










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista del Museo de Antropología
versión impresa ISSN 1852-060Xversión On-line ISSN 1852-4826
Rev. Mus. Antropol. vol.15 no.3 Córdoba dic. 2022
http://dx.doi.org/http://doi.org/10.31048/1852.4826.v15.n3.37850
DOI: http://doi.org/10.31048/1852.4826.v15.n3.37850
DOSSIER
Acumulaciones de restos óseos en nidos de carancho (Caracara plancus, Aves, Falconiformes) de la Patagonia Austral. Implicaciones tafonómicas y arqueológicas
Accumulations of bone remains in carancho (Caracara plancus, Aves, Falconiformes) nests from Southern Patagonia. Taphonomic and archaeological implications
María C. Álvarez1, María A. Gutiérrez1, Agustina Massigoge1, Luis A. Borrero2, Cristian A. Kaufmann1, Marcos Recofsky1 y Juan B. Belardi3
1INCUAPA, UNCPBA-CONICET. Universidad Nacional del Centro de la Provincia de Buenos Aires (UNCPBA), Facultad de Ciencias Sociales, Olavarría, Buenos Aires, Argentina.E-mails: malvarez@soc.unicen.edu.ar; mgutierr@soc.unicen.edu.ar; amassigo@soc.unicen.edu.ar; kaufman@soc.unicen.edu.ar; marcosrecofskyz@gmail.com
2IMHICIHU-CONICET. Saavedra 15. (C1083ACA) y Universidad de Buenos Aires, Ciudad Autónoma de Buenos Aires. E-mail: laborrero2014@gmail.com
3Universidad Nacional de la Patagonia Austral. Unidad Académica Río Gallegos. Laboratorio de Arqueología Dr. Luis. A. Borrero, ICASUR-CONICET. Piloto Lero Rivera s/n. Campus Universitario. (9400) Río Gallegos, Santa Cruz. E-mail: juanbautistabelardi@gmail.com
Recibido 01-06-2022
Recibido con correcciones 02-11-2022
Aceptado 06-11-2022
Resumen
Se estudiaron los restos faunísticos no ingeridos recuperados en el interior y bajo un nido de carancho (Caracara plancus) localizado en los niveles aterrazados del interfluvio Coyle-Gallegos con el objetivo general de evaluar su papel como agente tafonómico. El nido fue construido sobre una mata de calafate (Berberis sp.), ubicada en el centro de una concentración de carcasas de guanacos (Lama guanicoe) muertos por estrés invernal en el año 2020. Durante las tareas de campo se describieron y tomaron medidas del nido y se colectó su contenido. En el laboratorio realizamos identificaciones anatómicas y taxonómicas, además de relevar distintas variables tafonómicas. Los resultados indican el predominio de restos de guanaco, seguido por zorrino (Conepatus chinga), liebre (Lepus europaeus) y choique (Rhea pennata). Gran parte de los restos están desarticulados, fracturados y presentan marcas de mamíferos carnívoros. Al considerar los nidos de carancho en el marco de la arqueología regional, se destaca que el mayor potencial de contaminación del registro con los huesos acumulados por caranchos se daría en cañadones y roqueríos. Las observaciones y los resultados de los análisis permiten reconocer la incidencia del carancho como agente formador del registro, lo cual es relevante dado que por la escasa evidencia de su accionar, no suele ser considerado al momento de evaluar las historias tafonómicas de los conjuntos arqueológicos.
Palabras clave: Tafonomía Regional; Interfluvio Coyle-Gallegos; Aves Rapaces; Restos No Ingeridos; Guanaco.
Abstract
Non-ingested faunal remains recovered inside and under a carancho (Caracara plancus) nest located in the terraced levels of the Coyle-Gallegos interfluve are studied to evaluate its role as a taphonomic agent. The nest was built on a calafate (Berberis sp.) bush, located in the center of a concentration of guanaco (Lama guanicoe) carcasses killed by winter stress in 2020. The nest was described and measured during the fieldwork, and its contents were collected. In the laboratory, anatomical and taxonomic identifications were made, in addition to surveying different taphonomic variables. The results indicate the predominance of guanaco remains, followed by hog-nosed skunk (Conepatus chinga), hare (Lepus europaeus), and lesser rhea (Rhea pennata). Most of the remains are disarticulated and fractured, showing carnivorous mammal marks. When considering the carancho nests in the framework of regional archaeology, it should be noted that the greatest potential for contamination of the record with bones accumulated by caranchos would be in canyons and rocky areas. The observations and the results of the analyses allow us to recognize the incidence of the carancho as a record-forming agent, which is relevant given that, due to the scarce evidence of its action, it is not usually considered when evaluating the taphonomic histories of archaeological assemblages.
Keywords: Regional Taphonomy; Coyle-Gallegos interfluve; Raptors; Non-Ingested Remains; Guanaco.
Introducción
La investigación tafonómica naturalista de escala regional en ecosistemas modernos ha tenido un desarrollo importante a nivel mundial desde mediados de la década de 1970 (Hill 1976; Behrensmeyer 1978, 1993; Tappen 1995; Behrensmeyer y Dechant Boaz 1980; Gifford 1984; Blumenschine 1988; Domínguez-Rodrigo 2001; Faith y Behrensmeyer 2006). En sus inicios, uno de los principales objetivos de estos estudios fue determinar los procesos naturales que condicionan la depositación, modificación y preservación de huesos actuales de vertebrados para mejorar la comprensión de la información ecológica contenida en el registro fósil de sitios de homininos tempranos y sus posibles sesgos. En Argentina, los estudios tafonómicos pioneros fueron llevados a cabo especialmente por investigadores que estudiaban sitios de cazadores-recolectores de la Patagonia, quienes alentaron un enfoque regional de la tafonomía (Borrero 1988a, 1988b, 1989, 1990, 2001; Borrero y Martin 1996; Belardi y Carballo Marina 2003; Borella 2004; Cruz 2003, 2015). Estos autores propusieron que los análisis tafonómicos debían realizarse en la misma región donde se desarrollaban las investigaciones arqueológicas. El objetivo básico de este enfoque es entender la dinámica del ambiente en una escala espacial amplia; en particular los procesos responsables de la acumulación y la preservación ósea en diferentes estratos del paisaje, así como evaluar la lluvia natural de huesos (sensu Haynes 2018: 113) en el registro arqueológico regional (Borrero 1989, 2001).
Los estudios de tafonomía naturalista realizados en diferentes regiones de Argentina muestran que los depredadores son importantes agentes que acumulan y modifican los restos óseos. Dentro de estas investigaciones, los mamíferos carnívoros han recibido especial atención (Mondini 2018). En las últimas décadas, con el crecimiento de la tafonomía, los estudios sobre las aves como agentes formadores del registro cobraron mayor relevancia, aunque estos se han focalizado en el análisis de los restos ingeridos (e.g., Fernández et al. 2009, 2017; Montalvo et al. 2014, 2020; López 2020; Montalvo y Fernández 2019; López et al. 2021; Mignino 2021a, 2021b; Alcaraz y Kaufmann 2022). Como muestran distintos estudios a nivel global, las aves rapaces también pueden constituirse como importantes acumuladoras de restos óseos no ingeridos (e.g., Montalvo et al. 2011; Armstrong y Avery 2014; Lloveras et al. 2014; Mishra et al. 2017; Pardiñas et al. 2022). Estas investigaciones indican la acumulación de huesos en perchas y nidos de una gran variedad de presas, que incluyen ungulados domésticos, carnívoros, reptiles, aves y pequeños mamíferos. Además, en algunos nidos se han registrado huesos a los cuales se les asignaron funciones decorativas, defensivas y constructivas (Mishra et al. 2017).
Este trabajo se focaliza en la acción acumuladora de un ave rapaz ampliamente distribuida en América del Sur como es el carancho (Orden: Falconiformes, Familia: Falconidae). Las investigaciones tafonómicas previas sobre esta ave han estudiado muestras óseas acumuladas debajo de nidos y perchas y evidencias de su acción carroñera (Montalvo y Tallade 2009, 2010; Montalvo et al. 2011; Nasti 2021). Estas indicaron que el carancho genera acumulaciones de restos no ingeridos, con patrones de representación esquelética particulares y produce modificaciones en los huesos (Montalvo y Tallade 2009, 2010; Montalvo et al. 2011; Ballejo et al. 2016; Nasti 2021).
