










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista del Museo de Antropología
Print version ISSN 1852-060XOn-line version ISSN 1852-4826
Rev. Mus. Antropol. vol.15 no.2 Córdoba Aug. 2022
http://dx.doi.org/http://doi.org/10.31048/1852.4826.v15.n2.37109
DOI: http://doi.org/10.31048/1852.4826.v15.n2.37109
ANTROPOLOGÍA BIOLOGÍCA
Una aproximación a la reconstrucción de la historia poblacional del Nordeste argentino desde el estudio de los linajes uniparentales
An approach to reconstruction of the population history of the Argentine Northeast from the study of uniparental lineages
Daniela Rodríguez Golpe*, Eliana Aquilano**, Marisol E. Schwab***, Paula Paz Sepúlveda****, Natalia Silvero*****, Mariela Cuello******, José E. Dipierri*******, Emma L. Alfaro Gómez********, Graciela Bailliet*********, Josefina M.B. Motti********** y
Claudio M. Bravi***********
*Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: danielargolpe@gmail.com
**Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: elianaaquilano@gmail.com
*** Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: marisol_schwab@hotmail.com
**** Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: paulabeatrizpaz@gmail.com
***** CADIC, Ushuaia, Argentina. E-mail: silveromarian@gmail.com
****** Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: mcuello@gmail.com
******* Instituto de Biología de la Altura, Facultad de Humanidades y Ciencias Sociales, Universidad Nacional de Jujuy, San Salvador de Jujuy, Jujuy, Argentina. E-mail: jedjujuy@gmail.com
******** Instituto de Biología de la Altura, Facultad de Humanidades y Ciencias Sociales, Universidad Nacional de Jujuy, San Salvador de Jujuy, Jujuy, Argentina. Instituto de Ecorregiones Andinas, Universidad Nacional de Jujuy, San Salvador de Jujuy, Jujuy, Argentina. E-mail: ealfaro@inbial.unju.edu.ar
********* Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: graciela.bailliet@gmail.com
********** Laboratorio de Ecología Evolutiva Humana, CONICET, FACSO-UNCPBA, Quequén, Argentina. E-mail: josemotti@yahoo.com.ar
*********** Laboratorio de Genética Molecular Poblacional, IMBICE (CONICET-CIC-UNLP), La Plata, Argentina. E-mail: cmbravi@fcnym.unlp.edu.ar
Resumen
El Nordeste argentino se caracteriza por la presencia de diversidad étnica, cultural y lingüística; y por una historia de movimientos poblacionales ligada a variaciones geopolíticas. El objetivo de este trabajo es presentar los primeros resultados de una investigación, estructurada en tres ejes (local, regional y sudamericano), que busca explorar la historia poblacional de la región a través de la caracterización de los linajes maternos y paternos de las ciudades de Corrientes (CRR, N=151) y Formosa (FOR, N=274). Se detallan las técnicas y métodos de análisis, así como las preguntas de investigación abordadas en cada eje, y se presentan los resultados del primer eje. Se estimaron los aportes de origen nativo americano, del oeste de Eurasia y de África subsahariana. Se obtuvieron frecuencias mayoritarias de linajes nativos americanos por vía materna (FOR=85%, CRR=89,4%). Los linajes euroasiáticos (FOR=9,93%, CRR=12,41%) y africanos (FOR=2,55%, CRR=0,66%) también están presentes. Por otra parte, más del 80% de los linajes paternos resultaron de origen euroasiático, de los cuales el 55% corresponde al haplogrupo R1 (FOR=55,2%, CRR=54,7%). La fracción nativa de los linajes paternos aportó las frecuencias más bajas documentadas para Argentina (FOR= 4,9%; CRR=3,2%). Los datos moleculares se compararon con datos genealógicos y se contextualizaron con la información histórica.
Palabras clave: Antropología molecular; ADN mitocondrial; Cromosoma Y; Nordeste argentino.
Abstract
Northeast Argentina is characterized by the presence of ethnic, cultural and linguistic diversity; and by a history of population movements linked to geopolitical variations. The aim of this paper is to present a research design, structured in three analytical axes (local, regional and South American), to explore the population history of the region through the characterization of maternal and paternal lineages of the cities of Corrientes (CRR N=151) and Formosa (FOR N=274). Analysis techniques and methods were detailed, the research questions addressed in each axis were outlined and the results of the first axis were presented. Genetic contributions of Native American, western Eurasia, and sub-Saharan Africa origin were estimated. Majority frequencies of Native American lineages were obtained through the maternal line (FOR=85%, CRR=89,4%). The Eurasian (FOR=9,93%, CRR=12,41%) and African (FOR=2,55%, CRR=0,66%) lineages were also present. On the other hand, more than 80% of the paternal lineages were of Eurasian origin, with 55% corresponding to R1 (FOR=55,2%, CRR=54,7%). The native American fraction (FOR= 4,9%; CRR=3,2%), provided the lowest frequencies documented for Argentina. Molecular data were compared with genealogical data and contextualized with historical evidence.
Keywords: Molecular Anthropology; Mitochondrial DNA; Y Chromosome; Northeastern Region of Argentina.
Recibido 22-03-2022
Recibido con correcciones 23-05-2022
Aceptado 08-06-2022
Introducción
Antecedentes
Las poblaciones argentinas contemporáneas, al igual que en el resto de América Latina, se configuraron como resultado de un proceso de poblamiento inicial y del posterior mestizaje entre nativos y migrantes desde tiempos coloniales. Particularmente, en Argentina se registra un significativo flujo migratorio europeo entre 1860 y 1930 y, a diferencia de otros países aledaños, se caracteriza por el menor aporte de población de origen africano (Studer, 1958).
Los estudios genéticos de linajes haploides de transmisión exclusivamente materna o paterna permiten identificar el origen continental remoto y han revelado un proceso de mestizaje de tipo sexo-asimétrico entre las poblaciones nativas preexistentes y los migrantes -en su mayoría europeos y varones- (para Argentina: Avena et al., 2007; García et al., 2018; Dipierri et al., 1998; Martínez Marignac et al., 1999; entre otros). La presencia de linajes maternos nativos puede ser muy significativa en poblaciones urbanas de Argentina, alcanzando en algunos casos del Noroeste Argentino (NOA) frecuencias mayores al 90% (ver Motti et al., 2013). Como contrapartida, la frecuencia de linajes paternos nativos en poblaciones cosmopolitas argentinas oscila alrededor del 10%, aumentando drásticamente hasta ~40% en el NOA (ver, entre otros, Di Fabio Rocca et al., 2016; Jurado Medina et al., 2014; Ramallo et al., 2009).
El avance en técnicas moleculares ha facilitado la obtención de secuencias mitocondriales completas. Ésto ha posibilitado la definición de clados de menor jerarquía dentro de los haplogrupos, muchos de los cuales se encuentran restringidos geográficamente, y dan cuenta de la historia biológica regional prehispánica, al hacer foco en el componente nativo americano (Brandini et al., 2018; Bodner et al., 2012; De Saint Pierre et al., 2012; Gómez-Carballa et al., 2018).
Este trabajo se enmarca en el plan de tesis doctoral “Dinámica de poblaciones humanas del NEA. Aportes desde el estudio de linajes uniparentales”, que se centra en reconstruir las historias de poblamiento de las áreas de influencia de las ciudades de Corrientes y Formosa en la región del Nordeste argentino (NEA) a partir de la identificación de linajes mitocondriales y del Cromosoma Y, y el análisis de su distribución a nivel local, regional y sudamericano. En este trabajo se presenta el diseño de la investigación, estructurado en tres ejes de análisis y los resultados correspondientes al primer eje, dedicado a analizar el origen continental de los linajes. El objetivo de este primer eje, consiste en caracterizar la dinámica poblacional post-hispánica, ya que se analiza el origen continental remoto de los linajes paternos y maternos, así como también los movimientos migratorios recientes a través de información genealógica. Los datos moleculares y genealógicos son discutidos en el marco de las evidencias históricas y demográficas, de manera de contribuir al conocimiento de la historia de las poblaciones locales.
