










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista del Museo de Antropología
versión impresa ISSN 1852-060Xversión On-line ISSN 1852-4826
Rev. Mus. Antropol. vol.15 no.3 Córdoba dic. 2022
http://dx.doi.org/http://doi.org/10.31048/1852.4826.v15.n3.38010
DOI: http://doi.org/10.31048/1852.4826.v15.n3.38010
DOSSIER
Observaciones tafonómicas naturalistas en el Sistema Lagunar Hinojo-Las Tunas (Campo de Dunas del Centro Pampeano, Argentina)
Naturalistic taphonomic observations in the Hinojo-Las Tunas Shallow Lake System (Central Pampean Dune Field, Argentina)
Nahuel A. Scheifler1, Agustina Massigoge1, María Clara Álvarez1, Juan M. Rodríguez1, Marcos Recofsky1, Cristian A. Kaufmann1, Mariela E. González1 y María A. Gutiérrez1
1INCUAPA, UNCPBA-CONICET. Universidad Nacional del Centro de la Provincia de Buenos Aires (UNCPBA), Facultad de Ciencias Sociales, Olavarría, Buenos Aires, Argentina. E-mails: nscheifler@soc.unicen.edu.ar;
amassigo@soc.unicen.edu.ar; malvarez@soc.unicen.edu.ar; juanmanuel.rodriguez@soc.unicen.edu.ar; marcosrecofskyz@gmail.com; ckaufman@soc.unicen.edu.ar; mgonzalez@soc.unicen.edu.ar; mgutierr@soc.unicen.edu.ar
Recibido 15-06-2022
Recibido con correcciones 11-08-2022
Aceptado 29-08-2022
Resumen
En este trabajo se presentan los resultados de las primeras observaciones tafonómicas actualistas desarrolladas en dos lagunas salinas del Sistema Lagunar Hinojo-Las Tunas, ubicado en el Campo de Dunas del Centro Pampeano. Los objetivos son caracterizar los procesos naturales que favorecen la acumulación, preservación y destrucción del registro faunístico moderno en ambientes lacustres salinos, evaluar los posibles sesgos en la representación taxonómica y anatómica de vertebrados y determinar el potencial de mezcla del registro arqueológico regional con huesos depositados naturalmente. Se realizaron transectas tafonómicas en las playas de las lagunas Las Tunas Grandes y Laguna Chica, que permitieron registrar la depositación natural de restos pertenecientes a una gran variedad de taxones de vertebrados de diversos tamaños. Los grupos taxonómicos más abundantes en Las Tunas Grandes son las aves acuáticas y los peces, mientras que en Laguna Chica predominan los mamíferos pequeños. Los resultados indican que la interacción de distintos factores biológicos, atmosféricos y geoquímicos, junto con la elevada dinámica litoral no favorece la preservación de los huesos. Si bien se registró la mezcla superficial de huesos depositados naturalmente con materiales arqueológicos, las posibilidades de que a partir del conjunto faunístico actual se forme un registro fósil serían excepcionales.
Palabras clave: Tafonomía regional; Campo de Dunas del Centro Pampeano; Lagunas salinas; Preservación ósea; Diversidad taxonómica.
Abstract
This paper presents the results of the first actualist taphonomic observations developed in two saline shallow lakes of the Hinojo-Las Tunas Lake System, located in the Central Pampean Dune Field. The objectives are to characterize the natural processes that favor the accumulation, preservation, and destruction of the modern faunal record in saline lacustrine environments, evaluate the possible biases in the taxonomic and anatomical vertebrate representation, and determine the potential for mixing of the regional archaeological record with naturally deposited bones. Taphonomic transects were carried out along the shoreline of Las Tunas Grandes and Laguna Chica shallow lakes, which allowed us to record the natural deposition of carcasses and bones belonging to a broad variety of vertebrate taxa of different sizes. The most abundant species are aquatic birds and fish in Las Tunas Grandes and small mammals in Laguna Chica. The results indicate that the interaction of different biological, atmospheric and geochemical factors, together with the high coastal dynamics do not favor bone preservation. Although the superficial mixing of naturally deposited bones with archaeological materials was recorded, the chances of a fossil record being formed from the current faunal assemblage would be exceptional.
Keywords: Regional taphonomy; Central Pampean Dune Field; Saline shallow lakes; Bone preservation; Taxonomic diversity.
Introducción
Las investigaciones arqueológicas sistemáticas desarrolladas en las últimas décadas en el Campo de Dunas del Centro Pampeano (CDCP) han permitido caracterizar los modos de vida de los grupos cazadores-recolectores que ocuparon esta área durante el Holoceno (Ávila, 2011; Scheifler y Messineo, 2016; Álvarez, 2018; Messineo et al., 2019; Scheifler, 2019; Álvarez et al., 2022). Se estudiaron materiales de estratigrafía y superficie (e.g., artefactos líticos, tiestos cerámicos, materiales faunísticos, restos humanos) procedentes de sitios a cielo abierto y se reconocieron complejos procesos de formación del registro arqueológico. Los sitios en estratigrafía se encuentran emplazados en dunas ubicadas en torno a lagunas de diferentes dimensiones y estabilidad. Por su parte, los sitios de superficie se distribuyen mayormente en las playas de las lagunas como consecuencia de la erosión hídrica de las dunas. Los resultados zooarqueológicos obtenidos hasta el momento muestran que los animales explotados durante el Holoceno fueron principalmente el guanaco (Lama guanicoe), el venado de las pampas (Ozotoceros bezoarticus), el ñandú (Rhea americana), el zorro pampeano (Lycalopex gymnocercus), la vizcacha (Lagostomus maximus), el zorrino (Conepatus chinga), los armadillos (Zaedyus pichiy, Chaetophractus villosus y Tolypeutes matacus), los roedores (Galea leucoblephara, Reithrodon auritus y Ctenomys sp.) y algunas especies de aves, como rapaces y tinámidos (Álvarez, 2018; Scheifler, 2019; Álvarez et al., 2022).
Hasta el presente no se habían desarrollado estudios actualistas sistemáticos en la región que sirviesen como marcos de referencia para evaluar los procesos de formación de los registros arqueológicos en términos de su resolución, integridad y preservación. Es necesario considerar que los conjuntos faunísticos representados en los sitios arqueológicos pueden estar conformados por una mezcla de huesos aportados por distintos agentes a lo largo de miles de años y a su vez, haber estados sometidos a procesos de destrucción diferencial (Beherensmeyer y Dechant Boaz, 1980; Binford, 1981; Borrero, 2001; Lyman, 2003; Miller et al., 2014; Gutiérrez et al., 2018). Los estudios tafonómicos que abordan la distribución natural de huesos en los ecosistemas actuales permiten identificar los múltiples procesos que afectan a los restos orgánicos y controlan su destrucción, acumulación y preservación (Kidwell y Flessa, 1995; Belardi y Carballo Marina, 2003; Behrensmeyer y Miller, 2012; Marchionni et al., 2020; Domingo et al., 2020), así como evaluar el impacto de la lluvia natural de huesos sobre el registro arqueológico (Borrero, 1989, 2001, 2014). Asimismo, este enfoque naturalista es una fuente de información útil para analizar las extinciones locales y reemplazos de especies, variaciones en las abundancias taxonómicas y amenazas actuales, lo que es relevante para contribuir a la biología de la conservación y el manejo de la vida silvestre (Kidwell y Flessa, 1995; Miller, 2011; Prassack, 2011; Massigoge et al., 2015).