Durante los trabajos de campo realizados en febrero de 2021 en la planicie glacifluvial de los niveles aterrazados del interfluvio Coyle-Gallegos con el propósito de estudiar concentraciones de guanacos muertos por estrés invernal, se observaron numerosos nidos de carancho sobre matas de calafate (Berberis sp.). La mayoría de estos contenía gran cantidad de restos óseos, así como lana, huesos y plumas. Uno de ellos, que se encontraba asociado con una concentración de 33 carcasas de guanaco (Concentración 1) en una cota de 170 msnm, fue seleccionado para un análisis detallado de su contenido (Figura 1). Una capa de bosta de guanaco de un espesor de hasta 10 cm rodeaba parcialmente la mata. En marzo de 2022 se realizaron nuevos trabajos de campo en los que se identificaron nidos de carancho en el cañadón Mack Aike, que disecta la planicie glacifluvial. Dos de estos nidos fueron recolectados y sus materiales se encuentran en procesamiento.

Figura 1. Mapa del área de estudio con la localización del nido estudiado en este trabajo
Figure 1. Map of the study area with the nest’s location studied in this work.
El objetivo general de este trabajo es evaluar el papel del carancho como agente tafonómico. Los objetivos particulares son: a) determinar la representación taxonómica y anatómica del conjunto recuperado; b) evaluar la historia tafonómica del conjunto a partir del estudio de las modificaciones óseas; y c) discutir las oportunidades de preservación de estos restos y el potencial de contaminación del registro arqueológico regional. Este estudio pretende profundizar el conocimiento del papel de esta ave rapaz como agente acumulador, a través del análisis de un registro no explorado en investigaciones previas, como son los conjuntos de huesos transportados a los nidos. Si bien algunos naturalistas han informado sobre la presencia de huesos en los nidos (Hudson 1999 [1918]; Vigil 1973), hasta el presente no se ofrecieron descripciones detalladas de estos casos desde una perspectiva tafonómica o consideraciones acerca de sus implicaciones arqueológicas.
El Área de Estudio: Ambiente y Arqueología
El interfluvio de los ríos Coyle y Gallegos (S 51° 34´ 00´´; O 70° 38´ 00´´) tiene un clima templado-frío, con temperaturas medias anuales que oscilan entre los 0°C y los 12°C. Los inviernos son fríos a muy fríos por la acción de los vientos polares y subpolares y tienen una importante carga nival (Oliva et al. 2001). Los valores de precipitación anual se encuentran entre los 200 y los 300 mm. El paisaje corresponde a la denominada Unidad de Niveles Aterrazados, integrada por niveles glacifluviales del Pleistoceno, caracterizados por la presencia de material sedimentario que forma extensos mantos de relieve suave, con alturas de entre 138 y 170 msnm, con escasa pendiente y disectados por cañadones (Rial 2001). Los suelos tienen unos 30 cm de potencia y poseen un horizonte A1 de 10 cm de textura franco arcillo arenoso muy bien provisto de materia orgánica. Los restantes 20 cm corresponden al horizonte B2 arcilloso que apoya sobre rocas (Oliva et al. 2001). La comunidad vegetacional típica es la estepa graminosa seca, dominada por Festuca gracillima (coirón fueguino), que forma matas de 40 cm de altura. Entre estas matas se encuentran otras gramíneas o hierbas pequeñas (e.g., Rytidosperma virescens, Carex andina) así como individuos de Berberis sp. (calafate) o Mulguraea tridens (mata negra) (Prieto et al. 1999; Oyarzabal et al. 2018). La fauna principal está representada por el guanaco, carnívoros como el puma (Puma concolor) y el zorro colorado (Lycalopex culpaeus) y gris (Lycalopex griseus) y entre las aves se destaca el choique, el carancho, el águila mora (Geranoaetus melanoleucus), el chimango (Phalcoboenus chimango), el jote de cabeza negra (Coragypus atratus) y el cóndor andino (Vultur gryphus) (Albrieu y Ferrari 2000; Manero 2000).
La porción sur del área de estudio corresponde al límite norte del campo de lava Pali Aike donde se encuentran disponibles reparos rocosos, principalmente localizados sobre las laderas de las coladas y volcanes, que fueron utilizados por las poblaciones cazadoras recolectoras (Bird 1988; Barberena 2008; Borrero y Charlin 2010). Los trabajos arqueológicos que se desarrollan en el área de estudio lo hacen desde una perspectiva distribucional y en la misma escala espacial que el trabajo tafonómico (Borrero 2001). Sobre los niveles aterrazados se han registrado bajas densidades artefactuales líticas en superficie que pueden presentarse como concentraciones acotadas en bordes de lagunas temporarias que exponen rodados de distintas litologías, principalmente dacitas y rocas de grano fino oscuras (sensu Charlin 2005). Las ocupaciones humanas estuvieron probablemente acotadas a los meses cálidos. Se evidencia la realización de actividades de talla al paso que estarían principalmente relacionadas con la circulación de cazadores. A la vez, los reparos rocosos mencionados muestran un registro más concentrado que indicaría un uso más intenso de estas geoformas. Las mayores densidades y riquezas artefactuales y una dispersión prácticamente continua de artefactos asociados con arqueofaunas, principalmente de guanaco, ocurren en los fondos de valle de los cañadones que disectan los niveles aterrazados, en especial en el cañadón Mack Aike (Carballo Marina et al. 2019; Jones et al. 2019). Dicha señal, que se expresa tanto en superficie como en estratigrafía, es mucho más significativa que la registrada en los niveles aterrazados y en sus reparos y permite postular un uso redundante y residencial del cañadón, en cualquier estación del año. Esto tiene sentido en cuanto a la oferta de reparo, agua, leña y recursos faunísticos (guanacos y choiques) que ofrecen los cañadones.
Aspectos Eto-ecológicos del Carancho
El carancho es un ave rapaz diurna de la familia Falconidae. Tiene un tamaño de unos 51-64 cm de longitud y su masa corporal ronda alrededor de 1 kg (Fiora 1933; Ferguson-Lees y Christie 2001). Estas aves no tienen dimorfismo sexual, el plumaje de un adulto es de color negruzco (abdomen, corona, alas), el dorso y el pecho presentan una coloración barrada, la garganta posee tonos ocráceos, mientras que la cola es principalmente de color blanco, exceptuando el ápice de las plumas que es negro. Los individuos juveniles, en cambio, presentan una tonalidad más parda y estriada (Narosky e Yzurieta 2010). Su distribución geográfica es amplia y abarca todo el territorio argentino, donde ocupa una gran diversidad de ambientes que van desde llanuras a costas marinas e incluso áreas urbanas. También se lo encuentra en Bolivia, Brasil, Chile, Paraguay y Uruguay (Vigil 1973; Narosky e Yzurieta 2010). Tiene hábitos tróficos principalmente carroñeros, aunque en su dieta también son importantes las presas vivas en las que se incluyen mamíferos, aves, reptiles y artrópodos (Bó et al. 2007). Son depredadores oportunistas y generalistas (Engh et al. 1997; Travaini et al. 2001; Vargas et al. 2007; Formoso et al. 2019). Su alimentación varía dependiendo de la región; en Patagonia es primordialmente carroñera mientras que en la región pampeana la mayor parte de su dieta la constituyen las presas vivas (Bó et al. 2007). A diferencia de los individuos adultos, los juveniles tienen una alimentación basada principalmente en el consumo de carroña e invertebrados (Idoeta y Roesler 2012).
La construcción de los nidos de los caranchos se efectúa sobre las copas de los árboles de gran altura, pero en áreas desprovistas de este tipo de vegetación puede anidar sobre el suelo (Vigil 1973). En los paisajes llanos de la estepa patagónica austral, donde los sustratos naturales para anidar son raros, el empleo de matas de calafate o de acantilados costeros es muy común, solo precedido en términos de frecuencias por el uso de construcciones antrópicas (Saggesse et al. 2021: Tabla 1). El nido consiste en una estructura grande y compacta, que hace con gran cantidad de ramas y palos mezclados con huesos, estiércol y trozos de piel. El interior lo recubre con pastos secos, plumas, pelos y lanas (Vigil 1973; Hatcher 1985; Salvador 2013). El diámetro externo de los nidos es de unos 50-80 cm y su espesor de unos 40-50 cm (Goldstein 2000; Saggese et al. 2021). En el sur de Patagonia, la temporada de anidamiento ocurre desde septiembre a enero (Saggese et al. 2021). La puesta generalmente consta de tres a cuatro huevos de coloración blanca a castaña. El periodo de incubación dura aproximadamente un mes y está a cargo de la hembra. La alimentación de los pichones está a cargo de ambos miembros de la pareja (Salvador 2013). Una vez finalizada la época de cría, los caranchos abandonan el nido, que es habitualmente re-ocupado en la próxima puesta del año siguiente (Mallet 2016).