Contextualización regional
El NEA comprende las actuales provincias de Corrientes, Chaco, Misiones y Formosa y comienza a cobrar entidad como región en la década de 1950, cuando se provincializan las tres últimas jurisdicciones (Valenzuela, 2014). Sin embargo, se observa que, históricamente, los movimientos poblacionales y el impacto cultural no necesariamente han respondido a factores exclusivamente geográficos, y mucho menos a límites políticos (Hernández, 1995). Esta división no coincide con una delimitación cultural, pues existe una íntima relación entre esta región con el Paraguay meridional, Brasil meridional y Uruguay (Lafon, 1971), como también una vinculación con el área amazónica, el norte de América del sur y la franja oriental del noroeste de nuestro país. Además, se debe tener en cuenta que, en algunos casos, los procesos migratorios aquí estudiados anteceden a la conformación del Estado argentino.
Desde una perspectiva geohistórica, Ernesto Maeder, en conjunto con otros autores, ha intentado rastrear la existencia de la región antes de que se definiera formalmente como tal. Particularmente, desde el punto de vista geográfico Bruniard y Bolsi (1992) han definido al NEA como un “variado mosaico de paisajes geográficos” debido a la asociación de diversas formas naturales y culturales, por lo que resulta difícil de asimilar a este territorio como una región homogénea o como una región nodal y funcional, teniendo en cuenta su génesis y su funcionamiento (Leoni, 2016). En este sentido, Maeder (2015) analiza la construcción de la historia regional del NEA y caracteriza los diferentes procesos que la conforman: las misiones jesuíticas de guaraníes; la formación del estado correntino, en donde se destaca el rol de Asunción, actual capital de Paraguay, como fundadora de distintas ciudades de la zona; y la ocupación y organización de los espacios antes marginales del Chaco Austral y Central, que incluían al territorio de Formosa. Teniendo en cuenta un origen en la época colonial y centrándose en el escenario geográfico de las misiones jesuíticas, Maeder (1977) reflexiona acerca de que la delimitación de la región no sólo debería incluir a las provincias argentinas sino también a Paraguay y al sur de Brasil. Sin embargo, pese a los distintos trabajos realizados por el autor, se señala la dificultad para pensar en una historia regional del NEA en conjunto, ya que esta región se configura -en palabras de Maeder- a partir de “procesos sincrónicos pero independientes” (Leoni, 2016).
Los antecedentes de estudios de ADN mitocondrial en la región pueden dividirse en dos grandes grupos: los que se han orientado al análisis de poblaciones aborígenes (Cabana et al., 2006; Sala et al., 2010; Sevini et al., 2013; Sala et al., 2019) y aquellos que analizan las fracciones del componente nativo en las poblaciones “cosmopolitas” o mestizas (Bobillo et al., 2010; Catelli et al., 2011; Sevini et al., 2013; Badano et al., 2018). Particularmente, entre estos últimos, se informan valores de linajes maternos nativos que van desde el 47% (Bobillo et al., 2010) a 68,6% (Badano et al., 2018) en la provincia de Misiones, hasta 80% en Corrientes y Formosa (Bobillo et al., 2010). Por otra parte, algunos trabajos indagan en la diversidad existente al interior del componente nativo americano. Por ejemplo, Demarchi (2014) encuentra proximidad genética entre los grupos lingüísticos mataco y guaycurú, indicando que no existe una asociación entre las diferencias lingüísticas y genéticas. Por el contrario, Sala et al. (2010) señalan mayor afinidad entre grupos guaraníes de Argentina y Brasil, respecto de otros grupos geográficamente más cercanos. Sevini et al. (2013) distinguen un componente andino y otro amazónico entre los criollos1 y los wichís del Chaco, respectivamente. Por último, Sala et al. (2019) corroboran la homogeneidad de los grupos Qom que habitan el Gran Chaco, y lo atribuyen, en parte, a que comparten un lenguaje común y a recientes eventos de migración.
Entre los trabajos que estudian linajes del cromosoma Y para la región del NEA, tanto Ramallo et al. (2009) como Jurado Medina et al. (2014), observan contribuciones de linajes nativos de al menos 80% en las poblaciones analizadas. Además, en estos trabajos los autores plantean que las poblaciones chaqueñas muestran gran diversificación por línea paterna sin evidencia de aislamiento reproductivo entre ellas, ya que linajes idénticos son compartidos entre distintas poblaciones, en concordancia con las conclusiones planteadas por Demarchi (2014) para la línea materna. En un trabajo más reciente, Paz Sepúlveda et al. (2020), analizando comunidades de Gran Chaco, concluyen que estas poblaciones no presentan diferencias genéticas significativas entre sí y que, cuando se comparan con poblaciones del NOA, no hay evidencia de aislamiento por distancia ni de barreras genéticas entre ellas, señalando la posible existencia de antiguas conexiones entre las dos regiones. Por último, en Sevini et al. (2013) se obtuvieron frecuencias de haplogrupos paternos nativos de 80% en los wichís y 20% en los criollos analizados.
En resumen, se puede vislumbrar que el abordaje de las historias de poblamiento de la región del NEA no es una tarea sencilla, debido a que esta región incluye distintas áreas con características prehispánicas distintivas, tales como la región chaqueña y la del litoral, sumado a que la historia de esta región se ha visto fuertemente influenciada por las dinámicas comprendidas en los países limítrofes, tanto en momentos prehispánicos como en tiempos históricos (Maeder, 2015).
Finalmente, se observa que gran parte de los trabajos que analizan secuencias mitocondriales y del cromosoma Y del NEA se basan en muestreos de grupos indígenas. Por lo tanto, el análisis de nuevas secuencias de poblaciones cosmopolitas de la región realizará una contribución importante al estudio del NEA, que podría derivar en una interpretación innovadora de los procesos de poblamiento.
Metodología
Esquema de trabajo
Para abordar las múltiples dimensiones que conforman la historia poblacional del NEA, se implementó un diseño metodológico que estructura el análisis de la diversidad genética en tres ejes con distintos niveles de resolución. El primer eje se centra en la historia local de las migraciones intercontinentales, basándose en el análisis de las frecuencias de haplogrupos de distintos orígenes continentales (nativo americano, de Europa y Oriente Medio, y africano), tanto de los linajes maternos como paternos, en las localidades de Formosa y Corrientes. Se partió de la hipótesis general de que en las poblaciones cosmopolitas actuales es posible rastrear linajes de poblaciones ya desaparecidas, a pesar de los procesos de mestizaje que han ocurrido durante los últimos 500 años. Esto último ha sido corroborado para otras regiones de Argentina (García et al., 2018; Motti et al., 2012; Schwab, 2019; entre otros). A su vez, los resultados acerca del origen continental de los linajes paternos y maternos fueron contrastados con la información genealógica proporcionada por los individuos a través de una encuesta.
