1983). Eco-regiones de la Argentina (No. 333.73 E19). Buenos Aires: Secretaría de Recursos Naturales y Desarrollo Sustentable, Programa Institucional Ambiental Administración de Parques Nacionales, Buenos Aires (Argentina). [ Links ]
16. Cabrera, á. L. (1976). Regiones fitogeográficas argentinas. Enciclopedia Argentina de Agricultura y Jardinería, v. 2, parte 1. Buenos Aires: ACME. [ Links ]
17. Cuello, A. S. (2006). Guía ilustrada de la flora de Antofagasta de la Sierra, Catamarca (Puna Meridional Argentina) (tesis de Licenciatura inédita). Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán. Tucumán, Argentina. [ Links ]
18. Escola, P., Hocsman, S. y Babot, M. P. (2013). Entre las residencias y los campos de cultivo. Aportes de los cuchillos/raederas de módulo grandísimo a la cuestión del laboreo agrícola en Antofagasta de la Sierra (Puna de Catamarca) durante el primer milenio dC. Relaciones de la Sociedad Argentina de Antropología 38(1), 83-110. [ Links ]
19. Fernandez Honaine, M., Bernava Laborde, V., Zucol, A. (2008). Contenido de sílice en gramíneas del pastizal nativo del sudeste bonaerense. En: Korstanje, M.A., Babot, M. P. (Eds.), Matices Interdisciplinarios en Estudios Fitolíticos y de Otros Microfósiles pp. 57-63. Capítulo 6. Oxford: BAR International Series S1870. [ Links ]
20. Gallego, L., Distel, R. A., Camina, R., Rodríguez Iglesias, R.M. (2004). Soil phytoliths as evidence for species replacement in grazed rangelands of central Argentina. Ecography, 27, 725-732. [ Links ]
21. Gonzalez Baroni, L. G. y Babot, M. P. (2013). Caracterización de los microfósiles en tártaro dental de individuos del primer milenio A.D. de la Puna Argentina. En: Anais do Museu Nacional-UFRJ, Serie Livros 49: 227. Resúmenes del 5º Encontro Latinoamericano de Fitólitos. Rio de Janeiro: Museu Nacional, Universidade Federal do Rio de Janeiro. [ Links ]
22. Gonzalez Baroni, L. G.; Hocsman, S.; Aschero, C. A. (2017). Prácticas mortuorias de cazadores-recolectores tardíos de los desiertos de altura del Noroeste Argentino: El fardo funerario de Quebrada Seca 3. Revista Argentina de Antropología Biológica, 19, 1-18. [ Links ]
23. Grana, M. L., Tchilinguirian, P., Hocsman, S., Escola, P. S. y Maidana, N. (2016). Paleohydrological changes in highland desert rivers and human occupation, 7000-3000 cal. yr BP, South-Central Andes, Argentina. Geoarchaeology. An International Journal, 31, 412-433. [ Links ]
24. Haber, A. (1992). Pastores y pasturas. Recursos forrajeros en Antofagasta de la Sierra (Catamarca) en relación a la ocupación Formativa. Shincal, 2, 15-23. [ Links ]
25. Hocsman, S. y Babot, M. P. (2018) La transición de cazadoresrecolectores a sociedades agro-pastoriles en Antofagasta de la Sierra (Puna de Catamarca, Argentina): Perspectivas desde la agencia y las prácticas. Chungara, Revista de Antropología Chilena 50(1): 51-70. [ Links ]
26. Korstanje, M. A. y Babot, M. P. (2007). Microfossils Characterization from South Andean Economic Plants. En: Madella, M., Zurro, D., (Eds.) Plant, People and Places: Recent Studies in Phytolith Analysis, pp. 41-72. Cambridge: Oxbow Books. [ Links ]
27. López Campeny, Sara M. L., Andrés S. Romano, M. Fernanda Rodríguez, álvaro R. Martel y Mariano H. Corbalán (2014) De aquí y de allá: análisis integral de un contexto funerario. Vínculos e interacciones sociales entre Puna meridional y Tierras Bajas orientales. Intersecciones en Antropología 15: 201-218. [ Links ]