Materiales y Métodos
En el campo se describieron y tomaron medidas del nido, así como de la mata de calafate en la cual se hallaba. El nido, que se encontraba inactivo, fue desagregado y se recuperó el material óseo de su interior. Además, se recolectaron los huesos que se hallaban entrampados en las ramas de la mata, por debajo del nido (Figura 2). Las dos muestras (interior y debajo del nido) fueron analizadas y cuantificadas tanto de forma conjunta como por separado. Todo el proceso de registro y recuperación fue fotografiado.

Figura 2. a) localización del nido de carancho relevado en el año 2022, realizado en la misma ubicación que el analizado en este trabajo; b) vista desde arriba del contenido del nido; c) vista lateral del nido y del material debajo del mismo, entrampado en las ramas del calafate.
Figure 2. a) location of the carancho nest surveyed in 2022, in the same location as the one analyzed in this article; b) view from above of the nest content; c) side view of the nest and the material below it, entangled in the branches of the calafate.
En el laboratorio se limpió el material óseo en seco, que fue rotulado, pesado, fotografiado y analizado. Se relevó el taxón, la unidad anatómica y la presencia de tejido blando. En el caso de la meteorización, se consideraron los estadios del 0 al 5, propuestos por Behrensmeyer (1978). También se relevó la presencia y tipo de fractura (Johnson 1985). Se registró la presencia de marcas de dientes de mamíferos carnívoros, que fueron clasificadas en: perforaciones, hoyuelos, arrastres, furrows (se mantiene el término en inglés para evitar ambigüedades) y bordes crenulados (Haynes 1980; Binford 1981). También se consideraron las marcas producidas por aves, que suelen ser generadas por los picos en los huesos axiales para dar muerte a las presas (Serjeantson et al. 1993) o en epífisis de huesos de las alas para desgarrarlas y engullirlas más fácilmente (Bochenski y Tornberg 2003). Las aves carroñeras como jotes y buitres también pueden producir marcas con sus garras; estas son similares a los arrastres pero son más superficiales (Reeves 2009).
Cada resto se consideró como una ocurrencia, que incluye elementos desarticulados (un espécimen no relacionado con otro espécimen por el tejido blando); porciones articuladas (dos o más especímenes unidos por el tejido blando, que comprenden menos del 75% del esqueleto animal); y carcasas (especímenes unidos por el tejido blando, que comprenden más del 75% del esqueleto animal).
Se estimó el Número Mínimo de Individuos (NMI) teniendo en cuenta lateralidad y fusión, a partir del elemento óseo mejor representado. Se determinó la riqueza taxonómica a través del uso del Ntaxa (Grayson 1991), que considera la suma del número de taxones identificados, teniendo en cuenta los niveles taxonómicos de identificación alcanzados.
Para el caso del guanaco, taxón más abundante de la muestra, se construyó el perfil de partes esqueléticas a partir del cálculo del Número Mínimo de Elementos (NME) y Unidades Anatómicas Mínimas (MAU y MAU%) (Binford 1978; Klein y Cruz-Uribe 1984; Lyman 2008). Además, se estimó el desarrollo ontogénico de los individuos representados en el conjunto a través de la evaluación de la erupción y desgaste de los dientes y del estado de fusión de los distintos centros de crecimiento del esqueleto, incluyendo elementos apendiculares y axiales (Kaufmann 2009).
Resultados
La mata donde se hallaba el nido estudiado habría funcionado junto a otros tres calafates como reparo para guanacos durante eventos de olas de frío registrados en el área en el año 2020. Como producto del estrés invernal, murieron y quedaron depositados alrededor de los arbustos unos 33 guanacos de distinto sexo y edad. La mata tenía una altura de 1,9 m y era la más alta de las cuatro asociadas con la concentración de guanacos. El nido estaba conformado por ramas entrelazadas, junto con lana de guanaco, bosta y huesos de distintas especies (Figura 2; Tabla 1). Este tenía 0,5 m de diámetro y 0,25 m de espesor y su base se encontraba a 1,15 m del suelo. Se desconoce el momento de su formación.
Tabla 1. Representación taxonómica del material óseo. Referencias: MCM= masa corporal media; NOIN= n ocurrencias interior nido; NODN= n ocurrencias debajo nido; NA= no aplica.
Table 1. Taxonomic representation of bone material. References: MCM= mean corporal mass NOIN= n occurrences inside nest; NODN= n occurrences under nest; NA= not applicable.

Se recuperó un total de 52 ocurrencias óseas, 33 en el interior del nido y 19 por debajo de éste, entrampadas en las ramas de la mata. El peso total de la muestra es de 1515,05 g, siendo 162,69 g el peso máximo, correspondiente a una carcasa de zorrino y 0,66 g el mínimo, un fragmento de vértebra indeterminada. El promedio del peso de las ocurrencias es de 29,17 g. Además de los huesos, se registraron otros materiales de origen animal tales como cuero, pelos de guanaco, plumas, egagrópilas y bosta consolidada y disgregada de guanaco.
Los restos identificados corresponden a tres especies de mamíferos y una de aves (Tabla 1). El guanaco predomina ampliamente en el conjunto (88,46%), seguido en mucha menor frecuencia por el zorrino (3,85%), la liebre europea y el choique (ambos con 1,92%). Se estimó un número mínimo de cuatro individuos para guanaco (a partir del atlas y axis), dos para zorrino y uno para el resto de las especies identificadas.
Del total de las ocurrencias de guanaco, el 17,4% corresponde a unidades anatómicas articuladas y el 82,6% a huesos desarticulados. El 91,3% de las ocurrencias de guanaco presenta tejido blando, como restos de periostio, cuero, cartílago, tendones y pelo. En cuanto a las edades representadas, estimadas a través de la fusión de los huesos, predominan los individuos subadultos, particularmente las crías. Se destaca la presencia de al menos un individuo perinato, identificado a través de un cráneo (debajo del nido) y caja toráxica (interior del nido) (Figura 3).

Figura 3. Ocurrencias recuperadas: a) cráneo de guanaco perinato (debajo del nido); b) carcasa de zorrino (interior del nido); c) caja torácica de guanaco perinato (interior del nido); d) vértebras caudales de guanaco adulto (interior del nido); e) parte inferior de pata trasera de guanaco cría (debajo del nido). Escala= 1 cm.
Figure 3. Occurrences recovered: a) skull of perinate guanaco (under the nest); b) carcass of hog-nosed skunk (inside the nest); c) rib cage of perinate guanaco (inside the nest); d) caudal vertebrae of adult guanaco (inside the nest); e) lower part of the hind leg of newborn guanaco (under the nest). Scale= 1 cm.
El perfil de partes anatómicas de guanaco muestra un predominio del esqueleto axial, con los mayores valores para el atlas y el axis (100%, respectivamente), seguido por las vértebras caudales (66,7%) y las costillas (46,7%). En el caso del esqueleto apendicular, el fémur y la patela (50%, respectivamente) son los más representados, seguidos por los huesos del miembro anterior superior (Tabla 2, Figura 4).
Tabla 2. Representación anatómica de guanaco. Referencias: Izq=izquierdo, Der=derecho, Ax=axial, Indt=indeterminado V.=vértebra.
Table 2. Anatomical representation of guanaco. References: Izq=left, Der=right, Ax=axial, Indt=indeterminate V.=vertebra.

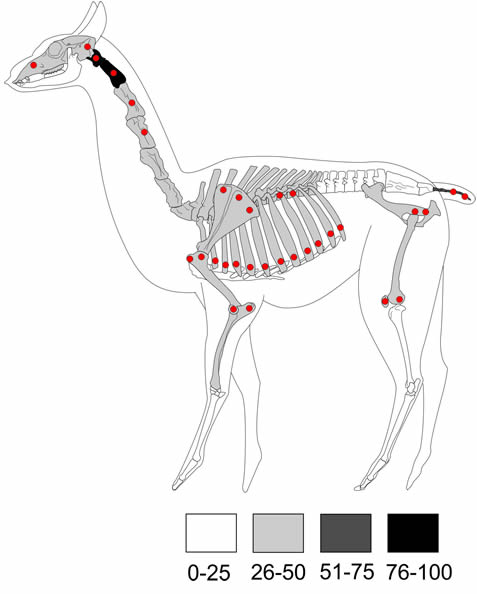
Figura 4. Representación de la abundancia anatómica de guanaco y distribución de las marcas de carnívoros.
Figure 4. Representation of guanaco anatomical abundance and distribution of carnivore tooth marks.
En lo que respecta al zorrino, se recuperaron dos individuos representados por una carcasa y una porción articulada, ambas con presencia de cuero con pelo y tejido conectivo. A la carcasa sólo le falta la porción distal de la cola y algunos fragmentos de las costillas (Figura 3). La porción articulada está compuesta por el cráneo, mandíbula, vértebras cervicales, escápulas y mitades proximales de húmeros.