Desde hace más de una década se están realizando estudios tafonómicos naturalistas sistemáticos en el sudeste de la región pampeana, basados principalmente en la recolección de datos mediante transectas tafonómicas en diferentes unidades ambientales (Massigoge et al., 2015; Gutiérrez et al., 2016, 2018; Álvarez et al., 2017; Scheifler et al., 2020). Al considerar la relevancia de la información que se obtuvo mediante estos estudios y el reciente desarrollo de investigaciones arqueológicas sistemáticas en el CDCP, se planteó la necesidad de iniciar una línea de tafonomía actualista en esta área que permita evaluar con mayor precisión los alcances y limitaciones de la información arqueológica y ecológica de los conjuntos faunísticos procedentes de sitios en estratigrafía y superficiales. Las características ambientales particulares del CDCP establecen la necesidad de generar información tafonómica naturalista específica, desde una perspectiva regional.
En este trabajo se presentan los primeros resultados de las investigaciones tafonómicas naturalistas desarrolladas en el CDCP. En esta oportunidad se focalizó en dos lagunas del Sistema Lagunar Hinojo-Las Tunas (SLHLT): Las Tunas Grandes (LTG) y Laguna Chica (LCH) (Figura 1). Los objetivos son: 1) caracterizar los procesos naturales que favorecen la acumulación, preservación y destrucción del registro faunístico moderno; 2) aportar al conocimiento tafonómico de vertebrados depositados en ambientes lacustres salinos, en particular a los sesgos potenciales en la representación taxonómica y anatómica; y 3) evaluar el potencial de contaminación y mezcla del registro arqueológico regional con huesos depositados naturalmente. Asimismo, se explora si las diferencias en las características de estas dos lagunas se traducen en rasgos distintivos de sus registros óseos.

Figura 1. Ubicación del Campo de Dunas del Centro Pampeano, el Sistema Lagunar Hinojo-Las Tunas y las localidades donde se realizaron las transectas tafonómicas.
Figure 1. Location of the Central Pampean Dune Field, the Hinojo-Las Tunas Shallow Lake System and the localities where the taphonomic transects were carried out.
Características Ambientales del Área de Estudio
El SLHLT (Figuras 1 y 2) se ubica en el centro de una amplia cubeta de escasa profundidad limitada por las cuencas intracratónicas de Macachín, Laboulaye y General Levalle, la cuenca tectónica del Salado y el Positivo Bonaerense (Zárate y Rabassa, 2005). La región estuvo sometida a una intensa actividad eólica durante periodos climáticos áridos-semiáridos del Pleistoceno tardío-Holoceno que alternaron con momentos de estabilidad geomorfológica durante lapsos de mayor humedad, lo cual ha conformado un paisaje de dunas longitudinales y parabólicas junto con lagunas desarrolladas en áreas interdunales que se encuentran en una unidad espacial que ha sido definida como Campo de Dunas del Centro Pampeano (Zárate y Tripaldi, 2012). Actualmente el paisaje se encuentra estabilizado como resultado de la fijación de médanos a través de la plantación de árboles y pastos durante el siglo XX (Taccari, 1948; Prego et al., 1965) y por el desarrollo de suelos a causa del aumento de la humedad en los últimos 40 años (Forte Lay et al., 2008).

Figura 2. a) Laguna Las Tunas Grandes (septiembre de 2021); b) Laguna Chica (septiembre de 2019); c) Restos óseos actuales de carpa asociados con plumas en Las Tunas Grandes; d) Tibia de vaca actual con sales en Laguna Chica; e) Restos óseos actuales de biguá en Las Tunas Grandes; f) Caparazón de mulita actual asociado con fecas de carnívoro en Laguna Chica.
Figure 2. a) Las Tunas Grandes shallow lake (September 2021); b) Laguna Chica (September 2019); c) Bone remains of modern carp associated with feathers in Las Tunas Grandes; d) Bone remains of modern cormorant in Las Tunas Grandes; e) Modern cow tibia with salts in Laguna Chica; f) Modern southern long-nosed armadillo carapace associated with carnivore feces in Laguna Chica.
El SLHLT está integrado principalmente por lagunas permanentes de grandes dimensiones concentradas en el paisaje, así como por lagunas de menor tamaño y espacialmente dispersas (Figura 1). La mayoría de las lagunas del sistema poseen aguas aportadas por el escurrimiento hídrico subterráneo regional y salinizadas a causa de la presencia de minerales solubles en los sedimentos (Kruse y Rojo, 1991). Análisis físico-químicos del agua de LTG dieron un pH de 9 y una baja alcalinidad, mientras que los de LCH dieron un pH de 9,3 y una alta alcalinidad (Ariel Grub Laboratorio Agrícola, 2022). El SLHLT es un sistema hidrológico arreico, pero en la actualidad, debido a la construcción de canales artificiales, constituye una de las zonas de derrame del río Quinto y de aporte al río Salado (Paoli y Giacosa, 2003). No obstante, las características (forma, tamaño, agregación y distribución, cuadro hidroquímico, etc.) de las lagunas son controladas principalmente por los aportes hídricos que provienen de las lluvias directas sobre el espejo de agua, el escurrimiento superficial y sobre todo del agua libre subterránea (Kruse y Rojo, 1991; López et al., 1991; Paoli y Giacosa, 2003). De este modo, se establece una gran dependencia e interrelación de estas masas de agua con los fenómenos hidrometeorológicos regionales (Dangavs, 2005). Datos provistos por el INTA Trenque Lauquen muestran precipitaciones medias anuales de 664 mm para el año 1995, que progresan hasta alcanzar los 1391 mm en el año 2001, cuando comienzan nuevamente a disminuir en el marco del ciclo hidrológico pampeano. Las temperaturas medias para el periodo 1960-2010 se ubican en un rango de 14,6 a 15,6 °C, con lo valores mas extremos durante los meses de otoño (marzo-abril) y primavera (octubre-noviembre) (Ferrelli y Aliaga, 2016).
La vegetación nativa se encuentra dominada por gramíneas como pasto colorado (Sorghastrum pellitum), paja amarga (Elionorus muticus), coirón (Poa lugalaris) y varias especies de pastos del género Stipa (Soriano et al., 1992). En menor medida se registran especies leñosas en forma aislada, como el chañar (Geofrea decorticans). En las orillas de las lagunas se desarrollan comunidades dominadas por espartillo (Spartina montividensis), pelo de chancho (Distichlis spicata), barba de indio (Chloris halophila) y hierbas latifoliadas como vinagrillo (Salicornia ambigua) y muérdago criollo (Limonium brasiliense).