Un segundo eje de análisis hace foco en la fracción nativa americana de los haplogrupos, mediante el análisis de haplotipos del ADNmt y del cromosoma Y. Este enfoque a un mayor nivel de resolución posibilita identificar subhaplogrupos diferenciados regionalmente que permiten una aproximación a la historia regional. Para ello, se propone la obtención de secuencias de la región control completa del ADNmt de muestras poblacionales de la localidad de Formosa y de Corrientes y se analizarán secuencias previamente recolectadas por nuestro grupo de investigación, como también secuencias tomadas de la bibliografía. También se realizarán análisis de microsatélites del cromosoma Y y se recopilará información publicada. El objetivo de este segundo nivel de análisis consiste en establecer las afinidades de las poblaciones estudiadas en el contexto del Cono Sur. El abordaje de las poblaciones desde un análisis molecular nos permitirá establecer las relaciones entre los haplotipos mitocondriales y del cromosoma Y de distintas localidades y regiones, aumentando nuestra capacidad para interpretar la diferenciación poblacional hacia el interior del continente y, de esta forma, discutir modelos de poblamiento a escala regional (Motti et al., 2017). Teniendo en cuenta la estructura geográfica observable para el ADNmt y el cromosoma Y, se espera que existan similitudes en la distribución de clados y haplotipos dentro de la región del NEA y con los países colindantes, en íntima relación con la región (Paraguay, sur de Brasil y Uruguay), así como diferencias con otras regiones de la Argentina.
Finalmente, en el tercer y último eje, se definirán y analizarán detalladamente nuevos clados mitocondriales caracterizados a partir de la combinación de polimorfismos de las regiones control y/o codificante. En base a una compilación de mitogenomas publicados e inéditos, se seleccionarán PNS informativos para los que se diseñarán ensayos de polimorfismo en la longitud de los fragmentos amplificados (APLP, por sus siglas en inglés), con el objetivo de identificar dichos clados en distintas localidades de la Argentina. La información molecular colectada se utilizará para postular presuntos clados monofiléticos de diferente jerarquía, explorando su distribución y estructuración a escala sudamericana o continental, con miras a identificar las principales rutas de poblamiento seguidas por las poblaciones que se asentaron en el NEA.
Muestreos poblacionales
Se realizaron viajes de campaña durante agosto de 2015 a las ciudades capitales de Formosa y Corrientes. Estas ciudades fueron seleccionadas debido a que atraen migrantes de otros lugares de dichas provincias. Los muestreos se realizaron en los Bancos de Sangre Central de ambas localidades y en un laboratorio privado en la ciudad de Formosa. Se invitó a participar en este estudio a todas aquellas personas que concurrieran para extracción de sangre, ya sea para donación o análisis clínicos. Se incluyeron en el estudio personas mayores de 18 años (inclusive), a las cuales se realizó una encuesta genealógica en la que se recolectó información sobre el lugar de nacimiento de los padres y abuelos por línea materna y paterna (ver modelo de encuesta genealógica en Anexo 1 de Material Suplementario). Las muestras de sangre fueron tomadas por personal de la institución capacitado para dicha tarea.
De las 506 muestras de sangre colectadas, se seleccionaron 425 pertenecientes a las 3 estaciones de muestreo: 274 de Formosa y 151 de Corrientes, de las cuales 142 y 95 fueron varones, respectivamente. Sólo se incluyeron individuos cuyos linajes maternos y/o paternos no compartieran ancestros genealógicos conocidos y que hubieran concurrido espontáneamente a la institución. Las muestras de los dos puntos de colecta de Formosa se unificaron a los efectos de este trabajo, al no existir diferencias significativas entre ellas en cuanto al origen continental de sus haplogrupos mitocondriales, ni de las frecuencias de haplogrupos del cromosoma Y (Test Exacto de Diferenciación Poblacional: P > 0,05 para todas las comparaciones).
Procesamiento de muestras biológicas
Se separaron los linfocitos del resto de los componentes sanguíneos en el sitio de muestreo. Para ello, 5-10 ml de sangre total fue centrifugada, lavada con solución de lisis de hematíes (1mM NH4 HCO3, 115mM NH4 Cl), resuspendida y conservada en solución de lisis de linfocitos (100mM Tris-HCl pH 7,6 40mM EDTA pH 8,0, 50mM NaCl, 0,2% SDS, 0,8mM azida sódica). Luego, se completó la extracción de ADN con 0,5 volúmenes de NaCl 5,4M para lograr la precipitación de proteínas, seguida por la precipitación de ácidos nucleicos con 1 volumen de alcohol isopropílico.
Consideraciones éticas y devolución de resultados a la población
Los donantes fueron previamente informados y expresaron su voluntad de participar a través de la firma de un formulario de consentimiento informado. Las muestras biológicas fueron codificadas para resguardar la confidencialidad de los participantes durante su procesamiento en el laboratorio. Este trabajo se encuentra dentro de un proyecto mayor de investigación que fue aprobado por el Comité Provincial de Bioética de la Provincia de Jujuy y por el Comité de Ética en Investigaciones Biomédicas del IMBICE.
Cada participante recibió sus resultados personales en sobre cerrado, informando sobre el haplogrupo y origen continental por línea materna (y paterna en el caso de los varones). Por otra parte, se informaron los resultados para la población general de la localidad muestreada a través de un afiche ilustrativo a colocarse en la institución en donde se realizó la recolección de las muestras. En ningún caso se obtuvieron o informaron resultados con posibles implicancias médicas para los sujetos participantes.
Análisis de Censos Nacionales
Pese a la existencia de Censos nacionales anteriores (de los años 1869 y 1895), se analizaron solo los de 1914 y 2010 a fin de determinar el origen de los migrantes en las provincias estudiadas (Tercer Censo Nacional, 1914; INDEC, 2010). La elección se realizó teniendo en cuenta que el territorio de Chaco y Formosa fue ocupado militarmente más recientemente, entre 1872-1884, luego de la guerra de la Triple Alianza.
Tipificación de haplogrupos uniparentales
Para el diagnóstico de los haplogrupos uniparentales se utilizó la técnica PCR-APLP multiplex y se siguió la metodología utilizada por Motti et al. (2009) para el ADN mitocondrial y de Jurado Medina (2015) para el cromosoma Y. Estos autores se basaron en los trabajos de Umetsu y colaboradores, quienes desarrollaron una técnica de tipificación alelo-especifica de SNPs para ADN mitocondrial (Umetsu y col., 2001; Umetsu y col., 2005). Se detallan a continuación las secuencias de los cebadores utilizados para la asignación de origen continental por línea materna (Tabla 1). Los cebadores utilizados para línea paterna se encuentran publicados en Jurado Medina et al. (2014).
Tabla 1. Cebadores utilizados para asignar el origen continental por línea materna.
Table 1. Primers used to assign continental origin by maternal line.
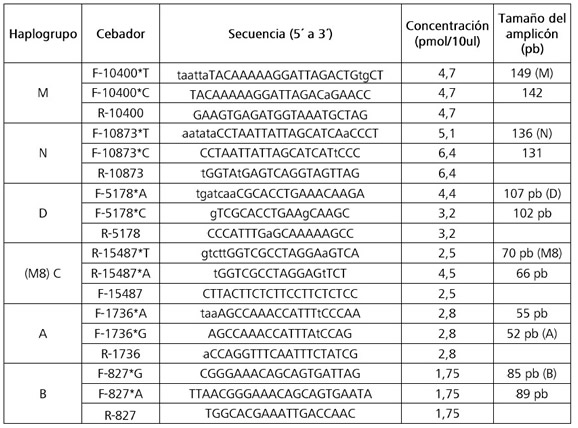
La asignación del origen continental de los linajes maternos y paternos se realizó teniendo en cuenta la historia migratoria y poblacional de Argentina. Los haplogrupos mitocondriales A-D fueron considerados de origen nativo americano y el parahaplogrupo N(xA,B), como proveniente de Europa o Medio Oriente. Las muestras que no pudieron ser asignadas a los súper-haplogrupos M o N fueron secuenciadas en su región control completa (datos no mostrados) y confirmadas como pertenecientes al parahaplogrupo L(xM,N), de origen africano. Los linajes paternos fueron considerados de origen nativo americano si pertenecían al clado Q (M3) y de origen alóctono si no lo eran. Las comparaciones entre frecuencias de haplogrupos y entre orígenes continentales se realizaron a partir del Test Exacto de diferenciación poblacional con el programa Arlequín versión 3.5 (Excoffier et. al., 2010).