En relación con las demás especies representadas, liebre europea y choique, sólo se recuperó una ocurrencia de cada una. Para la liebre europea se identificó una porción articulada del miembro posterior con tejido blando. La única ocurrencia del choique corresponde a la parte distal del fémur, sin tejido blando (Figura 5).

Figura 5. Ejemplos de modificaciones tafonómicas en las ocurrencias: a) furrows en olécranon de radiocúbito de guanaco; b) perforación y bordes crenulados en hemipelvis de guanaco; c) estadio 3 de meteorización en fémur de choique. Escala= 1 cm.
Figure 5. Examples of taphonomic modifications in the occurrences: a) furrows in olecranon of guanaco radiocubitus; b) perforation and crenulated edges in guanaco hemipelvis; c) stage 3 weathering in choique femur. Scale= 1 cm.
Los resultados del análisis de los efectos tafonómicos se presentan en la Tabla 3. En el 62% de las ocurrencias se observaron huesos fracturados. Sin embargo, más de la mitad de éstas tienen un elevado porcentaje de completitud (entre 75 y 100%). La mayoría de las ocurrencias no presentan evidencias de meteorización (63%) y de aquellas meteorizadas (37%), la mayor frecuencia se encuentra en el Estadio 1 (35%). Esta variable se expresa de manera diferente al comparar las muestras del interior y debajo del nido, con un mayor porcentaje de meteorización en esta última (18% y 68%, respectivamente). En relación con la presencia de modificaciones de mamíferos carnívoros, se registraron en el 60% de las ocurrencias, siendo más frecuentes en la muestra del interior del nido. Fueron identificados distintos tipos de marcas, con un predominio de bordes crenulados y perforaciones (38% y 35%, respectivamente). No pudieron identificarse marcas producidas por aves, aunque no puede descartarse que alguno de los arrastres o perforaciones pueda haber sido producto de este agente, pero en ese caso su incidencia sería mínima.
Tabla 3. Frecuencia absoluta y relativa de las variables tafonómicas. Referencias: NOIN=n ocurrencias interior nido; NODN= n ocurrencias debajo nido; NO= n ocurrencias.
Table 3. Absolute and relative frequency of taphonomic variables. References: NOIN=n occurrences inside nest; NODN= n occurrences under nest; NO= n occurrences.

Discusión
Este estudio muestra que los caranchos tienen la capacidad de acumular en su nido un importante número de huesos de animales de distinto tamaño (aproximadamente desde 2 a 100 kg de masa corporal). En este caso, se observó la presencia de restos óseos de al menos cuatro especies diferentes. La especie mejor representada es el guanaco, con un número mínimo de cuatro individuos, entre los cuales predominan los inmaduros, incluyendo un perinato. Los elementos esqueléticos de este taxón se presentan mayormente desarticulados y con escaso tejido blando. Se registraron elementos de todo el esqueleto, aunque predominan los axiales, en particular las vértebras cervicales y caudales. En el caso de los zorrinos, los dos individuos identificados corresponden a una carcasa con alta integridad y a una porción articulada. Ambas ocurrencias conservan el cuero con pelo. La liebre está representada por una porción articulada y el choique por un elemento desarticulado.
En relación con la capacidad de transporte, este análisis demuestra que el carancho puede trasladar hacia el nido restos de al menos 163 g de peso. Otros estudios mencionan el transporte de restos más pesados por esta rapaz. A modo de ejemplo, Borrero (1990: 366) observó el desplazamiento de cráneos de guanacos adultos recién muertos a través de distancias mayores a los 10 m. Nasti (2021) describe que el carancho transporta carroña a 130 m, aunque menciona que no constituye una especie que desarticule o transporte restos esqueléticos fuera del área de depositación original de la carcasa. No obstante, se considera que la falta de evidencia de transporte en su experimento puede obedecer a que las observaciones se realizaron en las etapas tempranas de la descomposición de las carcasas, cuando los elementos se encontraban mayormente articulados.
Un aspecto interesante del trabajo es que se observó una tendencia contraria a la registrada para otras aves que habitan el área de estudio. Los buitres del nuevo mundo, como por ejemplo el cóndor andino y el jote de cabeza negra, no suelen transportar huesos a sus nidos o dormideros. Ballejo et al. (2015) menciona que en general, estos carroñeros en general no transportan huesos de animales medianos y grandes desde los sitios de consumo a los de habitación, donde suelen recuperarse pequeños vertebrados y algunos huesos digeridos. En este análisis se observó una tendencia contraria para el caso del carancho, lo cual podría vincularse con la selección de huesos para la construcción del nido y/o alimentación, como se discutirá más adelante.
La elevada proporción de restos de guanaco se vincularía con la asociación espacial directa del nido con la acumulación de carcasas de estos camélidos muertos por estrés invernal. Esto generaría disponibilidad de carroña y lana que pudo haber constituido un factor que influyó en la elección del lugar de nidificación, además de la disponibilidad de matas de calafate como sustrato. Al respecto, en el último trabajo de campo que se realizó en el año 2022, se registró la construcción de un nuevo nido exactamente en el mismo calafate en el que se colectó el anterior (Figura 2). Además, se reconocieron otras situaciones de nidos vinculados espacialmente con concentraciones de guanacos muertos por estrés invernal. Es importante destacar que si bien se piensa que la fuente principal de los huesos de guanaco del nido son las carcasas disponibles en las inmediaciones del mismo, no se descarta que algunos restos hayan sido colectados a mayores distancias, como se observa en otros estudios. Tampoco puede dejar de considerarse la posibilidad de que se hayan carroñado carcasas de animales muertos en otras estaciones, como por ejemplo la primavera. Esto último puede vincularse con la presencia de un cráneo de un perinato, cuyo nacimiento en el área de estudio tiene lugar entre noviembre y diciembre.
Un porcentaje muy elevado de las ocurrencias se encuentra desarticulado y exhibe modificaciones por carnívoros. Éstas corresponden principalmente a las marcas típicas generadas por los mamíferos (Haynes 1983, Binford 1981). Predominan los restos con borde crenulado, seguido por los que presentan perforaciones y en menor medida arrastres, furrows y hoyuelos (Tabla 3). Dichas modificaciones podrían indicar la selección de elementos previamente desarticulados y consumidos por otros depredadores. Distintos autores han analizado acumulaciones de huesos generadas por buitres (Gyps fulvus) en sus nidos y observan que es frecuente la presencia de huesos con marcas de mamíferos carnívoros, los cuales serían producto del carroñeo sobre sitios de matanza de estos (Mundy y Ledger 1976; Plug 1978). En el caso de las presas medianas como el zorrino y la liebre, sobre la base de la presencia de marcas de mamíferos carnívoros, también habrían sido obtenidas mediante carroñeo. En concordancia, se ha señalado que los caranchos normalmente cazan presas con una masa corporal inferior a los 500 g (Rodríguez Estrella y Rivera Rodriguez 1997). En este caso, no se encontró ninguna marca claramente atribuible a la acción del carancho, a diferencia de Montalvo et al. (2011). Estos resultados están en línea con lo que mencionan Bochenski et al. (2009) acerca de que las aves que se alimentan de otros animales rara vez dejan evidencias directas de sus actividades.
Un interrogante que surge del análisis del conjunto es si todas las ocurrencias recuperadas fueron utilizadas como fuente de alimento o si algunas de ellas fueron seleccionadas para ser empleadas como material constructivo. El estado de preservación de los huesos, con muy baja o nula meteorización, sugiere el ingreso de los restos al nido en estado relativamente fresco, lo cual en principio concordaría con la hipótesis de alimentación.
En relación con el uso de los restos de guanaco como alimento, estudios realizados sobre egagrópilas de carancho en otros sectores de la Patagonia mostraron que los guanacos constituyen un componente importante de la dieta de estas aves (Engh et al. 1997; Travaini et al. 2001). Puntualmente en lo que refiere al transporte de restos de vertebrados grandes hacia los nidos, Travaini et al. (2001) señalan que este comportamiento se corresponde con una estrategia de minimización de viajes al nido, que disminuye el costo energético necesario para alimentar a las crías.