Las comunidades faunísticas nativas del CDCP están dominadas por vertebrados pequeños, que incluyen roedores, marsupiales, aves y ofidios (Ringuelet, 1961). Los ungulados nativos como el guanaco (Lama guanicoe) y el venado de las pampas (Ozotoceros bezoarticus), abundantes en los sitios arqueológicos del Holoceno medio y tardío (Scheifler, 2019), se encuentran ausentes en el área de estudio en la actualidad. En cuanto a los peces, las primeras referencias para el área de especies como pejerrey argentino (Odontesthes bonaeriensis) y carpa común (Cyprinus carpio) se producen a principios de la década de 1990 (Kruse y Rojo, 1991; López et al., 1991). Su presencia en las lagunas del área podría responder a la siembra antrópica para pesca recreativa y comercial, a la dispersión por aves acuáticas o los desbordes del río Quinto (Kruse y Rojo, 1991; López et al., 1991). La carpa común fue introducida en Argentina a mediados del siglo XIX y su propagación en la región pampeana se produce desde la década de 1980, lo que estaría relacionado con el aumento del tamaño de las lagunas y de la interconexión de las mismas a causa del incremento de las precipitaciones y de las canalizaciones, lo que generó ambientes adecuados para el desove (Maiztegui et al., 2016). En el caso del pejerrey, comenzó a ser sembrado en lagunas del interior de esta región a partir de 1920 (Somoza et al., 2008).
Materiales y métodos
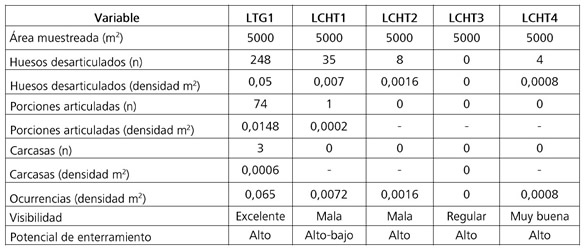
Se llevaron a cabo observaciones tafonómicas naturalistas en dos lagunas del SLHLT. En LTG, se realizaron cinco transectas de 500 m de longitud y 10 m de ancho. En el presente trabajo se analizaron los materiales de la transecta 1 (LTG1). En LCH se efectuaron cuatro transectas de iguales dimensiones y se analizó la totalidad de los restos recuperados. Las transectas fueron recorridas de a pie, siguiendo los bordes de las lagunas y prospectando la franja más próxima al agua. La posición inicial y final de cada una fue georreferenciada en el campo. Cada transecta se dividió en muestreos de 50 m y en cada uno de ellos se registraron las siguientes variables: tipo de sedimento; pendiente; potencial de enterramiento; tipo y distribución de la vegetación; bioturbación; presencia de animales vivos; actividad humana moderna; materiales arqueológicos; presencia de hallazgos óseos y visibilidad en función de la cubierta vegetal (excelente: sin cubierta vegetal; muy buena: 1-25%; buena: 26-50%; regular: 51-75%; y mala: 76-100% de cubierta vegetal). Cada hallazgo óseo se consideró como una ocurrencia que incluye restos desarticulados (un espécimen no relacionado con otro espécimen por el tejido blando); porciones articuladas (dos o más especímenes unidos por el tejido blando que comprenden menos del 75% del esqueleto animal); y carcasas (especímenes unidos por tejido blando que comprenden más del 75% del esqueleto animal).
Todas las ocurrencias faunísticas, con la excepción de aquellas de ungulados domésticos, se recolectaron para ser procesadas y analizadas en el laboratorio. Para cada una de ellas se relevaron las siguientes variables: taxón, unidad anatómica, presencia de tejido blando, estado de fusión, integridad de los elementos (completo [100% del elemento], pérdida ósea [95%], incompleto 50 a 95%], fragmento [< 50%]), articulación entre elementos, modificaciones tafonómicas, orientación y estado de enterramiento. En relación con esta última variable, siguiendo a Behrensmeyer y Dechant Boaz (1980), se consideró que una ocurrencia se encontraba enterrada cuando más del 50% de su superficie se hallaba cubierta por sedimentos. Además, para dar cuenta de aquellas ocurrencias parcialmente sepultadas, se utilizó la categoría de semienterrado, que es definida en este trabajo como aquellas ocurrencias cubiertas por sedimentos entre 1 y 49% de su superficie. El relevamiento de las variables tafonómicas se realizó utilizando al espécimen como unidad de análisis, es decir, en el caso de las porciones articuladas y carcasas, se analizó cada uno de los restos óseos que las integraban. Este análisis fue llevado a cabo en los huesos y se excluyeron los dientes aislados y placas dérmicas. Las variables tafonómicas registradas fueron estadio de meteorización, estadio de abrasión sedimentaria, presencia y tipo de marcas de carnívoros, marcas de roedores, de raíces, de pisoteo y manchas de manganeso (Behrensmeyer, 1978; Haynes, 1980; Binford, 1981; Shipman, 1981; Grayson, 1984; Olsen y Shipman, 1988; Lyman, 1994; Behrensmeyer et al., 2003; Gutiérrez y Kaufmann, 2007; Svoboda y Moreno, 2014). Además, se relevó la presencia de disolución química. En este trabajo esta variable es utilizada para designar a las alteraciones macroscópicas que se observan como una disolución del tejido cortical, en algunos casos con presencia de hoyos. Éstas no pueden ser aún asignadas a un agente o proceso particular.
El conjunto faunístico fue clasificado en las siguientes categorías: peces, aves, mamíferos medianos (1-20 kg) y mamíferos grandes (>20 kg). El Ntaxa se obtuvo contando el número de taxones identificados no superpuestos (Grayson, 1991). Se estimó el Número Mínimo de Individuos (NMI) teniendo en cuenta lateralidad y fusión, a partir del elemento óseo mejor representado (Lyman, 2008). Para el caso de las aves y los peces, los taxones más abundantes, se construyeron los perfiles de partes esqueléticas a partir del cálculo del Número Mínimo de Elementos (NME) y Unidades Anatómicas Mínimas (MAU y MAU%) (Binford, 1978; Klein y Cruz-Uribe, 1984; Lyman, 2008). En el caso de los caparazones de armadillo o fragmentos de estos, se realizó una observación tafonómica general a nivel de la porción, se contabilizaron las placas y se determinó si estaban completas o fragmentadas, así como si tenían presencia de alteración térmica.