Procesamiento de información genealógica
De la información genealógica brindada por los donantes, se recopilaron los datos sobre el último ancestro conocido (UAC) por vía materna y/o paterna. Para aquellos encuestados en los que este dato no fuera conocido o certero, se consideró el lugar de nacimiento de la generación parental (madre o padre) o del mismo donante. Se trabajó con dos categorías de procedencia: “regional” y “extra regional”. Se consideró que el lugar de origen era regional cuando el UAC tuvo como lugar de nacimiento a las provincias que conforman la región del NEA (Corrientes, Formosa, Misiones y Chaco) o los países limítrofes al NEA (Paraguay, Uruguay y Brasil). A las localidades informadas de otras provincias de la Argentina, de otros países de Sudamérica o de otros países europeos, se las consideró dentro de la categoría extra regional.
A partir del origen declarado en la encuesta genealógica, se detectó que ciertos linajes no eran locales del sitio de muestreo. Sin embargo, tomamos la precaución de no eliminar ni redistribuir las muestras en concordancia al origen declarado, de forma tal de no interferir en la distribución actual de los linajes maternos y paternos, siguiendo el mismo criterio que en Motti et. al. (2013). Una vez obtenidos los datos moleculares y genealógicos del UAC de cada donante, se compararon los resultados. Sólo se pudo comparar la proporción de ancestros por vía materna que tuvieron un origen ultramarino con aquellos que pertenecen al superhaplogrupo N. Para la vía paterna, sólo se compararon los ancestros con un origen local declarado (Argentina + países sudamericanos), con aquellos que pertenecen al clado Q-M3, mientras que el resto de las categorías utilizadas se agrupó como “ultramar”. Las demás categorías no pudieron ser homologadas con datos moleculares, y en ningún caso hubo ancestros declarados de origen africano.
Resultados
Los resultados de este trabajo comprenden el primer eje de análisis, descripto previamente, en el que se trabajó con frecuencias de haplogrupos de linajes maternos y paternos en las localidades de Formosa y Corrientes. A partir de la tipificación de mutaciones diagnósticas del ADNmt y del Cromosoma Y, se determinaron los orígenes continentales de las poblaciones (procedencia de América, Europa o Medio Oriente o África subshariana). Además, se contrastaron los resultados moleculares con información genealógica.
Linajes maternos
Frecuencias de haplogrupos mitocondriales
Los resultados demostraron una presencia mayoritaria de linajes nativos americanos por vía materna a nivel global (86,59%) y en cada una de las localidades de muestreo (FOR= 85%, CRR= 89,4%). Por otra parte, el aporte de linajes europeos y de Medio Oriente fue variable entre ellas (FOR=9,93%, CRR=12,41%). Es interesante destacar la relativamente alta proporción de linajes africanos en la muestra FOR=2,55%, siendo menor en CRR=0,66%. Sin embargo, un Test Exacto de Diferenciación Poblacional (TEDP) (Raymond & Rousset, 1995) muestra que no existen diferencias significativas en cuanto a los aportes relativos continentales entre ambas poblaciones (TEDP: P > 0,05 para todas las comparaciones).
Información genealógica para la línea materna
Respecto a la información genealógica brindada por los donantes, se recopiló el dato sobre el UAC por vía materna, siendo en un 74% de los casos de la tercera generación (FOR= 73,4%; CRR=74,8). Aproximadamente un 20% de la información provino de la madre (FOR= 23,7%; CRR=21,2%) y el resto del donante (FOR= 2,9%; CRR=4%).
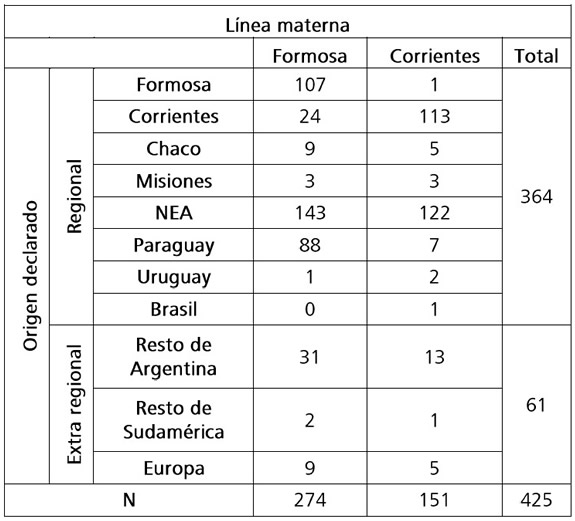
Los resultados demostraron un mayor componente “regional” (85,6%) -en el corto plazo- del origen declarado por los donantes por línea materna (ver Tabla 2). El 62,4% declara que sus ancestros nacieron en la región del NEA y un 22,4% en Paraguay. De aquellos que declararon un origen extra-regional, 10,4% fue del resto de Argentina, 0,7% del resto de Sudamérica y 3,3% de origen europeo (Tabla 2). Se destaca que el 74,8% de los donantes de Corrientes declara que sus ancestros nacieron dentro de la misma provincia y que se registra un aporte de 32,1% de ancestros de origen paraguayo en Formosa (Tabla 2).
Tabla 2. Lugar de origen declarado de último ancestro conocido por vía materna, en porcentaje por localidad. NEA = Nordeste Argentino.
Table 2. Declared place of origin of the last known maternal ancestor, in percentage by locality. NEA = Northeast region.
Comparación entre resultados moleculares y genealógicos para la línea materna
Los resultados de esta sección se analizaron en conjunto, debido a que no hubo diferencias significativas entre las localidades estudiadas (datos no mostrados).
Cuando se realizaron las comparaciones de los orígenes identificados genéticamente con aquellos declarados por los donantes, se observó que 14 de los 425 donantes analizados para la línea materna declaran un origen ultramarino, de los cuales 11 (79%) fueron corroborados por el resultado molecular, mientras que los tres restantes resultaron portadores de linajes nativos. Estas discrepancias en las comparaciones por vía materna podrían explicarse por pérdida de este dato en la historia familiar, casos de adopción, apropiación ilegítima, entre otros. En aquellos que declaran un origen local de sus ancestros (N=411), ~89% (N=365) presenta ancestría nativa americana. Sin embargo, un 11% (N=46) presenta un origen alóctono, ya que los resultados moleculares arrojan que el 9% es de origen europeo y el 2% de origen africano.