En el caso del uso de los huesos como material constructivo, esta idea podría apoyarse en que se registró una importante presencia de unidades esqueléticas con muy bajo aporte nutricional, como vértebras caudales articuladas y partes distales de las patas. El traslado de huesos por los caranchos a sus nidos con propósitos distintos a la alimentación ha sido mencionado por distintos naturalistas. Tempranamente, W. H. Hudson (1999 [1918]) refiere que en un hueco en ramas de “árbol grande y añoso … los caranchos habían construido su gran nido compuesto de palitos, panes de césped, huesos secos de oveja y de otros animales, pedazos de soga y de tiento y cualquier otro objeto que pudieran llevarse … dormían en el de noche y lo visitaban a distintas horas del día, por lo general llevando un hueso blanqueado, un tallo de cardo u otra cosa semejante para añadir a la pila” (Hudson 1999 [1918]: 84-85). Una descripción similar es ofrecida por C. Vigil (1973), quien, en referencia a los nidos de esta rapaz comenta: “Es una estructura grande y descuidada, que hace con ramas mezcladas con huesos, estiércol, pedazos de piel, etc., el interior lo recubre con pasto seco y plumas” (Vigil 1973:94).
Debido a la amplia distribución geográfica del carancho y a su capacidad para acumular huesos, esta ave debe ser considerada en la evaluación de los procesos de formación del registro arqueológico. Estos resultados permiten proponer que es un importante agente tafonómico, al cual debe prestarse especial atención, en particular cuando existe una superposición de las áreas de ocupación humana y de nidificación. Además, el carancho reutiliza los mismos sustratos de nidificación, e incluso los mismos nidos, aumentando su capacidad de acumulación en el largo plazo. En este sentido, estudios sobre su comportamiento de nidificación han mostrado la reutilización de un mismo sustrato para anidar (Saggese et al. 2021) así como de nidos de años previos (Engh et al. 1997; Goldstein 2000; Mallet 2016), lo que demuestra el uso a largo plazo de un mismo lugar. Mallet (2016) registró la reutilización de un mismo nido hasta cinco años consecutivos.
Además de acumular restos óseos, el carancho puede modificar conjuntos generados por los humanos mediante la sustracción de huesos. En este estudio, la relación espacial entre el nido y la concentración de carcasas de guanaco permite inferir la utilización de ésta última como fuente de aprovisionamiento. El comportamiento de sustracción de huesos fue observado por otros investigadores. Montalvo et al. (2011) mencionan que los caranchos frecuentemente sacan ventaja de los desechos humanos, llevándose restos de los basurales antrópicos. Asimismo, esta situación podría haber tenido lugar en el pasado. Por su parte, Ballejo et al. (2020) señalan que esta especie aprovecha los productos subsidiarios de los alimentos antrópicos, tales como basura. Esto ha llevado a un incremento en su población (Plaza y Lambertucci 2017; Zimmerman et al. 2019). Se considera que estas interacciones podrían dar lugar a conjuntos acumulados por caranchos que presentan huesos con modificaciones antrópicas. Esto ha sido observado en otros estudios actualistas con pequeños mamíferos carnívoros, que transportan a larga distancia restos desde los basurales de las casas de campo, hasta sus madrigueras, ubicadas en oquedades rocosas (Gutiérrez et al. 2016).
El potencial de entierro y preservación de los restos acumulados por carancho dependerá de las condiciones particulares de cada ambiente. En el caso del nido analizado en este trabajo, ubicado en las planicies glacifluviales del interfluvio de los ríos Coyle y Gallegos, la tasa de sedimentación es nula o baja. Solamente los depósitos acrecionales de bosta de guanaco ofrecen la posibilidad de enterramiento al menos en el corto plazo. En consecuencia, existen bajas chances de preservación de este registro. Esta situación genera que –exceptuando los casos de los depósitos de bosta- los huesos se destruyan antes de enterrarse, aunque en algunos sectores y en el corto plazo, podrían generarse asociaciones espurias en superficie entre materiales arqueológicos y huesos transportados por el carancho. Como se mencionó anteriormente, la firma arqueológica de estos espacios se caracteriza por amplias distribuciones de artefactos aislados. Solo las lagunas someras –muchas veces vecinas a las matas- concentran artefactos líticos.
En otros contextos ambientales del interfluvio Coyle-Gallegos puede esperarse otro tipo de situaciones en relación con el potencial de preservación y consecuente mezcla con materiales arqueológicos. Si bien aún no se han efectuado estudios sistemáticos, se han registrado nidos con huesos en matas de calafate sobre el borde de un maar volcánico, en las laderas de faldeos orientadas hacia el norte. En este sector también se identificó la presencia de artefactos arqueológicos. Si bien las posibilidades de enterramiento y perduración de los restos son similares a las de las planicies glacifluviales, el potencial de dispersión y mezcla con materiales arqueológicos es mayor debido a la pendiente, que dinamiza el escurrimiento ante el derretimiento de la nieve.
Otro sector en donde se reconoció nidos de carancho sobre matas es en los faldeos de los cañadones. En este lugar, al igual que en el borde del maar, es probable la dispersión causada por la pendiente, pero además existe un mayor potencial de enterramiento y mezcla en el fondo de los cañadones debido a la presencia de extensos mallines y a la amplia distribución de materiales arqueológicos. Por último, en los roqueríos de basalto que se encuentran en el sur del área de estudio se forman reparos que, además de entrampar sedimentos y preservar restos orgánicos, han sido loci ocupados reiteradamente por los seres humanos. Por ejemplo, inmediatamente al sur del área de estudio, Campan et al. (2007:687) observaron en torno a la cueva La Carlota nidos de bandurria reocupados por caranchos. Esto informa sobre las posibilidades de mezcla entre los huesos caídos de los nidos y los restos arqueológicos, situación sobre la cual -enfatizando el caso de los roedores- alertaron estas autoras. Cabe destacar que en el sondeo realizado en el sitio se recuperaron ramas procedentes de los nidos. Observaciones de nidos de carancho en roqueríos también fueron realizadas en el alero Montecarlo, Estancia Markatsch Aike, hacia el sudeste de la cueva La Carlota (K. Borrazzo, com. pers. 2022). Es claro que los abrigos han sido un factor de localización muy importante para los cazadores-recolectores, por lo que, aunque no sea el locus favorito del carancho, allí aumentan las oportunidades de contaminación.
Conclusiones
El carancho genera acumulaciones de huesos de las presas consumidas en las áreas de nidificación. Debido a que es una especie generalista y oportunista, el patrón de representación taxonómica puede ser muy diverso. Incluso traslada algunos elementos esqueléticos con propósitos constructivos, lo cual contribuye aún más a aumentar la diversidad de taxones que pueden estar presentes en sus nidos. En este caso de estudio se observó, observamos un claro predominio de restos esqueléticos de guanaco, lo cual estaría directamente relacionado con la asociación espacial del nido a una concentración de guanacos muertos por estrés invernal. Es posible incluso que este factor haya influido en la selección del lugar donde fue construido. Otro factor que debe considerarse es la posibilidad de acumulación de egagrópilas debajo de los nidos. Si bien en este caso éstas eran muy escasas, se cuenta con numerosos registros de bolos de otros sectores del área de estudio. El análisis de estos conjuntos permitirá realizar un estudio más integral de las acumulaciones generadas por el carancho.
La reutilización de los nidos por los caranchos a lo largo de distintas temporadas reproductivas puede conducir a la formación de importantes conjuntos de huesos, proceso que puede ser aún más acentuado en aquellos ambientes donde los sustratos de nidificación son limitados. Bajo condiciones de alta sedimentación, este tipo de conjuntos puede llegar a preservarse y pasar a formar parte del registro fósil. Debido a que una parte importante de los huesos transportados a los nidos son resultado del carroñeo, estos pueden presentar modificaciones de otros animales, como aves carroñeras y principalmente mamíferos carnívoros, que actuaron previamente sobre las carcasas o incluso por los humanos, lo cual complejiza la identificación del agente causal de la acumulación. Asimismo, si las áreas de nidificación coinciden con los sectores del paisaje seleccionados por los humanos como lugares de habitación, tal como pueden ser en el área de estudio los faldeos próximos al fondo de cañadones o los reparos rocosos, debe evaluarse necesariamente la posibilidad de mezcla de restos acumulados por los caranchos con materiales arqueológicos.