Resultados
Laguna Las Tunas Grandes
La transecta LTG1 cubrió una superficie de 5000 m2 sobre una playa arenosa que poseía una visibilidad excelente y en donde predominaban los procesos de depositación lacustre. Dentro de esta transecta se observaron evidencias de actividades ganaderas y de pesca y se identificaron siete artefactos líticos distribuidos en tres muestreos. En cuanto al registro faunístico, se relevaron 248 restos desarticulados, 74 porciones articuladas y 3 carcasas. Estos restos se distribuían a lo largo de la playa y no se encontraban asociados con rasgos microambientales particulares (e.g., hoyadas). La densidad del total de las ocurrencias fue de 0,065/m2; la mayoría de ellas se hallaba en superficie (89,9%), aunque unas pocas se encontraban semienterradas (8,3%) y enterradas (1,9%) (Tabla 1).
Tabla 1. Principales variables registradas en las transectas.
Table 1. Main variables recorded in the transects.
El análisis taxonómico permitió obtener algún nivel de determinación sobre el 99,1% de las ocurrencias, a partir de las cuales se reconoció que el grupo taxonómico dominante fueron las aves (67,1%), seguido por los peces óseos (25,5%) y los mamíferos (6,5%) (Tabla 2). La representación taxonómica calculada a partir de los restos desarticulados muestra resultados similares, con una mayor proporción de aves (60,5%), en menor frecuencia peces (29,8%) y una representación marginal de mamíferos (8,5%). El patrón se acentúa al considerar las abundancias taxonómicas sobre la base de las porciones articuladas y las carcasas, las cuales pertenecen en su mayoría a aves (88,3%) y en menor medida a peces (11,7%). El cálculo del Ntaxa arrojó un valor de 17.
Tabla 2. Representación taxonómica de los hallazgos óseos registrados en las transectas analizadas. Referencias: RD= resto desarticulado; PA= porción articulada; CA= carcasa.
Table 2. Taxonomic representation in the analyzed transects. References: RD= disarticulated rest; PA= articulated portion; CA= carcass.

La avifauna se encuentra representada por nueve taxones (Tabla 2): gaviotas (Larus sp.), gaviotines (Sterna sp.), gallareta escudete rojo (Fulica rufifrons), gallareta chica (Fulica leucoptera), gallareta ligas rojas (Fulica armillata), cisne coscoroba (Coscoroba coscoroba), biguá (Phalacrocorax brasilianus), flamenco austral (Phoenicopterus chilensis) y gallineta común (Pardirallus sanguinolentus). Las especies más abundantes son la gallareta ligas rojas (Ocurrencias= 39, NME = 183, NMI = 7), el cisne coscoroba (Ocurrencias= 37, NME = 132, NMI = 6) y el biguá (Ocurrencias= 25, NME = 199, NMI = 6). Además, se identificaron 85 ocurrencias de aves que no pudieron ser determinadas a un nivel taxonómico específico; gran parte de estas ocurrencias (85%) son elementos no fusionados que, de acuerdo con su tamaño y caracteres morfológicos, podrían corresponder a individuos juveniles de flamenco austral. Los elementos no fusionados dan cuenta de un NMI de 19 y los fusionados de 1.
Los restos de peces que pudieron ser identificados pertenecen a las especies carpa (Cyprinus carpio) y pejerrey (Odontesthes bonaeriensis); asimismo, se determinaron unos pocos restos de los órdenes Perciformes (percas y chatas) y Siluriformes (bagres). Por último, los escasos restos de mamíferos corresponden a vaca (Bos taurus), oveja (Ovis aries), liebre europea (Lepus europaeus) y coipo (Myocastor coypus).
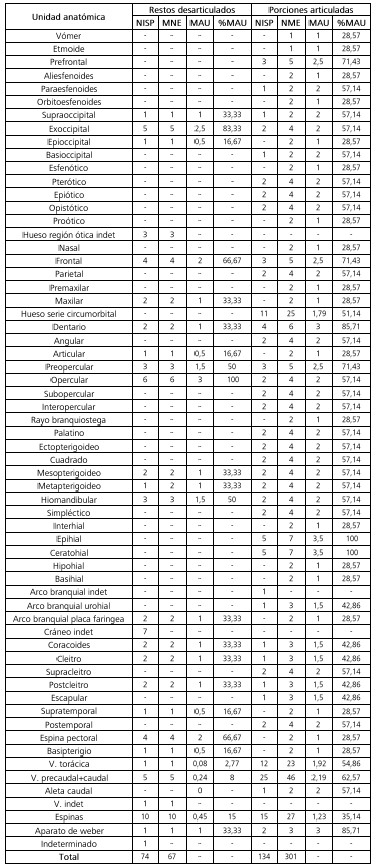
En lo que respecta a la representación de unidades anatómicas en las aves, se observan diferencias entre las porciones articuladas y carcasas, por un lado, y los restos desarticulados por el otro (Tabla 3; Figura 3a). En el primer caso, se registra la presencia de la mayoría de los elementos del esqueleto axial -craneal y postcraneal-, de las cinturas pélvica y escapular y de ambas extremidades. Los elementos más abundantes son el esternón, el coracoides, el fémur, el tibiotarso y la fíbula (MAU% > 67). Los menos representados son las vértebras torácicas y caudales, las costillas, la fúrcula, la patela y los huesos pequeños del autopodio como el metatarsiano 1, los carpianos y las falanges del ala y de la pata (MAU% < 33). Entre los restos desarticulados también se encuentran representadas todas las regiones anatómicas, aunque varios elementos están ausentes. El fémur es el elemento más abundante (MAU%= 100). Le siguen el cráneo, el sinsacro, el tibiotarso y el tarsometatarso en frecuencias moderadas (MAU%= 34-66). Los restantes elementos se registran en baja frecuencia (i.e., mandíbula, vértebras cervicales y torácicas, costillas, esternón, coracoides, húmero, ulna, carpometacarpo, patela, fíbula y falanges) o se encuentran ausentes (i.e., vértebras caudales, fúrcula, escápula, radio, carpianos y metatarsiano 1).
Tabla 3. Representación anatómica de los restos de aves registrados en la laguna Las Tunas Grandes. Referencias: V.= vértebra. *Especimen formado por dos vértebras torácicas fusionadas (parte del notarium).
Table 3. Anatomical representation of the birds of Las Tunas Grandes shallow lake. References: V.= vertebra. *Specimen formed by two fused thoracic vertebrae (part of the notarium).


Figura 3. Representación gráfica del perfil de partes esqueléticas de aves (a) y peces (b) recuperadas en la laguna Las Tunas Grandes.
Figure 3. Graphical representation of the profile of skeletal parts of birds (a) and fish (b) recovered in the Las Tunas Grandes shallow lake.
En cuanto a los peces, se identificaron tanto elementos del cráneo como del esqueleto axial y apendicular (Tabla 4, Figura 3b). No se registraron otolitos y elementos dorsales de la región axial. En las porciones articuladas y carcasas todas las regiones se encuentran bien representadas, tanto por la variedad de elementos identificados como por su abundancia (MAU% > 28). Los elementos mejor representados (MAU%= 67-100) son el prefrontal, el frontal, el dentario, el preopercular, el epihial, el ceratohial y el aparato de weber. Entre los restos no articulados, el cráneo se encuentra representado por escasos elementos, pero algunos de ellos, como el exoccipital y el opercular, poseen abundancias elevadas (> 67 MAU%). En cuanto al esqueleto apendicular, se encuentran presentes la mayoría de los elementos con frecuencias moderadas y bajas. Por último, los elementos del esqueleto axial se registran en muy baja frecuencia (Tabla 4, Figura 3b).