Linajes paternos
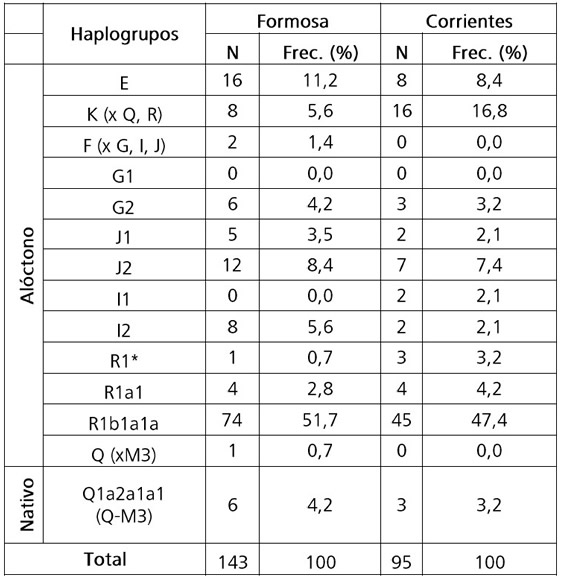
Frecuencias de haplogrupos del Cromosoma Y
Más del 80% de los linajes paternos en las poblaciones muestreadas correspondieron a un origen ancestral europeo y/o de Medio Oriente, con valores de alrededor de 55% para R1 (haplogrupo R1: FOR= 55,2%, CRR= 54,7%) (Tabla 3). Cabe destacar la baja proporción de linajes nativos Q-M3 (FOR: 4,9%; CRR: 3,2%) en relación a la fracción alóctona (Tabla 3). Otro dato interesante es que el 100% de los donantes que poseen un linaje nativo por línea paterna también resultaron nativos por línea materna.
Tabla 3. Frecuencia de los haplogrupos del cromosoma Y por localidad de muestreo, en porcentaje.
Table 3. Frequency of Y chromosome haplogroups by sampling locality, in percentage.
No se encontraron diferencias significativas entre ambas poblaciones en cuanto a los aportes relativos de los clados (TEDP: P > 0,05).
Información genealógica para la línea paterna
Los datos de los donantes que informaron acerca de su UAC por línea paterna fueron en un 55,3% de los casos de la tercera generación a nivel global (FOR=59,2%; CRR=49,5%), registrándose, además, una frecuencia mayor al 30% de datos que provenían de la segunda generación (FOR=34,5%; CRR=42,1%). El resto de la información provino de los donantes (FOR=6,3%; CRR=8,4%).
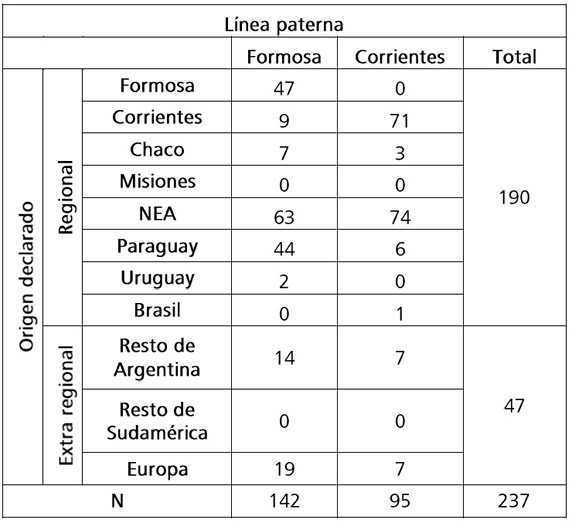
Se observa un mayor componente “regional” ( ~80%) en el corto plazo, exhibido a través de la historia oral (Tabla 4), preponderantemente en aquellos cuyo lugar de nacimiento corresponde al NEA y especialmente para la localidad de Corrientes. Se registra un aporte del 31% de ancestros de origen paraguayo en Formosa.
Tabla 4. Lugar de origen declarado de último ancestro conocido por vía paterna, en porcentaje por localidad. NEA = Nordeste Argentino.
Table 4. Declared place of origin of the last known paternal ancestor, in percentage by locality. NEA = Northeast region.
Comparación entre resultados moleculares y genealógicos para la línea paterna
Se obtuvieron los datos genealógicos para la línea paterna y se analizaron en contraste con los resultados moleculares obtenidos, en los casos que eran comparables.
Los resultados demuestran una mayor frecuencia de donantes varones que declaran un origen local de sus ancestros paternos más remotos (210/236), en comparación con aquellos que declaran origen ultramarino (26/236). En ese sentido, los resultados moleculares indican que 25 de los 26 que declaran un origen ultramarino efectivamente son portadores de linajes euroasiáticos, mientras que el restante lleva uno de origen nativo americano. Por el contrario, de aquellos que mencionan un origen local de sus ancestros, sólo 4% de los resultados moleculares coinciden con la información declarada, mientras que ~96% (N= 201) tiene un origen euroasiático.
Por último, se compilaron los resultados de frecuencias de linajes nativos para la línea materna y paterna de este trabajo con los de 15 localidades de Argentina del NOA y COA (Motti, 2012; Jurado Medina, 2015; Schwab, 2019; Ramallo et al., 2009) analizados previamente (Figura 1).
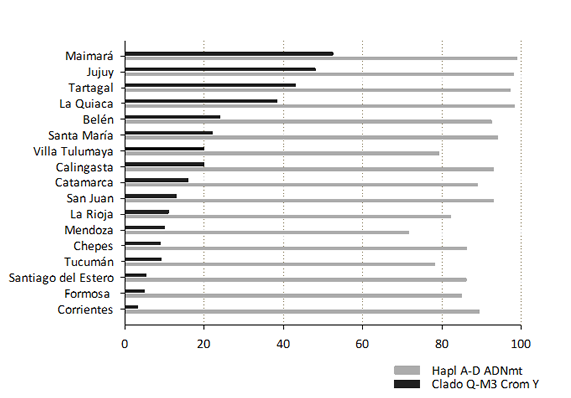
Figura 1. Frecuencia relativa de linajes nativos americanos por línea materna (haplogrupos A-D) y por línea paterna (clado Q-M3) en Formosa y Corrientes, y en las localidades muestreadas hasta el momento en la colección del IMBICE, en porcentaje. Hapl= haplogrupo; Crom Y= Cromosoma Y.
Figure 1. Relative frequency of native american lineages by maternal line (A-D haplogrups) and by paternal line (Q-M3 clade) in Formosa and Corrientes and in sampling localities until now in IMBICE collection, in percentage. Hapl= haplogroup; Crom Y= Y chromosome.
Discusión
Migraciones intercontinentales
Una fracción importante de las personas que contribuyeron al acervo génico actual de Argentina arribó durante las migraciones ultramarinas entre 1860-1930. El trabajo de Dipierri et al. (1998) fue pionero en demostrar un proceso de mestizaje entre las poblaciones preexistentes y los migrantes -en su mayoría europeos y varones-, con aportes diferenciales según el sexo, utilizando tanto marcadores mitocondriales como del cromosoma Y. De esta forma, se observa que, mayoritariamente, los linajes maternos presentan ancestría nativa y los linajes paternos realizan un aporte alóctono. En este sentido, nuestros resultados se asemejan a los resultados de estudios previos y a los realizados en nuestro laboratorio en 15 localidades de la Argentina (Motti, 2012; Jurado Medina, 2015; Schwab, 2019). Particularmente, se observa que la frecuencia de linajes nativos maternos en Corrientes y Formosa es mayor al 85%, al igual que en la mayoría de las localidades del NOA y centro-oeste de Argentina analizadas previamente por nuestro grupo (Figura 1). Este predominio de linajes de origen nativo se relaciona con un menor ingreso de inmigrantes ultramarinos en la zona norte del país, tanto para el NEA como para el NOA, a diferencia de un mayor flujo de migrantes de ultramar en regiones como la pampeana y la patagónica, en donde se reportan frecuencias del componente nativo del 50% para la provincia de Buenos Aires y de alrededor del 65% para las provincias patagónicas (Avena et al., 2009; Bobillo et al., 2010; Parolín et al., 2013).