Debido a que el presente trabajo constituye el primer estudio del contenido de un nido de carancho desde una perspectiva tafonómica, y a que solo se ha analizado una muestra, no puede aún ofrecerse una caracterización general de este tipo de registro en términos de representación taxonómica y anatómica en nidos. Además, dado que el carancho es oportunista y generalista, su dieta dependerá de la oferta ambiental e incluso situacional (e.g., acumulación de guanacos muertos estrés invernal en el año 2020). Asimismo, no se ha reconocido ningún tipo de modificación en los huesos que ayude a la identificación de este agente. Las marcas que se han observado en este análisis fueron producidas por carnívoros mamíferos que estuvieron involucrados en el consumo de las carcasas antes de ser obtenidas por los caranchos. Es esperable que este tipo de marcas sean comunes en acumulaciones de caranchos en nidos, ya que es frecuente la alimentación de los pichones con carroña. Es posible, entonces, que no exista un conjunto “típico” de acumulación para esta ave rapaz. Sin embargo, la dificultad para reconocer la acción del carancho como agente tafonómico no debe desalentar la evaluación de su participación en la formación de conjuntos particulares. Por el contrario, esto obliga a reflexionar sobre lo complejas que pueden ser las historias tafonómicas de las acumulaciones, lo cual puede pasar inadvertido si nos concentramos en buscar lo obvio. Asimismo, este hecho impulsa a seguir desarrollando herramientas metodológicas que eventualmente puedan ayudar a reconocer la firma tafonómica de este agente que al día de hoy parece actuar de modo invisible. Para esto, se debe analizar un mayor número de nidos que sean representativos de la variabilidad de situaciones ambientales de nuestra área de estudio. La tafonomía actualista justamente ofrece la posibilidad de evaluar escenarios con agentes y procesos que de otro modo no se presentan como evidentes en el registro arqueológico.
Agradecimientos
Estas investigaciones se realizaron en el marco del convenio UNPA-UNICEN (Resolución CS 7938-21) y con el financiamiento del Programa de Fortalecimiento a la Ciencia y la Tecnología en las Universidad Nacionales - Universidad Nacional del Centro de la Provincia de Buenos Aires 03-JOVIN-75F, Unidad Ejecutora INCUAPA-CONICET-UNICEN (PUE 22920160100079), PICT 2018-0686 y UNPA 29/A476-1 (2021-2023). Agradecemos a Julián Mignino, Paula Weihmüller y José Manuel López, organizadores del II Taller de Tafonomía Actualística: Metodología y Aplicaciones Arqueológicas por la oportunidad brindada para presentar nuestras investigaciones; a las Dras. Flavia Carballo Marina y Karen Borrazzo por facilitarnos información y contribuir a la discusión de las implicaciones de este trabajo. A los Sres. Brian Halvorsen (Estancia Bella Vista) y Claudio Ruibal (Estancia Alquinta), por su hospitalidad e inestimable colaboración durante los trabajos de campo. A dos evaluadores anónimos, que con sus comentarios y sugerencias realizaron valiosos aportes a este artículo.
1. Albrieu, C. y Ferrari, S. (2000). La estepa. En El Gran Libro de la Provincia de Santa Cruz. Tomo 1, 280-301. España, Milenio Ediciones y Alfa Centro Literario. [ Links ]
2. Alcaraz, A. P. y Kaufmann, C. A. (2022). Patrón de consumo de anuros por Athene cunicularia: Implicancias para el registro arqueológico de la región Pampeana. Comenchigonia. Revista de Arqueología, en prensa. [ Links ]
3. Armstrong, A. y Avery, G. (2014). Taphonomy of Verreaux’s Eagle (Aquila verreauxii) prey accumulations from the Cape Floral Region, South Africa: Implications for archaeological interpretations. Journal of Archaeological Science, 52, 163-183. https://doi.org/10.1016/j.jas.2014.08.024
4. Ballejo, F., Fernández, F. J., Montalvo, C. I., y De Santis, L. (2016). Taphonomy and dispersion of bones scavenged by New World vultures and caracaras in Northwestern Patagonia: implications for the formation of archaeological sites. Archaeological and Anthropological Sciences, 8(2), 305-315. http://dx.doi.org/10.1007/s12520-015-0277-9 [ Links ]
5. Ballejo, F., Plaza, P. I., y Lambertucci, S. A. (2020). The conflict between scavenging birds and farmers: Field observations do not support people’s perceptions. Biological Conservation, 248(April), 108627. https://doi.org/10.1016/j.biocon.2020.108627
6. Barberena, R. (2008). Arqueología y biogeografía humana en Patagonia meridional. Buenos Aires, Sociedad Argentina de Antropología. [ Links ]
7. Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology, 1, 150-162. https://doi.org/10.1017/S0094837300005820 [ Links ]
8. Behrensmeyer, A. K. (1993). The bones of Amboseli: bone assemblages and ecological change in a modern African ecosystem. National Geographic Research, 9, 402-421. [ Links ]
9. Behrensmeyer, A. K. y Dechant Boaz, D. E. (1980). The recent bones of Amboseli Park, Kenya in relation to East African paleoecology. En: Behrensmeyer, A. K. y Hill A. P. (eds), Fossils in the making, pp. 72-92. Chicago, University of Chicago Press. [ Links ]
10. Belardi, J. B. y Carballo Marina, F. (2003). Tafonomía regional en la cuenca media del río Coyle (Santa Cruz, Patagonia Argentina). Intersecciones en Antropología, 4, 59-73. [ Links ]
11. Binford, L. R. (1978). Nunamiut ethnoarchaeology. New York, Academic Press. [ Links ]
12. Binford, L. R. (1981). Bones: Ancient men and modern myths. New York, Academic Press. [ Links ]
13. Bird, J., (1988). Travels and Archaeology in South Chile. Iowa City, University of Iowa Press. [ Links ]
14. Blumenschine, R. J. (1988). An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages. Journal of Archaeological Science, 15(5), 483-502. https://doi.org/10.1016/0305-4403(88)90078-7 [ Links ]
15. Bó, M. S., Baladron, A. V. y Biondi, L. M. (2007). Ecología trófica de Falconiformes y Strigiformes: Tiempo de síntesis. Hornero, 22, 97-115. [ Links ]
16. Bochenski, Z. M. y Tornberg, R. (2003). Fragmentation and preservation of bird bones in uneaten food remains of the Gyrfalcon Falco rusticolus. Journal of Archaeological Science, 30 (12), 1665-1671. https://doi:10.1016/S0305-4403(03)00069-4 [ Links ]
17. Bochenski, Z. M., Tomek, T., Tornberg, R. y Wertz, K. (2009). Distinguishing nonhuman predation on birds: patterns of damage done by the white-tailed eagle Haliaeetus albicilla, with comments on the punctures made by the golden eagle Aquila chrysaetos. Journal of Archaeological Science, 36, 122-129. https://doi.org/10.1016/j.jas.2008.07.018 [ Links ]
18. Borella, F. (2004). Tafonomía regional y estudios arqueofaunísticos de cetáceos en Tierra del Fuego y Patagonia meridional. Oxford, British Archaeological Reports. [ Links ]
19. Borrero, L. A. (1988a). Tafonomía regional. En N. Ratto y A. Haber (Eds.), De contextos, procesos y otros huesos, pp. 9-15. ICA (FFyL-UBA). [ Links ]
20. Borrero, L. A. (1988b). Estudios tafonómicos en Tierra del Fuego: su relevancia para entender procesos de formación del registro arqueológico. En H. Yacobaccio (Ed.), Arqueología Contemporánea Argentina. Actualidad y Perspectivas, pp. 13-32. Buenos Aires, Ediciones Búsqueda. [ Links ]
21. Borrero, L. A. (1989). Sites in action: the meaning of guanaco bones in Fueguian archaeological sites. ArchaeoZoologia, III, 9-24. [ Links ]
22. Borrero, L. A. (1990). Taphonomy of guanaco bones in Tierra del Fuego (Argentina). Quaternary Research, 34, 361-371. https://doi.org/10.1016/0033-5894(90)90047-O [ Links ]
23. Borrero, L. A. (2001). Regional taphonomy: background noise and the integrityof the archaeological record. En: Kuznar, L. (Ed.), Ethnoarchaeology of Andean South America: contributions to archaeological method and theory. Ann Arbor (MI): International Monographs in Prehistory.Ethnoarchaeological Series 4; p. 243-254. [ Links ]