Tabla 4. Representación anatómica de los restos de peces registrados en la laguna Las Tunas Grandes. Referencias: V.= vértebra.
Table 4. Anatomical representation of fish from Las Tunas Grandes shallow lake. References: V.= vertebra.
El análisis tafonómico se llevó a cabo sobre un total de 1283 especímenes (Tabla 5, Figura 4). La mayor parte de estos pertenecen a aves (738 especímenes de porciones articuladas -PA- y carcasas -CA- y 150 restos desarticulados -RD-), seguidos por los peces (301 especímenes de porciones articuladas y carcasas y 74 restos desarticulados). Unos pocos corresponden a especímenes desarticulados de mamíferos medianos y grandes (5 y 15, respectivamente). Se registró una mayor diversidad y frecuencia de modificaciones tafonómicas entre los especímenes desarticulados. Entre los especímenes de aves, las modificaciones tafonómicas de la superficie cortical más comunes son las marcas de carnívoro (PA: 6,2%; RD: 10%), la abrasión sedimentaria (PA: 2,4%; RD: 14%), la disolución química (PA: 1,2%; RD: 10,7%) y la meteorización (PA: 0,1%; RD: 6,7%). Le siguen las tinciones por manganeso (4,7%, solo en RD) y las impresiones producidas por raíces (RD: 2%). En cuanto a los especímenes de peces, no se observan modificaciones superficiales entre aquellos que formaban parte de carcasas y porciones articuladas. Entre los desarticulados, se observa meteorización (13,5%), abrasión (5,4%) y disolución química (2,7%). Entre los especímenes correspondientes a mamíferos grandes (vaca y/o caballo) se observa un alto porcentaje de disolución química (60%), marcas de carnívoro (26,7%) y huellas de procesamiento antrópico (20%). En lo que refiere a estas últimas, se registró una costilla con huellas de corte producidas posiblemente por un filo metálico y marcas de aserrado, así como dos vértebras aserradas. También se registraron marcas de raíces y abrasión (6,7% en ambos casos). Además, dos especímenes se encontraban meteorizados, uno en el estadio 3 y otro en el estadio 4. Por su parte, entre los especímenes de mamíferos medianos, se observó un caso con disolución química, uno con abrasión y otro con meteorización (estadio 1).
Tabla 5. Variables tafonómicas relevadas. Referencias: RD=resto desarticulado; RA= porción articulada; Mam.= mamífero; Med.= mediano; Gran.= grande.
Table 5. Taphonomic variables surveyed. References: DR= disarticulated remain; RA= articulated portion; Mam.= mammalian; Med.= medium; Gran.= large.


Figura 4. Ejemplos de modificaciones tafonómicas registradas en las transectas. a) Cráneo de hurón menor de Laguna Chica con disolución química, b) Tibiotarso de coscoroba con disolución química de la laguna Las Tunas Grandes, c) Porción articulada de esternón y costilla de coscoroba de la laguna Las Tunas Grandes con marcas de carnívoros, d) Escápula de oveja de Laguna Chica con marcas de carnívoros. Escalas= 1 cm.
Figure 4. Examples of taphonomic modifications recorded in the transects. a) Skull of lesser ferret from Chica shallow lake with chemical dissolution, b) Tibiotarsus of coscoroba with chemical dissolution from Las Tunas Grandes shallow lake, c) Articulated portion of sternum and rib of coscoroba from Las Tunas Grandes shallow lake with carnivore marks, d) Scapula of sheep from Chica shallow lake with carnivore marks. Scales= 1 cm.
Con respecto a los patrones de fragmentación, en las aves, el 5,4% de los especímenes articuladas y el 11,3% de los especímenes desarticulados están fragmentados. En los peces, solo se registró fragmentación entre los especímenes desarticulados (9,5%). En lo que refiere a los mamíferos, el 60% de los especímenes de taxones grandes y el 80% de los medianos se encuentran fragmentados.
Laguna Chica
Las cuatro transectas realizadas en LCH cubrieron una superficie de 20.000 m2. Se realizaron sobre la playa arenosa de la laguna, la cual presentaba sectores con pastizales. Debido a los cambios en la vegetación, la visibilidad era variable: en LCH1 era en mayor medida mala, con sectores restringidos de visibilidad buena y muy buena; en LCH2 y LCH3 era predominantemente mala y regular; y en LCH4 era en general muy buena. En las primeras tres transectas se destacaron los procesos de pedogénesis y depositación lacustre, mientras que en LCH4 se registraron procesos de depositación sobre la playa y de erosión sobre la barranca de un médano. En todas las transectas se observaron evidencias de actividad ganadera y en una de ellas se registraron desechos antrópicos recientes. En las transectas LCH1 y LCH4 se identificaron artefactos líticos dispersos (uno en el primer caso y seis en el segundo). En cuanto a las ocurrencias faunísticas, se registraron 47 restos desarticulados y 1 porción articulada (Tabla 1). Al igual que en LTG1, los hallazgos faunísticos se distribuían a lo largo de la playa y no se encontraban asociados con rasgos microambientales particulares. La densidad del total de las ocurrencias fue de 0,0024/m2 y todas ellas se encontraban en superficie. Además, se relevaron 194 placas dérmicas y 12 fragmentos de coraza de armadillos.
El análisis taxonómico permitió obtener algún nivel de determinación sobre la totalidad de las ocurrencias faunísticas (Tabla 2). Predominan ampliamente los restos de mamíferos grandes (71%), seguidos por los de mamíferos medianos (27%) y en mucha menor medida de aves (2%). Entre los mamíferos se identificó la presencia de vaca, oveja, coipo, peludo (Chaetophractus villosus), mulita (Dasypus hybridus), zorrino (Conepatus chinga) y hurón menor (Galictis cuja). Con respecto a otras clases taxonómicas, solo se identificó un tibiotarso de cisne coscoroba (Tabla 2). El Ntaxa de esta localidad es de ocho.
El análisis tafonómico (Tabla 5, Figura 4) de los especímenes de mamíferos grandes muestra que la modificación más frecuente de la superficie cortical es la meteorización (56%); la mayoría de los restos se encuentran en el estadio 1, aunque también se registraron restos en los estadios 2, 3 y 4. Otra alteración que se registra con frecuencia es la disolución química (29,4%). Además, en tres especímenes se observaron marcas de carnívoro (8,8%). En el caso de los especímenes de mamíferos medianos, un alto porcentaje exhibe disolución química (63,6%). Las marcas de carnívoro y la meteorización -estadios 1 y 2- también son frecuentes (36,4% en cada caso). Entre los especímenes de mamíferos medianos de las porciones articuladas no se observaron efectos tafonómicos. El único espécimen de ave se encuentra meteorizado en estadio 1 y posee marcas de raíces y disolución química. Por otra parte, se destaca que gran parte de los restos faunísticos de esta localidad presentaban depositación de sales, aunque este atributo no fue registrado de modo sistemático.