Contrariamente, para los linajes paternos se destaca la muy baja frecuencia del haplogrupo nativo, particularmente en las muestras de Corrientes (3,2%) y Formosa (4,9%). Además, la representación del aporte que realiza el clado Q-M3 al acervo génico de distintas localidades de Argentina permite vislumbrar que las frecuencias obtenidas para las provincias del NEA son, junto a las de Santiago del Estero (5,3%), las más bajas informadas hasta el momento para Argentina (Figura 1). Para contextualizar estos resultados, es necesario tener en cuenta el escenario histórico de la Guerra de la Triple Alianza, reconocida como una de las contiendas más largas y sangrientas de la historia latinoamericana, en donde se enfrentaron Argentina, Uruguay y Brasil contra el Paraguay desde 1864 a 1870. Baratta (2013) realiza un análisis de la producción historiográfica sobre la guerra en donde destaca que, pese a la activa participación de la Argentina en la contienda, se ha desdibujado su registro de la historia, a pesar del impacto demográfico de la misma debido a la gran cantidad de bajas y su significación clave en la consolidación del Estado Argentino. Esto se ha puesto de manifiesto en trabajos realizados en las últimas décadas con nuevas perspectivas. En particular, se ha dado relevancia al papel de Corrientes durante la guerra del Paraguay en la historiografía argentina en forma reciente. Por ejemplo, Ramírez Braschi (2004), realizando un análisis de la prensa en esta provincia, coloca a Corrientes como escenario geográfico del campo de batalla durante los primeros años de la guerra, y luego como retaguardia y campo de aprovisionamiento de los ejércitos aliados, con lo cual la dinámica de la ciudad se vio alterada radicalmente hasta 1869. En este contexto, es altamente probable que haya ocurrido una disminución drástica de varones, con la consecuente pérdida de linajes nativos americanos, tal como se observa a través de los análisis moleculares. Por otra parte, teniendo en cuenta que Formosa es una ciudad atractora de migrantes oriundos del Paraguay, lo cual pudo vislumbrarse cuando se analizaron los censos nacionales de 1914 y de 2010, y teniendo en cuenta el “Holocausto masculino” vivido durante la Guerra de la Triple Alianza por el pueblo paraguayo (Baratta, 2013), consideramos que este mismo escenario histórico permite explicar la baja frecuencia de linajes paternos nativos en ambas localidades.
Historia oral
Respecto a los resultados obtenidos a partir del análisis de la historia oral de los donantes, se observa una mayor frecuencia de información genealógica proveniente de la abuela materna que del abuelo paterno (74% y 56% respectivamente), indicando una mayor profundidad temporal en la historia oral registrada para la línea materna. A su vez, se destaca que predomina un origen regional del ancestro más remoto declarado (materno: 85,6%; paterno: 80,2%) y se verifica una fuerte influencia de migrantes provenientes de Paraguay en la ciudad de Formosa, tendencia que ya podía observarse en los datos reportados en el Censo Nacional de 1914. Este fenómeno es especialmente notable en Formosa donde, sobre una población de 8.774 habitantes, el 45% era extranjero y, a su vez, de esta fracción el 84% provenía de Paraguay y 13,6% era de origen ultramarino; mientras que, para Corrientes, sobre una población de 24.462 habitantes, el 7% era extranjero, 16,9% era de origen paraguayo y el 39% era de origen ultramarino. Por lo tanto, la frecuencia obtenida nos indica la influencia de Paraguay en la región del NEA respecto a los procesos poblacionales y, contextualizada con los datos censales, permite inferir una continuidad histórica de esta influencia hasta la actualidad.
Además, la comparación de datos moleculares y genealógicos permite dar cuenta de que una mayor parte de los donantes declara un origen local de sus ancestros paternos y maternos más remotos en las localidades estudiadas. En particular, para los resultados obtenidos de la línea materna de aquellos que declararon un origen local del ancestro más remoto (en 89% se verificó ancestría nativa americana), se considera que esta imagen podría ser el reflejo de las poblaciones criollas de la región en tiempos coloniales. Resultados semejantes se han obtenido en nuestro laboratorio por Motti (2012) en 1734 muestras de la Región del NOA y Centro Oeste argentino.
Por otra parte, para la línea paterna se destaca un componente mayoritario que declara un origen local (210/236) pero que, sin embargo, en aproximadamente el 96% de los casos se identificó un origen molecular euroasiático. Estos resultados reflejan la antigüedad de los eventos de mestizaje, que probablemente se remontan a las primeras etapas de colonización del territorio, razón por la cual no ha quedado registrado en la historia oral. Por otra parte, dan cuenta de la asimetría del mestizaje en la población criolla. Además, se observa una mayor pérdida de la historia paterna por paternidades no conocidas. En línea con estos resultados, en un trabajo realizado por Muzzio et al. (2012), en el que se evalúa la proporción de transmisión de apellidos por línea paterna en diferentes regiones de Argentina, se demostró una alta proporción de paternidades no reconocidas entre la población general, una muy rápida pérdida de asociación entre los linajes paternos y los apellidos, y significativas diferencias regionales.
Las discrepancias entre los resultados obtenidos de la historia oral y de análisis moleculares, tal como plantea Sans (2009), posiblemente reflejen los procesos de construcción de la identidad nacional, que explícitamente acepta el exterminio indígena, por una integración de los indígenas a la sociedad luego de las guerras, por las uniones entre mujeres indígenas y varones europeos, que habrían ocultado o perdido la identificación de su origen.
Por último, se debe tener en cuenta que la determinación de la ancestría a través de la historia oral –particularmente en este trabajo, del origen del ancestro más remoto- está limitada por el desconocimiento de las personas acerca de sus ancestros, aún de los recientes, sea por adopción u otros factores (Race, Ethnicity, and Genetics Working Group, 2005).
Conclusiones
Las investigaciones que analizan la variabilidad genética de las poblaciones argentinas se han incrementado en los últimos años, generando una importante cantidad de datos moleculares. Sin embargo, los intentos por comprender el contexto y las consecuencias sociales del conocimiento genético son asimétricamente menores. En este sentido, se observa que la investigación bioantropológica opera como “espacio-tiempo de resignificación, interviniendo activamente en la construcción de esas memorias e identidades” (García et al., 2016). El diseño metodológico planteado en este trabajo y la investigación que se deriva del mismo, contribuye a incrementar el conocimiento de la historia local, a través de la aplicación de técnicas que aportan una mirada independiente e innovadora y cuyos resultados permiten sumar un nuevo abordaje a las interpretaciones sobre los procesos que intervinieron en la conformación de las poblaciones del NEA.
Las nuevas perspectivas emergentes de interpretación histórica, bioarqueológica y demográfica, permitirán dar un marco de análisis a los datos genéticos, a fin de poder reconstruir las historias fracturadas y suprimidas y dar cuenta del complejo entramado cultural que habita cada región.
La Plata, 8 de Marzo de 2022
1. Los autores del citado trabajo hacen referencia al término “criollo” como la población mestiza que surge como resultado de los matrimonios entre las mujeres de poblaciones aborígenes y los primeros colonizadores varones europeos que arribaron al territorio.
Agradecimientos
A los voluntarios que donaron su muestra para esta investigación. A los directivos y al personal de los hospitales en donde realizamos los muestreos por su colaboración. Este trabajo fue financiado a través de los proyectos FONCyT: PICT 2013 N°1611, PICT 2015 N° 2167, y del PICT 2017 Nº 27 y el proyecto de la UNLP N° 836.
1. Avena S., Goicoechea A., Bartomioli M., Fernández V., Cabrera A., Dugoujon J.M., Dejean C., Fabrykant G., Carnese F.R. (2007). Mestizaje en el sur de la región pampeana (Argentina). Su estimación mediante el análisis de marcadores proteicos y moleculares uniparentales. Revista Argentina de Antropologia Biologica, 9(2), 56–76. Recuperado a partir de https://revistas.unlp.edu.ar/raab/article/view/383.