24. Borrero, L. A. y Martin, F. M. (1996). Tafonomía de carnívoros: un enfoque regional. En: Gómez Otero, J. (Ed.). Arqueología: sólo Patagonia, pp. 189-198. Puerto Madryn, CENPAT-CONICET. [ Links ]
25. Borrero, L. A. y Charlin, J. (Eds.). (2010). Arqueología de Pali Aike y Cabo Vírgenes (Santa Cruz, Argentina). Buenos Aires, CONICET-IMHICIHU. [ Links ]
26. Campan, P., Carballo Marina, F. y Manzi, L. (2007). Arqueología de estancia La Carlota (campo volcánico Pali Aike, Santa Cruz). En: Morello, F., Martinic, M., Prieto, A. y Bahamonde, G. (Eds.), Arqueología de Fuego-Patagonia. Levantando piedras, desenterrando huesos... y develando arcanos, pp. 687-699. Punta Arenas, Editorial CEQUA. [ Links ]
27. Carballo Marina, F., Belardi, J. B. y Tiberi, P. (2019). Un hallazgo inusual: un núcleo de ópalo y calcedonia en la cuenca media del río Gallegos (Santa cruz, Argentina). Magallania, 47(2), 125-131. http://dx.doi.org/10.4067/S0718-22442019000200125 [ Links ]
28. Charlin, J. (2005). Utilización de materias primas líticas en el Campo Volcánico Pali Aike (Pcia. Santa Cruz, Argentina). Werken, 7, 39-56. [ Links ]
29. Cruz, I. (2003). Paisajes tafonómicos de restos de Aves en el sur de Patagonia continental. Aportes para la interpretación de conjuntos avifaunísticos en registros arqueológicos del Holoceno. [Tesis de Doctorado, Facultad de Filosofía y Letras, Universidad de Buenos Aires]. [ Links ]
30. Cruz, I. (2015). Las investigaciones sobre preservación de huesos de aves y mamíferos grandes en Patagonia (Argentina). Archaeofauna, 24, 209-224. [ Links ]
31. Domínguez-Rodrigo, M. (2001). A study of carnivore competition in riparian and open habitats of modern savannas and its implications for hominid behavioral modeling. Journal of Human Evolution, 40(2), 77-98. https://doi.org/10.1006/jhev.2000.0441 [ Links ]
32. Engh, A. L., Frnklin, W. I. y Sarno, R. J. (1997). Breeding biology and Food habits of the Andean Crested Caracara (Polyborus plancus plancus) in the Patagonia of southern Chile. Vida Silvestre Neotropical, 6, 48-52. [ Links ]
33. Faith, J. T. y Behrensmeyer, A. K. (2006). Changing patterns of carnivore modification in a land-scape bone assemblage, Amboseli Park, Kenya. Journal of Archaeological Science, 33(12), 1718-1733. https://doi.org/10.1016/j.jas.2006.03.004 [ Links ]
34. Ferguson-Lees, J. y Christie, D.A. (2001). Raptors of the world. Christopher Helm, London. [ Links ]
35. Fernández, F. J., Moreira, G., Ferraro, D. y De Santis, L. (2009). Presas consumidas por la lechuza de campanario (Tyto alba) en la localidad de Olavarría, Buenos Aires: un caso de elevada batracofagia. Nuestras Aves, 54, 20-21 [ Links ]
36. Fernández, F. J., Montalvo, C. I., Fernández-Jalvo, Y., Andrews, P. y López, J. M. (2017). A re-evaluation of the taphonomic methodology for the study of small mammal fossil assemblages of South America. Quaternary Science Review, 155, 37-49. https://doi.org/10.1016/j.quascirev.2016.11.005 [ Links ]
37. Fiora, A. (1933). El peso de las aves. Hornero, 05(02), 174-188. [ Links ]
38. Formoso, A. E., Agüero, L. y Udrizar Sauthier, D. E. (2019). Diet of the Southern Caracara in a near-shore insular system in southern Patagonia, Argentina. Journal of King Saud University – Science, 31, 1339-1343. https://doi.org/10.1016/j.jksus.2018.10.001
39. Gifford, D. P. (1984). Taphonomic specimens, Lake Turkana. En: National Geographic Research Reports, National Geographic Research Reports, 17, 419-428. [ Links ]
40. Goldstein, M. I. (2000). Nest-site characteristics of Crested Caracaras in La Pampa, Argentina. Journal of Raptor Research, 34, 330-333. [ Links ]
41. Grayson, D. K. (1991). Alpine faunas from the White Mountains, California: Adaptive change in the late prehistoric Great Basin? Journal of Archaeological Science, 18, 483-506. https://doi.org/10.1016/0305-4403(91)90039-R [ Links ]
42. Gutiérrez, M. A., Kaufmann, C. A., González, M. E., Scheifler, N. A., Rafuse, D. J., Massigoge, A. y Álvarez, M. C. (2016). The role of small carnivores in the movement of bones: implications for the Pampas archaeofaunal record, Argentina. Archaeological and Anthropological Sciences, 8(2), 257-276. http://doi.org/10.1007/s12520-015-0272-1. [ Links ]
43. Hatcher, J. B. (1985). Bone hunters in Patagonia. Narrative of the expedition. Woodbridge, Ow Bow Press. [ Links ]
44. Haynes G. (1980). Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology, 6 (3), 341-351. https://doi:10.1017/ S0094837300006849. [ Links ]
45. Haynes, G. (1983). A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones. Paleobiology, 9 (2), 164-172. https://doi.org/10.1017/S0094837300007545 [ Links ]
46. Haynes, G. (2018). Raining more than cats and dogs: Looking back at field studies of noncultural animal-bone occurrences. Quaternary International, 466, 113-130. https://doi.org/10.1016/j.quaint.2015.10.119 [ Links ]
47. Hill, A. (1976). On Carnivore and Weathering Damage to Bone. Current Anthropology, 17(2), 335-336. https://doi.org/10.1086/201732 [ Links ]
48. Hudson, W. H. (1999 [1918]). Allá lejos y hace tiempo. Buenos Aires, Emecé [ Links ].
49. Idoeta, F. M. y Roesler, I. (2012). Presas consumidas por el carancho (Caracara plancus) durante el periodo reproductivo, en el noreste de la Provincia de Buenos Aires. Nuestras Aves, 57, 80-83. [ Links ]
50. Johnson, E. (1985). Current developments in bone technology. Advances in Archaeological Method and Theory, 8, 157-235. https://doi.org/10.1016/B978-0-12-003108-5.50010-5 [ Links ]
51. Jones, E. L., Taylor, W. T., Belardi, J. B., Neme, G., Gil, A., Roberts, P., Thornhill, C., Hodgins, G. W. L. y Orlando, L. (2019). Caballos y humanos en el nuevo mundo: investigaciones arqueológicas en América del Norte y perspectivas para Argentina. Anales de Arqueología y Etnología, 74 (2), 247-268. [ Links ]
52. Kaufmann, C. A. (2009). Estructura de edad y sexo en Lama guanicoe (guanaco). Estudios actualísticos y arqueológicos en Pampa y Patagonia. Buenos Aires, Sociedad Argentina de Antropología. [ Links ]
53. Klein, R. G. y Cruz-Uribe, K. (1984). The Analysis of Animal Bones from Archaeological Sites. Chicago, The University of Chicago Press. [ Links ]
54. López, J. M. (2020). Actualistic taphonomy of barn owl pellet-derived small mammal bone accumulations in arid environments of South America. Journal of Quaternary Science, 35 (8), 1057-1069. https://doi.org/10.1002/jqs.3251 [ Links ]
55. López, J. M., Quintana, M. F., Mignino, J. y Pereyra-Lobos, R. (2021). Tafonomía actualística de restos óseos y dentarios de micromamíferos recuperados en egagrópilas generadas por la lechucita de las vizcacheras (Athene cunicularia): una discusión desde las tierras bajas áridas del Centro Occidente Argentino. Intersecciones en Antropología, 22 (1), 55-67. https://doi.org/10.37176/iea.22.1.2021.589 [ Links ]
56. Lyman, R. L. (2008). Quantitative Paleozoology. Cambridge, Cambridge University Press. [ Links ]
57. Lloveras, L., Nadal, J., Moreno-García, M., Thomas, R., Anglada, J., Baucells, J., Martorell, C. y Vilasís, D. (2014). The role of the Egyptian Vulture (Neophron percnopterus) as a bone accumulator in cliff rock shelters: An analysis of modern bone nest assemblages from North-eastern Iberia. Journal of Archaeological Science, 44, 76-90. https://doi.org/10.1016/j.jas.2014.01.018 [ Links ]
58. Mallet, J. (2016). Uso y selección de sitio reproductivo y hábitos tróficos del carancho (Caracara plancus) en el noreste de La Pampa. [Tesis de Licenciatura, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa].