Con respecto a la fragmentación, esta afectó a un 38,2% de los especímenes desarticulados de mamíferos grandes y a un 18,2% de los mamíferos medianos. Por otro lado, si bien los elementos de la coraza de armadillo no fueron incluidos en el análisis tafonómico, es importante mencionar que se observaron fracturas probablemente producidas por carnívoro en una coraza de peludo y en varios fragmentos de coraza de mulita (estos últimos posiblemente de un mismo individuo). En ambos casos, se hallaron fecas de carnívoro asociadas con estas ocurrencias (Figura 2).
Discusión
El primer trabajo tafonómico naturalista llevado a cabo en el CDCP muestra importantes diferencias en los registros faunísticos entre distintos ambientes lagunares. La cantidad de restos, así como la riqueza taxonómica es comparativamente menor en LCH (Densidad total m2= 0,0024, Ntaxa= 8) en relación con LTG (Densidad total m2= 0,065, Ntaxa= 17) a pesar de que el área analizada en la primera localidad comprende mayor superficie. Si bien se considera que la menor visibilidad de LCH con respecto a LTG pudo contribuir al contraste del registro faunístico de ambas lagunas, se plantea que éste podría obedecer en gran parte a las diferencias en las características bióticas de cada laguna, aspecto que se discutirá más adelante. En LCH gran parte de los restos pertenecen a mamíferos medianos y grandes, que se presentaban en su totalidad en superficie y en su mayoría desarticulados, con evidencia de meteorización. Por el contrario, las características del conjunto faunístico de LTG son diferentes, ya que se registró la presencia de nueve especies de aves acuáticas y de al menos cuatro tipos de peces. Ambas clases de vertebrados se encontraban representadas tanto por restos desarticulados como por porciones articuladas o carcasas. La mayoría de las ocurrencias se hallaban en superficie; sin embargo, algunas de ellas estaban en proceso de incorporación al depósito sedimentario de la playa.
Los marcados contrastes que se observan en los registros faunísticos de los ambientes lacustres del SLHLT responderían principalmente a las diferencias limnológicas particulares de las lagunas estudiadas. LTG constituye un cuerpo lacustre estable, que reúne condiciones favorables de hábitat para las aves acuáticas, entre las que se destacan las propiedades físico-químicas del agua, la disponibilidad y distribución de alimento, así como la oferta de lugares adecuados para la reproducción y descanso de las aves. Por el contrario, el tamaño del cuerpo de agua de LCH la hace más sensible al régimen pluvial, llegando a secarse durante períodos de déficit hídrico. Durante los trabajos de campo realizados en LCH, se observaron amplias variaciones diarias y estacionales en la superficie y posición del cuerpo de agua, que se relacionarían con los cambios en la precipitación y en la dirección de los vientos. Estas variaciones influyen en las características físicas, químicas y biológicas de este ambiente de humedal, lo cual condiciona la abundancia de aves (Wiens, 1989).
La comunidad biótica de las lagunas está limitada principalmente por la concentración de nutrientes (Grosman, 2008). LTG es una laguna habilitada para la pesca comercial de pejerrey (Berasain y Argemi, 2007). En cambio, LCH, no ha sustentado hasta el presente una comunidad de peces, posiblemente por sus características limnológicas ya descritas. Estas diferencias se relacionarían con la capacidad productiva orgánica de ambas lagunas, lo que sugiere una riqueza biótica superior en LTG y, en consecuencia, la mayor abundancia y diversidad del registro faunístico de esta última laguna, en especial de las aves y peces.
A partir de los análisis taxonómicos, los restos de peces de LTG1 fueron asignados a dos especies (carpa y pejerrey) y dos órdenes (Perciformes y Siluriformes). La especie más abundante es la carpa. Los relevamientos ictiológicos realizados en el SLHLT a través de diferentes técnicas de captura (enmalles, trampas, ranio) señalan la obtención de pejerrey, carpa, bagre sapo (Rhamdia quelen), tosquero (Jenynsia lineata), mojarra (Characidae),dientudo (Oligosarcus jenynsii) y sabalito (Cyphocharanx voga) (Berasain y Argemi, 2007). La diferencia entre lo relevado en las transectas y el informe del Departamento de Desarrollo y Tecnología Pesquera puede deberse en parte a que los esqueletos de los peces pequeños (e.g., mojarra, tosquero) suelen depositarse en los fondos de los cuerpos de agua, mientras que los restos de peces de mayor tamaño (e.g., carpa) son movidos por las corrientes a la costa (Zohar et al., 2008). Asimismo, en el caso de que restos de especies más pequeñas llegaran a la playa, seguramente se destruirían/desaparecerían más fácilmente que los de las especies más grandes. Además, estos informes destacan que el pejerrey es la especie dominante en LTG y que está sometido a una elevada presión de pesca comercial. Las diferencias en la diversidad taxonómica de la comunidad ictícola viviente y la muestra esqueletaria analizada también pueden explicarse en parte, por las limitaciones metodológicas en la recuperación de los restos de peces más pequeños. Sin embargo, se ha señalado que, dada la tendencia del aumento constante de la salinidad a partir del año 2004, la población de carpas se encuentra sometida a un fuerte estrés ambiental que posiblemente pueda generar grandes mortandades naturales (Berasain y Argemi, 2007).
En LTG se determinaron restos esqueléticos de al menos nueve taxones de aves acuáticas. Entre éstas, la gallareta ligas rojas, el cisne coscoroba y el biguá son las que poseen mayor abundancia. En el CDCP los estudios actuales sobre la avifauna se restringen a especies que habitan ambientes terrestres (Codesido et al., 2012). Por este motivo, no se cuenta con información detallada de la riqueza y diversidad actual de aves acuáticas que permitan evaluar la fidelidad del conjunto muerto. Sin embargo, durante la realización de las transectas se observaron en las inmediaciones ejemplares vivos de 17 especies de aves: flamenco austral, gallareta chica, gallareta escudete rojo, gallareta ligas rojas, biguá, gaviota cocinera (Larus dominicanus), gaviota capucho gris (Larus cirrocephalus), gaviotín lagunero (Sterna trudeaui), gallineta común, tero (Vanellus chilensis), tero real (Himantopus melanurus), chimango (Phalcoboenus chimango), coscoroba, pato cuchara (Anas platalea), pato maicero (Anas georgica), pato gargantilla (Anas bahemensis) y pato zambullidor chico (Oxyura vittata). De acuerdo con estos datos, la muestra esquelética analizada hasta el momento daría cuenta solo parcialmente de la riqueza de aves que caracteriza a este ambiente lacustre, diversidad que podría aumentar cuando se amplíe el área estudiada.