28. López Campeny, S. M. L. 2006-2007 El poder de torcer, anudar y trenzar a través de los siglos. Textiles y ritual funerario en la Puna Meridional Argentina. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 21: 143-155. [ Links ]
29. Lund, J. (2016). Tubérculos y raíces útiles de la Puna Meridional Argentina. Un abordaje desde la Arqueobotánica y la Etnobotánica en Antofagasta de la Sierra (provincia de Catamarca) (trabajo Final de la Carrera de Arqueología Inédita). Facultad Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional Tucumán. San Miguel de Tucumán, Argentina. [ Links ]
30. Lund, J. y Babot, M. P. (2014). Plantas tuberosas y raíces en la Puna de Catamarca, Argentina. Los pobladores y el uso actual, microfósiles y experimentación para su identificación en contextos pasados. En: Libro de Resúmenes del Taller Micropaleoetnobotánica: Relevancia de una red interdisciplinaria de investigaciones en fitolitos y almidones, pp. 116. Centro Universitario Regional Este, Universidad de La República. La Paloma. [ Links ]
31. Madella, M., Alexandre, A., Ball, T. (2005). International Code for Phytolith Nomenclature 1.0. Annals of Botany 96, 253-260. [ Links ]
32. Martínez, J.G. (2012). Evidence of early human burials in the Southern Argentinian Puna. Current Research in the Pleistocene Special edition Southbound: Late Pleistocene Peopling of Latin America, 75-78. [ Links ]
33. Mondini, M., Martínez, J. G., Pintar, E. y Reigadas, M. C. (2013). Middle Holocene foraging, mobility and landscape use in the southern argentinean Puna. Hunter-gatherers from Antofagasta de la Sierra, Catamarca, Argentina. En Quaternary International Vol. 307 "Human populations and environments during the mid-Holocene in the South-Central Andes". M. Mondini (Editora invitada). Journal of the International Union for Quaternary Research-Elsevier. Pp.66-73. [ Links ]
34. Mulholand, S. C. y Rapp, G. R. (1989). Characterization of grass phytoliths for archaeological analysis. Material Research Society Bulletin, 14(3), 36-39.
35. Musaubach, M. G. (2012). Potencialidad de estudios arqueobotánicos sobre tártaro dental de cazadores recolectores de la provincia de La Pampa, Argentina. Revista Argentina de Antropología Biológica, 14 Número especial, 105-113. [ Links ]
36. Musaubach, M. G. (2014). Estudios Arqueobotánicos en sociedades cazadoras-recolectoras de ambientes semiáridos. Análisis de microrrestos vegetales en contextos arqueológicos de Pampa Occidental (Argentina) (tesis de doctorado inédita). Facultad de Filosofía y Letras. Universidad de Buenos Aires, Ciudad Autónoma de Buenos Aires, Argentina. [ Links ]
37. Musaubach, M. G. (2015). Microrestos vegetales en contenedores cerámicos. Análisis arqueobotánicos en un contexto de cazadores recolectores de pampa occidental (Argentina). En: Belmar C., Lema, V. (Eds.) Avances y desafíos metodológicos en arqueobotánica: miradas consensuadas y diálogos compartidos desde Sudamérica, pp. 278-300. Chile: Facultad de Patrimonio Cultural y Educación. Universidad SEK. [ Links ]
38. Musaubach, M. G., Babot, M. P. y Plos, A. (2013a). Fitolitos de gramíneas silvestres nativas (Poaceae Barnhart) de la Pampa Occidental, Argentina. Utilidad en estudios arqueobotánicos y paleoecológicos. En Anais do Museu Nacional-UFRJ, Serie Livros 49: 229. Resúmenes del 5º Encontro Latinoamericano de Fitólitos, Río de Janeiro, 13 al 16 de Mayo de 2013. Museu Nacional da Universidade Federal do Rio de Janeiro. [ Links ]