59. Manero, A. (2000). Los mamíferos de Santa Cruz. En: El Gran Libro de la Provincia de Santa Cruz. Tomo 1:311-319. España, Milenio Ediciones y Alfa Centro Literario. [ Links ]
60. Mignino, J. (2021a). Tafonomía de restos óseos y dentarios de micromamíferos recuperados en egagrópilas generadas por Megascops choliba (Strigiformes) en ambientes boscosos del centro de Argentina. Intersecciones en Antropología, 22, 69-81. https://doi.org/10.37176/iea.22.1.2021.590 [ Links ]
61. Mignino, J. (2021b). Tafonomía actualística de huesos de anfibios predados por estrigiformes del centro de Argentina. Contribuciones para estudios paleontológicos y arqueológicos. Spanish Journal of Palaeontology, 36 (1), 79-90. https://doi.org/10.7203/sjp.36.1.20517 [ Links ]
62. Mishra, S., Kumar, A., y Kanaujia, A. (2017). Nest Material selection by Egyptian Vultures. Journal of Entomology and Zoology Studies, 5 (6), 1649-1655. [ Links ]
63. Mondini, M. (2018). Carnivore taphonomy in South America: a review of actualistic studies and their implications in the southern Neotropics. Historical Biology, 30 (6), 774-785. https://doi.org/10.1080/08912963.2017.1319831 [ Links ]
64. Montalvo, C. I., Tallade, P. (2009). Taphonomy of the accumulations produced by Caracara plancus (Falconidae). Analysis of prey remains and pellets. Journal of Taphonomy, 7 (2-3), 235- 248. [ Links ]
65. Montalvo, C. I. y Tallade, P. O. (2010). Análisis tafonómico de restos no ingeridos de roedores presa de Caracara plancus (Aves, Falconidae). En: De Nigris, M., Fernández, P.M., Giardina, M., Gil, A.F., Gutiérrez, M.A., Izeta, A., Neme, G., Yacobaccio, H.D. (Eds.), Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio. Ediciones del Espinillo, Buenos Aires, Argentina, pp. 419e428. [ Links ]
66. Montalvo, C. I., Tallade, P. O., Fernández, F. J., Moreira, G. J., Rafuse, D. J., De Santis, L. (2011). Bone damage patterns found in the avian prey remains of crested caracara Caracara plancus (Aves, Falconiformes). Jornal of Archaeological Science, 38 (12), 3541-3548. https://doi.org/10.1016/j.jas.2011.08.021 [ Links ]
67. Montalvo, C. I., Fernández, F. J., Liébana, M. S., Santillán, M. y Sarasola, J. H. (2014). Taphonomic analysis of rodent bone accumulations produced by the White-tailed Kite (Elanus leucurus, Accipitriformes) in Central Argentina. Journal of Archaeological Science, 52, 354-362. https://doi.org/10.1016/j.jas.2014.09.003 [ Links ]
68. Montalvo, C. I., Fernández, F. J., Galmes, M. A., Santillán, M. A., y Cereghetti, J. (2016). Crowned solitary eagle (Buteogallus coronatus) as accumulator of armadillo osteoderms in the archaeological record? An actualistic taphonomic study for central Argentina. Quaternary International, 391, 90-99. https://doi.org/10.1016/j.quaint.2015.08.017 [ Links ]
69. Montalvo, C. I. y Fernández. F. J. (2019). Review of the actualistic taphonomy of small mammals ingested by South American predators. Its importance in the interpretation of the fossil record. Publicación Electrónica de la Asociación Paleontológica Argentina, 19, 18-46. https://doi.org/10.5710/PEAPA.11.03.2019.275 [ Links ]
70. Montalvo, C. I., Fernández, F. J., Tomassini, R. L., Mignino, J., Kin, M. S. y Santillán, M. A. (2020). Spatial and temporal taphonomic study of bone accumulations of the burrowing owl (Athene cunicularia) in central Argentina. Journal of Archaeological Science: Reports, 31, 102197. https://doi.org/10.1016/j.jasrep.2020.102197 [ Links ]
71. Mundy, P. J. y Ledger, J. A. (1976). Griffon vultures, carnivores, and bones. South African Journal of Science, 72 (4), 106-110. [ Links ]
72. Narosky, T. e Yzurieta, D. (2010). Aves de Argentina y Uruguay, Guía de Identificación. Buenos Aires, Editorial Vázquez Mazzini Editores, 16ª ed. [ Links ]
73. Nasti, A. (2021). Carroñeo y dispersión de restos de cerdo doméstico (Sus scrofa) en contextos de humedales: implicancias forenses. Revista Argentina de Antropología Biológica, 23 (2), 038. https://doi.org/10.24215/18536387e038 [ Links ]
74. Oliva, G., González, L ., Rial, P. y Livrachi, E. (2001). Áreas Ecológicas de Santa Cruz y Tierra del Fuego. En Borrelli, P. y Oliva, G. (Eds.). Ganadería ovina sustentable en la Patagonia Austral. Tecnología de manejo extensivo, pp. 41- 82. Buenos Aires, INTA. [ Links ]
75. Oyarzabal, M., Clavijo, J., Oakley, L., Biganzoli, F., Tognetti, F., Barberis, I., Maturo, H. M., Aragón, R., Campanello, P. I., Prado, D., Oesterheld, M. y León, R. J. C. (2018). Unidades de vegetación de la Argentina. Ecología Austral, 28 (1), 040-063. [ Links ]
76. Pardiñas, U. F. J., Borrero, L. A., Martin, F. M., Massone, M. y Fernández, F. J. (2022). Fossil rodents in Mylodon Cave as indicators of late Pleistocene–Holocene environmental evolution in southern Chile. Quaternary Research, 105, 218-234. https://doi.org/10.1017/qua.2021.44
77. Plaza, P. I. y Lambertucci, S. A. (2017). How are garbage dumps impacting vertebrate demography, health, and conservation? Global Ecology and Conservation, 12, 9-20. https://doi.org/10.1016/j.gecco.2017.08.002 [ Links ]
78. Plug, I. (1978). Collecting patterns of six species of vultures (Aves: Accipitridae). Annals of the Transvaal Museum, 31 (6), 51-63. [ Links ]
79. Prieto, A. R., Stutz, S. y Pastorino, S. (1999). Arqueopalinología de la cueva Las Buitreras. Praehistoria, 3, 169-181. [ Links ]
80. Reeves, N. M. (2009). Taphonomic effects of vulture scavenging. Journal of Forensic Sciences, 54 (3), 523-528. https://doi.org/10.1111/j.1556-4029.2009.01020.x [ Links ]
81. Rial, P. (2001). Grandes unidades de paisaje. En Borrelli, P. y Oliva, G. (Eds.). Ganadería ovina sustentable en la Patagonia Austral. Tecnología de manejo extensivo, pp. 22-82. Buenos Aires, INTA. [ Links ]
82. Tappen, M. (1995). Savanna ecology and natural bone deposition: Implications for early hominid site formation, hunting, and scavenging. Current Anthropology, 36 (2), 223-260. https://10.1086/204353 [ Links ]
83. Rodríguez Estrella, R. y Rivera Rodríguez, L. B. (1997). Crested caracara food habits in the Cape region of Baja California, Mexico. Journal of Raptor Research, 31 (3), 228e233. [ Links ]
84. Saggese, M.D., Morrison, J.L., Quaglia, A.I.E., Ellis, D.H., Ellis C.H., Nelson, R.W., Amorós, M.B., Amorós, C.D. y Cadierno, S. A. (2021). Breeding Ecology of Southern Caracaras (Caracara plancus) in Southern Continental Patagonia, Santa Cruz Province, Argentina. Journal of Raptor Research, 55 (2), 190-200. https://doi.org/10.3356/0892-1016-55.2.190 [ Links ]
85. Salvador, S. (2013). Reproducción del Carancho (Caracara plancus) en Villa María, Córdoba, Argentina. (Aves, Falconidae). Xolmis, 27, 1-5. [ Links ]
86. Serjeantson, D., Irving, B. y Hamilton-Dyer, S. (1993). Bird bone taphonomy from the inside out: the evidence of gull predation on the Manx Shearwater Puffinus puffinus. Archaeofauna, 2, 191-204. [ Links ]
87. Travaini, A., Donázar, J. A., Ceballos, O. y Hiraldo, F. (2001). Food habits of the Crested Caracara (Caracara plancus) in the Andean Patagonia: the role of breeding constraints. Journal of Arid Environments, 48, 211e219. https://doi.org/10.1006/jare.2000.0745 [ Links ]
88. Vargas, R. J., Bó, M. S. y Favero, M. (2007). Diet of the Southern Caracara (Caracara plancus) in Mar Chiquita Reserve, Southern Argentina. Journal of Raptor Research, 41 (2), 113-121. https://doi.org/10.3356/0892-1016(2007)41[113:DOTSCC]2.0.CO;2 [ Links ]
89. Vigil, C. (1973). Aves Argentinas y Sudamericanas. Buenos Aires, Editorial Atlántida. [ Links ]
90. Zimmerman, G. S., Millsap, B. A., Avery, M. L., Sauer, J. R., Runge, M. C. y Richkus K. D. (2019). Allowable take of black vultures in the eastern United States. Journal of Wildlife Management, 83, 272-282. https://doi.org/10.1002/jwmg.21608 [ Links ]