Una característica destacable del registro de LTG es la abundancia de elementos de aves no fusionados que podrían corresponder a individuos subadultos de flamenco austral. Esto, sumado a las observaciones de individuos juveniles realizadas en abril de 2014 (Scheifler, 2019), sugiere que la laguna funciona como un lugar de anidamiento de esta especie. Hasta el momento no se ha observado la presencia de nidos de flamencos, lo cual puede deberse a que las visitas al campo se realizaron por fuera de la temporada reproductiva (primavera/verano, Petracci et al., 2020). El flamenco austral nidifica preferentemente en lagunas de agua salada (de la Peña, 1987). Chiale y coautores (2021) mencionan que, para la provincia de Buenos Aires, el sector de las lagunas encadenadas (aproximadamente a 100 km del SLHLT) constituye una de las áreas reproductivas más importantes de la especie.
Al comparar los conjuntos faunísticos de ambas lagunas, se observan algunas similitudes en los tipos de modificaciones tafonómicas de los restos, aunque no en sus abundancias. Las evidencias de la acción de carnívoros son frecuentes en los restos de mamíferos grandes en ambas lagunas, así como en los restos de mamíferos medianos de LCH y en los de aves de LTG. Las márgenes de lagunas son importantes sitios de caza y/o carroñeo para diferentes depredadores, debido a la disponibilidad natural de animales. El carroñeo pudo haber contribuido a la dispersión y destrucción de elementos esqueléticos de animales muertos por distintas causas (Bickart, 1984; Oliver y Graham, 1994; Rodríguez-Hidalgo et al., 2016). En el caso particular de las aves se considera además que, tanto carnívoros mamíferos como aves rapaces, también pudieron haber tenido un rol destacado en su muerte, especialmente en el caso de los individuos más jóvenes. El accionar de los carnívoros podría explicar parcialmente la representación desigual de unidades anatómicas en los perfiles de partes esqueléticas del grupo de las aves, donde se observa una menor frecuencia del esqueleto axial y extremidades anteriores (Rodríguez-Hidalgo et al., 2016). Sin embargo, numerosos estudios actualistas muestran que los patrones de supervivencia en aves dependen de la estructura física y densidad mineral de los elementos, la cual varía en relación con la anatomía funcional de cada grupo taxonómico (e.g., Livingston, 1989; Bovy, 2002; Cruz, 2005). En futuros trabajos se explorarán los perfiles de partes esqueléticas clasificando a las aves según su anatomía funcional.
En el caso de los restos de peces, abundantes en LTG, no se identificaron modificaciones producidas por carnívoros. Una muy baja incidencia de carnívoros (< 1%) fue informada por Stewart (1991) para un conjunto de huesos modernos localizados en la playa del Lago Turkana (África), a pesar de la abundancia de carnívoros en el entorno local. Tampoco se registraron modificaciones producidas por procesamiento antrópico (e.g., huellas de corte y alteración térmica), que indiquen que fueron descartados por pescadores modernos. En la muestra de elementos de porciones articuladas y carcasas, todas las regiones anatómicas se encuentran bien representadas, mientras que en el conjunto de huesos desarticulados el esqueleto axial está subrepresentado. Estudios actualistas muestran que en general, las acumulaciones naturales de peces exhiben una representación relativamente uniforme de elementos craneales y postcraneales (Stewart, 1991; Butler, 1993; Corbat, 2014). A pesar de la ausencia de modificaciones por carnívoros en los restos de peces, se considera que la baja representación de los elementos axiales puede deberse en parte a su ingestión o transporte para posterior consumo por este agente. Zangrando (2003: 47) ha observado que los zorros consumen la porción axial de los cuerpos de peces, sin producir modificaciones en los cráneos. Además, debe tenerse en cuenta el transporte diferencial de partes esqueletales por acción hídrica, ya que se ha demostrado que las vértebras son más susceptibles a este tipo de procesos (Corbat et al., 2017). Asimismo, estos elementos podrían enterrarse más fácilmente, dado su pequeño tamaño y la naturaleza arenosa del sustrato. A pesar de la ausencia de huellas de procesamiento, no se puede descartar la posibilidad de que el perfil de partes esqueléticas exprese también un descarte selectivo por pescadores actuales.
Otro tipo de modificación común en ambas lagunas es la presencia de disolución química, la cual se registra particularmente entre los restos desarticulados de mamíferos medianos y grandes. El origen de este tipo de alteración no se ha podido establecer hasta el momento, pero se considera que puede estar relacionado con las características del ambiente de depositación, en particular la naturaleza salina del medio acuoso y el régimen hídrico imperante en la franja litoral donde se realizaron las transectas. Ese sector de las lagunas se encuentra sometido a eventos de inundación recurrentes seguidos por episodios de desecación, como consecuencia de las fluctuaciones estacionales en el tamaño del cuerpo lagunar e incluso diarias por cambios en la dinámica del oleaje. La presencia de agua en el ambiente donde se encuentran depositados los huesos es uno de los principales factores que condicionan su preservación y/o modificación (Hedges y Millard, 1995; Trueman et al., 2004; Kendall et al., 2018). Los ambientes fluctuantes, en los cuales se alternan condiciones de humedad y sequedad, son los más perjudiciales para los huesos ya que provocan que estos se hinchen y se encojan estableciendo tensiones físicas que dan lugar a grietas y exfoliación (Behrensmeyer, 1978; Conard et al., 2008). Si el medio acuoso además tiene alta salinidad, este proceso puede generar cristalización de sales sobre la superficie y al interior del hueso, lo cual acentúa estas tensiones y aumenta su alteración (Behrensmeyer, 1978; Prassack, 2011). Por otro lado, estudios experimentales indican que los suelos con valores de pH mayores a 7,5 afectan principalmente al colágeno del hueso, no tanto al componente mineral (Kendall et al., 2018). Sin embargo, debido a que ambos componentes se encuentran íntimamente relacionados, es posible que la rápida degradación del colágeno, en un ambiente altamente alcalino como LCH y con un alto régimen hídrico, pueda favorecer también la disolución de la hidroxiapatita debido a equilibrios del carbono inorgánico en el ecosistema lagunar (Manahan, 2007). Además de la degradación físico-química arriba mencionada, en estos ambientes acuáticos también es esperable que los huesos sufran bioerosión causada por distintos microorganismos (Kendall et al., 2018). Por el momento, no es posible afirmar que el deterioro químico observado en los huesos de LTG y LCH se deba a alguno de estos procesos en particular, aunque lo más probable es que obedezca a la acción combinada de varios de ellos. Se considera que los procesos característicos de estos microambientes inmediatos de depositación contribuyen significativamente a la destrucción ósea.