2. Avena S.A., Parolin M.L., Dejean C.B, Ríos Part M.C., Fabrikant G., Goicoechea A.S., Dugoujon J.M. y Carnese F.R. (2009) Mezcla génica de linajes uniparentales en Comodoro Rivadavia (Provincia de Chubut, Argentina). Revista Argentina de Antropología Biológica, 11(1), 25-41. Recuperado a partir de https://revistas.unlp.edu.ar/raab/article/view/261. [ Links ]
3. Badano, I., Sanabria, D. J., Totaro, M. E., Rubinstein, S., Gili, J. A., Liotta, D. J., ... y Schurr, T. G. (2018). Mitochondrial DNA ancestry, HPV infection and the risk of cervical cancer in a multiethnic population of northeastern Argentina. PloS One, 13(1), e0190966. https://doi.org/10.1371/journal.pone.0190966. [ Links ]
4. Baratta M. V. (2013). La Guerra del Paraguay y la historiografía argentina. História da Historiografia: International Journal of Theory and History of Historiography, Ouro Preto, 7(14), 98–115. DOI: 10.15848/hh.v0i14.614. Disponible en línea: https://www.historiadahistoriografia.com.br/revista/article/view/614.
5. Bobillo M.C., Zimmermann B., Sala A., Huber G., Röck A., Bandelt H.J., Corach D., Parson W. (2010) Amerindian mitochondrial DNA haplogroups predominate in the population of Argentina: towards a first nationwide forensic mitochondrial DNA sequence database. International Journal of Legal Medicine 124(4), 263-8. doi: 10.1007/s00414-009-0366-3 [ Links ]
6. Bodner, M., Perego, U. A., Huber, G., Fendt, L., Röck, A. W., Zimmermann, B., ... y Bobillo, M. C. (2012). Rapid coastal spread of First Americans: Novel insights from South America’s Southern Cone mitochondrial genomes. Genome Research, 22(5), 811–820. https://doi.org/10.1101/gr.131722.111
7. Brandini S., Bergamaschi P., Cerna M.F., Gandini F., Bastaroli F., Bertolini E., Cereda C., Ferretti L., Gómez-Carballa A., Battaglia V., Salas A., Semino O., Achilli A., Olivieri A., Torroni A. (2018) The Paleo-Indian Entry into South America According to Mitogenomes. Molecular biology and evolution, 35(2):299-311. doi: 10.1093/molbev/msx267 [ Links ]
8. Bruniard, E. y Bolsi, A. (1992). “Region agro-silvo-ganadera con frentes pioneros de ocupacion del Nordeste”. En: Roccatagliata, J (coord.) La Argentina. Geografía general y los marcos regionales. Buenos Aires, Planeta.
9. Cabana G., Merriwether D.A., Hunley K., Demarchi D.A (2006) Is the Genetic Structure of Gran Chaco Populations Unique? Interregional Perspectives on Native South American Mitochondrial DNA Variation. American Journal of Physical Anthropology, 131:108-119. DOI: 10.1002/ajpa.20410. [ Links ]
10. Catelli M.L., Alvarez-Iglesias V., Gómez-Carballa A., Mosquera-Miguel A., Romanini C., Borosky A., Amigo J., Carracedo A., Vullo C., Salas A. (2011). The impact of modern migrations on present-day multi-ethnic Argentina as recorded on the mitochondrial DNA genome. BMC Genetics, 12(1), 77. https://doi.org/10.1186/1471-2156-12-77 [ Links ]
11. de Saint Pierre M., Bravi C.M., Motti J.M.B., Fuku N., Tanaka M., Llop E., Bonatto S.L., Moraga M. (2012) An alternative model for the early peopling of southern South America revealed by analyses of three mitochondrial DNA haplogroups PLoS ONE7(9): e43486. https://doi.org/10.1371/journal.pone.0043486. [ Links ]
12. Demarchi D. A. (2014). Análisis de la estructura genética en poblaciones nativas del Gran Chaco. Folia Histórica del Nordeste, 22, 169-185. DOI: http://dx.doi.org/10.30972/fhn.02255 [ Links ]
13. Di Fabio Rocca F., Albeza M. V. B., Postillone M., Acreche N., Lafage L., Parolín M.L., Dejean C., Carnese F.R., Avena, S. (2016). Historia poblacional y análisis antropogenético de la ciudad de Salta. Andes, vol. 27, Instituto de Investigaciones en Ciencias Sociales y Humanidades Salta, Argentina. [ Links ]
14. Dipierri J., Alfaro E., Martínez Marignac V., Bailliet G., Bravi C., Cejas S., Bianchi N. (1998). Paternal Directional Mating in Two Amerindian Subpopulations Located at Different Altitudes in Northwestern Argentina. Human Biology, 70(6), 1001-1010. [ Links ]
15. Excoffier, L. & Lischer, H.E. (2010) “Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows”, Molecular Ecology Resources, 10, 564-567. https://doi.org/10.1111/j.1755-0998.2010.02847.x.
16. García A., Oliveira Rufino R., Bergese A.B., Agüero J.F., Cuevas A., Díaz-Rousseau G., Pauro M., Nores R., Garita-Onandía Y., Tavella M.P., Demarchi D.A. (2016). El cruce entre las antropologías. Una mirada interdisciplinaria en torno a la genética de poblaciones, las memorias familiares y la construcción identitaria. Revista del Museo de Antropología, 9(2), 105-112. DOI: https://doi.org/10.31048/1852.4826.v9.n2.13614 [ Links ]
17. García, A., Pauro, M., Bailliet, G., Bravi, C. M., & Demarchi, D. A. (2018). Genetic variation in populations from central Argentina based on mitochondrial and Y chromosome DNA evidence. Journal of human genetics, 63(4), 493-507. https://doi.org/10.1038/s10038-017-0406-7. [ Links ]
18. Golden Software. (2012). Surfer. Quick Start Guide. Golden, Colorado, E.E.U.U. Recuperado de www.GoldenSoftware.com [ Links ]
19. Gómez-Carballa A., Pardo-Seco J., Brandini S., Achilli A., Perego U.A., Coble M.D, Diegoli T.M., Álvarez-Iglesias V, Martinón-Torres F., Olivieri A., Torroni A, Salas A. (2018). The peopling of South America and the trans-Andean gene flow of the first settlers. Genome Research. (6):767-779. doi: 10.1101/gr.234674.118. [ Links ]
20. Hernández, I. (1995). Los indios de Argentina. Colección pueblos y lenguas indígenas 4. Madrid, Ed MAPFRE, 335p. [ Links ]
21. INDEC. (2010). Censo Nacional de Población, Hogares y Viviendas. [ Links ]
22. Jurado Medina L.S., Muzzio M., Schwab, M.E., Bravi C., Costantino M.L, Barreto G. Guillermo, Bailliet, G. (2014). Human Y-chromosome SNP characterization by multiplex amplified product-length polymorphism analysis. Electrophoresis, 35(17), 2524-2527. DOI: https://doi.org/10.1002/elps.201400020.