39. Musaubach, M. G., Plos, A. y Babot, M. P. (2013b). Differentiation of Maize (Zea mays L.) from native wild grasses based on starch grain morphology. An Archaeobotanical perspective from Central Pampas of Argentina. Journal of Archaeological Science, 40(2), 1186-1193. [ Links ]
40. Olivera, D. E. (1988). La opción productiva: apuntes para el análisis de sistemas adaptativos del Período Formativo del NOA. Precirculados de las ponencias científicas del IX Congreso Nacional de Arqueología Argentina, pp. 83-101. Instituto de Ciencias Antropológicas, UBA, Buenos Aires. [ Links ]
41. Olivera, D. E. (2006). Recursos bióticos y subsistencia en sociedades agropastoriles de la Puna meridional argentina. Comechingonia, Revista de Arqueología, 9, 19-56. [ Links ]
42. Olivera, D. E., Tchilinguirian, P. y Grana, L. (2004). Paleoambiente y arqueología en la Puna Meridional argentina: archivos ambientales, escalas de análisis y registro arqueológico. Relaciones de la Sociedad Argentina de Antropología, 29, 229-248. [ Links ]
43. Pérez, E. L. (2006). Las plantas utilizadas por la comunidad de Antofagasta de la Sierra, Puna catamarqueña, Argentina. (Informe del curso de entrenamiento). Universidad Nacional de Tucumán, San Miguel de Tucumán. [ Links ]
44. Pérez de Micou, C. B. y Ancibor, E. (1994). Manufactura cestera en sitios arqueológicos de Antofagasta de la Sierra, Catamarca. República Argentina. Journal de la Société des Américanistes, Nanterre, 80, 207-216. [ Links ]
45. Pérez de Micou, C. B., López Campeny, S. M. L. y Lessa Costa, R. (2016). Basketry of South America. En: Helaine Selin, (Ed.) Encyclopedia of the history of science, technology, and medicine in non-western cultures. Springer Reference, Springer Science+Business Media, Heidelberg. [ Links ]
46. PHYTCORE. 2016. Phytolith database. http://www.phytcore. org/phytolith/index.php
47. [ Links ] Pintar, E. M. y Rodríguez, M.F. (2015). Understanding foraging radius and mobility in a high desert. Journal of Archaeological Science, 59, 142-158. [ Links ]
48. Piperno, D. (2009). Identifying crop plants with phytoliths (and starch grains) in Central and South America: A review and an update of the evidence. Quaternary International, 193, 146-159. [ Links ]
49. Piperno, D. y Pearsall, D. (1998). The Silica Bodies of Tropical American Grasses: Morphology, Taxonomy, and Implications for Grass Systematics and Fossil Phytolith Identification. Smithsonian Contributions to Botany 85. Washington DC: Smithsonian Institution Press. [ Links ]
50. Podestá, M. M. (1991). Cazadores y pastores de la Puna: apuntes sobre sus manifestaciones de arte rupestre. Shincal, 3,12-16. [ Links ] Prasad, V., Strömberg, C.A.E., Leaché, A.D., Samant, B., Patnaik, R., Tang, L., Mohabey, D.M., Ge, S. y Sahni, A. (2011). Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae. Nature communications 2: 480. DOI: 10.1038/ncomms1482
51. Rodríguez, M. F. (1999). Plant species at an archaeological site of the Southern Argentina Puna (Families: Poaceae, Asteraceae, Fabaceae and Solanaceae). Journal of Ethnobiology 19, 2, 228-247.
52. Rodríguez, M. F. (2008). Recursos vegetales y tecnofacturas en un sitio arqueológico de la Puna Meridional Argentina, área Centro - Sur Andina. Darwiniana 46, 2, 240-257.