23. Jurado Medina L.S., Ramallo V., Calandra H., Lamenza G., Braunstein J., Salceda S., Bailliet G. (2014). Linajes paternos del Gran Chaco, un abordaje desde el ADN. Folia Histórica del Nordeste (22): 187-202. DOI: http://dx.doi.org/10.30972/fhn.02256. [ Links ]
24. Jurado Medina, L. S. (2015). Tipificación de marcadores uniparentales en poblaciones mestizas de Argentina, Tesis Doctoral Inédita, La Plata, Universidad Nacional de La Plata. [ Links ]
25. Lafon, C. 1971. Introducción a la arqueología del Nordeste argentino. Relaciones V (2): 119-152. Buenos Aires. [ Links ]
26. Leoni M. S. (2016) La región NEA en la perspectiva de Ernesto Maeder. XXXVI Encuentro de Geohistoria regional. ISBN: 978-987-4450-00-5 [ Links ]
27. Maeder E.J.A. (2015). Los grupos subalternos en el nordeste del Virreinato del Río de la Plata. Rosario, Prohistoria Ediciones. [ Links ]
28. Maeder, Ernesto J.A (1977). “Breve historia del Nordeste Argentino en su relacion con el Paraguay y Rio Grande do Sul. En: Revista de Estudios Regionales, Año 1, N° 2, Corrientes.
29. Martínez-Marignac V., Bravi C., Lahitte H., Bianchi N. (1999). Estudio del ADN mitocodrial de una muestra de la ciudad de La Plata. Revista Argentina De Antropología Biológica, 2(1):281-300. http://sedici.unlp.edu.ar/handle/10915/6003. [ Links ]
30. Motti J.M.B., Rodenak B., Bailliet G., Bravi C.M. (2009). Tipificación de haplogrupos mitocondriales mediante multiplex PCR-AFLP. Novenas Jornadas Nacionales de Antropología Biológica, Puerto Madryn. [ Links ]
31. Motti, J. M. B., Muzzio, M., Ramallo, V., Rodenak Kladniew, B., Alfaro, E. L., Dipierri, J. E., Bailliet, G., & Bravi, C. M. (2013). Origen y distribución espacial de linajes maternos nativos en el noroeste y centro oeste argentinos/ Origin and spatial distribution of native maternal lineages in Northwest and Center West of Argentina. Revista Argentina De Antropología Biológica, 15(1), 03-14. Recuperado a partir de https://revistas.unlp.edu.ar/raab/article/view/607. [ Links ]
32. Motti, J., Schwab, M. E., Beltramo, J., Jurado-Medina, L. S., Muzzio, M., Ramallo, V., ... & Bravi, C. M. (2017). Diferenciación regional de poblaciones nativas de América a partir del análisis de los linajes maternos. Intersecciones en antropología, 18(3), 271-282. http://sedici.unlp.edu.ar/handle/10915/95143. [ Links ]
33. Motti, J.M.B. (2012). Caracterización de linajes maternos en la población actual del noroeste y centro-oeste argentinos. Facultad de Ciencias Naturales y Museo, UNLP. Tesis doctoral. [ Links ]
34. Muzzio, M., Motti, J. M. B., Chiarullo, S. M., Bravi, C. M., & Bailliet, G. (2012). Genealogical surveys show a high rate of non-paternal surname transmission with regional differences in Argentina. HOMO, 63(1), 43-49. DOI: 10.1016/j.jchb.2011.11.005. [ Links ]
35. Parolín, M. L., Avena, S. A., Fleischer, S., Pretell, M., Di Fabio Rocca, F., Rodríguez, D. A., ... & Carnese, F. R. (2013). Análisis de la diversidad biológica y mestizaje en la ciudad de Puerto Madryn (Prov. de Chubut, Argentina). Revista Argentina de Antropologia Biológica, 15(1), 61-75. Recuperado a partir de https://revistas.unlp.edu.ar/raab/article/view/693. [ Links ]
36. Paz Sepúlveda P. Jurado Medina L. Ramallo V., Muzzio M., Motti J., Santos M.R., Dipierri J., Gómez, E., Demarchi D. Bravi C., Salceda S., Bailliet, G. (2020). Linajes paternos autóctonos de Gran Chaco analizados con microsatélites. Revista del Museo de La Plata. 5. 553-562. 10.24215/25456377e129. [ Links ]
37. Race, Ethnicity and Genetics Working Group. (2005). The use of racial, ethnic, and ancestral categories in human genetics research. American Journal of Human Genetics, 77, 519-532. https://doi.org/10.1086/491747. [ Links ]
38. Ramallo V, Mucci JM, García A, y otros (2009) Comparison of Y-chromosome haplogroup frequencies in eight Provinces of Argentina. Forensic Science International: Genetics Supplement Series 2:431–432. DOI: https://doi.org/10.1016/j.fsigss.2009.08.047.
39. Ramírez Braschi, D. (2004) La guerra de la Triple Alianza a través de los periódicos correntinos. Corrientes: Moglia ediciones. [ Links ]
40. Raymond M. & Rousset F. (1995). An exact test for population differentiation. Evolution; 49:1280-1283. https://doi.org/10.2307/2410454. [ Links ]
41. Sala, A., Caputo, M., & Corach, D. (2019). Genetic structure of Mataco-Guaycurú speakers from Argentina and the extent of their genetic admixture with neighbouring urban populations. Scientific Reports, 9(1), 1-13. https://doi.org/10.1038/s41598-019-54146-6. [ Links ]
42. Sala A, Argüelles CF, Marino ME, Bobillo C, Fenocchio A, Corach D (2010) Genetic analysis of six communities of Mbyá-Guaraní inhabiting northeastern Argentina by means of nuclear and mitochondrial polymorphic markers. Human Biology 82(4):433-56. doi: 10.3378/027.082.0406. [ Links ]
43. Sans, M. (2009). “Raza”, adscripción étnica y genética en Uruguay. RUNA, Archivo Para Las Ciencias Del Hombre, 30(2), 163-174. https://doi.org/10.34096/runa.v30i2.771.
44. Schwab M. E. (2019). Afinidades filogeográficas y estructuración geográfica de los linajes maternos y paternos presentes en poblaciones humanas del Noroeste. Facultad de Ciencias Naturales y Museo, UNLP. Tesis doctoral. [ Links ]
45. Sevini F, Yao DY, Lomartire L, Barbieri A, Vianello D, Ferri G, et al., (2013) Analysis of population substructure in two sympatric populations of Gran Chaco, Argentina. PLoS One 8(5):e64054. doi: 10.1371/journal.pone.0064054. [ Links ]
46. Studer, E. F.S. (1958). La Trata de Negros en el Río de la Plata Durante el Siglo XVIII. Publicaciones del Instituto de Historia Argentina Doctor Emilio Ravignani. Buenos Aires, Universidad de Buenos Aires. Number 101. Pp. 378. [ Links ]
47. Tercer Censo Nacional. (1914). República Argentina. Población Tomo II. [ Links ]
48. Umetsu, K., Tanaka, M., Yuasa, I., Adachi, N., Miyoshi, A., Kashimura, S., … Osawa, M. (2005). Multiplex amplified product-length polymorphism analysis of 36 mitochondrial single-nucleotide polymorphisms for haplogrouping of East Asian populations. Electrophoresis, 26(1), 91–98. https://doi.org/10.1002/elps.200406129.
49. Umetsu, K., Tanaka, M., Yuasa, I., Saitou, N., Takeyasu, I., Fuku, N., … Osawa, M. (2001). Multiplex amplified product-length polymorphism analysis for rapid detection of human mitochondrial DNA variations. Electrophoresis, 22(16), 3533–8. https://doi.org/10.1002/1522-2683(200109)22:16<3533::AID-ELPS3533>3.0.CO;2-S.
50. Valenzuela, Cristina. 2014. “Principales Problemáticas y Potencialidades de La Región Nordeste.” Geograficando 10(2). Disponible en línea: http://www.geograficando.fahce.unlp.edu.ar/article/view/Geov10n02a07/html_24.