53. Rodríguez, M. F. (2013a). Acerca de la flora de Antofagasta de la Sierra, Catamarca, Argentina. Ambiente y Paleoambiente. Darwiniana, 1, 2, 295-323.
54. Rodríguez M. F. (2013b). Los grupos humanos y las plantas en la Puna meridional argentina: arqueobotánica de Antofagasta de la Sierra. Intersecciones en Antropología, 14, 315-339.
55. Rodríguez, M. F. y Martínez, J. G. (2001). Especies vegetales alóctonas como recursos arqueológicos en el ámbito puneño. Publicación Especial de la Asociación Paleontológica Argentina, 8, 139-145.
56. Rodríguez, M. F. y Rúgolo de Agrasar, Z. E. (1999). Deyeuxia eminens (Poaceae: Agrostideae) en un sitio arqueológico de la Puna Meridional Argentina (Provincia de Catamarca). Darwiniana, 37, 229-242.
57. Rodríguez, M. F.; Rúgolo de Agrasar, Z. E. y Aschero C. A. (2003). El género Deyeuxia (Poaceae, Agrostideae) en sitios arqueológicos de la Puna meridional argentina. Provincia de Catamarca. Chungara Revista de Antropología Chilena, 35, 51-72.
58. Rodríguez, M. F., Rúgolo de Agrasar, Z. E. y Aschero, C. A. (2006). El uso de las plantas en unidades domésticas del sitio arqueológico Punta de la Peña 4, Puna meridional argentina. Chungara Revista de Antropología Chilena, 38, 257-271.
59. Rovner, I. (1971). Potential of opal phytoliths for use in paleoecological reconstruction. Quaternary Research, 1: 345359.
60. Runge, F. (1999). The opal phytolith inventory of soils in central Africa quantities, shapes, classification, and spectra. Review of Palaeobotany and Palynology, 107, 23-53.
61. Tchilinguirian, P. y Olivera, D. E. (2014). Late quaternary paleoenvironments, south Andean puna (25º-27º S), Argentina. En Hunter-gatherers from a high-elevation desert: people of the Salt Puna: Northwest Argentina E. Pintar (ed.), pp. 43-69. Oxford, UK: Archaeopress.
62. Turland, N. J., Wiersema, J. H., Barrie, F. R., Greuter, W., Hawksworth, D. L., Herendeen, P. S., Knapp, S., Kusber, W.-H., Li, D.-Z., Marhold, K., May, T. W., McNeill, J., Monro, A. M., Prado, J., Price, M. J. y Smith, G. F. (eds.) (2018). International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159. Glashütten: Koeltz Botanical Books. DOI https://doi. org/10.12705/Code.2018
63. Twiss, P. C. (1987). Grass-opal phytoliths as climatic indicators of the Great Plains Pleistocene. En W. Johnson (Eds.), Kansas Geological Survey Guidebook Series 5, pp. 179-188. Kansas: University of Kansas.
64. Twiss, P. C., Suess, E. y Smith, R. (1969). Morphological Classification of Grass Phytoliths. Soil Science Society of America Proceedings, 33, 1, 109-115.
65. Villagrán, C. y Castro, M. V. (2003). Ciencia Indígena de Los Andes del Norte de Chile. Santiago de Chile: Editorial Universitaria.
66. Wallis, L. (2003). An overview of leaf phytolith production patterns in selected northwest Australian flora. Review of Palaeobotany and Palynology 125, 201-248.
67. Zucol, A. F. 1995. Microfitolitos: II. Análisis de las clasificaciones. Ameghiniana 32 (3): 243-248.
68. Zucol, A. F. 2014. ¿Fitolitos o microfósiles? Consideraciones sobre el tratamiento de fitolitos de origen pluricelular. En: H. Inda (Ed.) Libro de Resúmenes del Taller "Micro paleoetnobotánica". Relevancia de una red interdisciplinaria de investigaciones en fitolitos y almidones, pp. 38-44. Centro Universitario Regional Este, Universidad de la República, Rocha.