En el caso de la meteorización, se observan altos porcentajes para mamíferos grandes y medianos en LCH. En LTG los porcentajes son menores y se registran para mamíferos grandes (≥ 20 kg), peces y, en menor medida, aves. Asimismo, en LTG, los restos de peces y aves presentan un mejor estado de preservación en comparación con los de mamíferos grandes, lo cual se evidencia en el registro más frecuente de elementos articulados y en la mayor completitud de los restos desarticulados.
En términos generales, al comparar entre grupos taxonómicos se observa una mayor incidencia de los efectos de la meteorización y disolución química en los restos de mamíferos grandes y medianos. Estas variables también ayudarían a explicar la mayor fragmentación observada para este grupo taxonómico. Esta variabilidad en las modificaciones tafonómicas muestra que los conjuntos de mamíferos representan tiempos más prolongados de exposición que los conjuntos de aves y peces. Como ya han indicado otros autores, esta diferencia es esperable dadas las propiedades óseas intrínsecas de cada grupo taxonómico, como por ejemplo tamaño y estructura ósea (Behrensmeyer, 1978; Behrensmeyer et al., 2003; Cruz, 2003).
De acuerdo con observaciones del ambiente de depositación efectuadas durante la realización de los trabajos de campo, las chances de entierro de los restos esqueléticos son mayores en LTG, dada la alta movilización de arena por el oleaje. Esto es respaldado por los datos de estado de entierro, que muestran que aproximadamente un 8% de las ocurrencias de esta laguna se encontraban semienterradas y un 2% enterradas. No obstante, a causa de la alta dinámica litoral, los restos que se entierran pueden ser re-expuestos por la erosión hídrica, como lo evidencia la presencia de algunos elementos de aves con impronta de raíces.
De continuar las condiciones ambientales actuales, la alta tasa de reciclado de huesos en superficie, limitarían las posibilidades de que el conjunto faunístico actual se preserve y dé lugar a un registro fósil. De preservarse parte de este conjunto, se esperaría que el registro resultante esté formado por una baja frecuencia de restos óseos con un sesgo marcado a favor de los vertebrados más grandes y de los elementos anatómicos más resistentes. Alternativamente, este conjunto faunístico podría preservarse si ocurriese un evento transgresivo rápido de la laguna por un aumento significativo de la humedad a nivel regional. En esa situación, el sector litoral actual pasaría a estar sumergido, por lo cual los restos esqueléticos dejarían de estar expuestos a los procesos destructivos y comenzarían a ser sepultados por la sedimentación lacustre. El registro resultante será una acumulación atricional con patrones de preservación diferencial y tiempos de promediamiento diferentes para los distintos grupos taxonómicos.
Este primer estudio desde la tafonomía actualista sugiere que el potencial de mezcla del registro arqueológico regional con huesos depositados naturalmente varía en relación con las características de las lagunas, tanto en la diversidad taxonómica como en la densidad de restos. En ambas lagunas se registraron asociaciones entre artefactos líticos y huesos actuales. Si bien distintos procesos tafonómicos conducirían a una elevada tasa de destrucción de los restos faunísticos, el uso continuo y recurrente de este espacio por una gran diversidad de aves y la existencia de peces, garantiza un aporte continuo de nuevos huesos. En consecuencia, los conjuntos óseos superficiales en sectores de playa de las lagunas representan, en la mayoría de los casos, depositaciones muy recientes. Sin embargo, el hallazgo de restos óseos con precipitaciones de carbonatos y marcas de raíces podría sugerir una mayor antigüedad, ya que se trata de procesos que involucran un considerable tiempo de formación.
En contextos arqueológicos enterrados la mezcla con huesos modernos puede producirse por migración vertical. Este mecanismo se vería favorecido por el pisoteo sobre el sustrato arenoso, la acción de raíces y la acción de animales cavadores (Borrero, 1988; Lyman, 1994). En distintos sitios arqueológicos del Holoceno medio y tardío del CDCP (e.g., La Susana 1, Laguna Cabeza de Buey y Laguna Chica 1) se ha registrado la presencia de restos de peces y aves acuáticas que por sus características tafonómicas (coloración, textura, preservación, entre otros) han sido interpretados como resultado de la migración de huesos modernos (Scheifler y Messineo, 2016; Scheifler et al., 2022).
Dadas las expectativas de mezcla arriba señaladas, el conocimiento sobre la distribución de las diferentes especies, la presencia de taxones introducidos históricamente, las características tafonómicas de los huesos y la identificación de procesos de incorporación eto-ecológicos son claves para evaluar la integridad de los registros fósiles en los diferentes momentos cronológicos del CDCP.
Conclusiones
Este trabajo constituye un estudio tafonómico actualista pionero en el centro pampeano. El análisis de los restos óseos actuales depositados naturalmente en las playas de las lagunas salinas del área permite discutir diversos aspectos de la ecología moderna de estos cuerpos de agua y generar implicancias para interpretar el registro faunístico de los sitios arqueológicos. Los dos cuerpos de agua estudiados poseen características limnológicas diferenciales que se reflejan en la producción biológica y sus aspectos ecológicos, traducidos en la diversidad taxonómica y la abundancia de individuos por especie. LTG reúne condiciones favorables de hábitat que permite sostener una mayor riqueza biótica que LCH. Así, en la primera de ellas, las aves acuáticas y los peces encuentran un ambiente propicio para habitar y reproducirse, tal como se observa en los conjuntos óseos modernos estudiados. En general, las condiciones químicas del agua y del suelo de estas lagunas, entre las que se destacan el pH básico y la alta salinidad, en combinación con otros fenómenos meteorológicos y biológicos, no favorecerían la preservación ósea a largo plazo. Los conjuntos óseos actuales constituyen una imagen parcial de la ecología de los últimos años, con una incorporación y destrucción constante de organismos y sesgada en detrimento de los taxones más pequeños.
Las lagunas salinas están sujetas a complejas dinámicas ecológicas y ambientales que las convierten en ecosistemas inestables. En consecuencia, los estudios a corto plazo posiblemente no captan la variabilidad ecológica y su correlato en la distribución natural de huesos necesarios para la construcción de modelos tafonómicos generales. Sin embargo, este trabajo sienta las bases para entender la complejidad ecológica y de preservación que da lugar a la formación de los depósitos óseos. Bajo este tipo de ambientes, resulta necesario llevar a cabo estudios ecológicos y tafonómicos a largo plazo, los cuales permitirán evaluar la dinámica particular de estos sistemas y generar información confiable para realizar interpretaciones del pasado, así como elaborar planes de manejo y conservación de especies. Comprender en qué medida estos depósitos se ven transformados con el paso del tiempo requiere de estudios específicos, como experimentaciones controladas, observaciones naturalistas longitudinales y excavaciones de conjuntos naturales enterrados, lo que permitirá enriquecer estas interpretaciones.
Olavarría, 15 de junio de 2022
Agradecimientos
Los autores agradecen a los organizadores del II Taller de Tafonomía Actualística: Metodología y Aplicaciones Arqueológicas, Julián Mignino, Paula Weihmüller y José Manuel López. También agradecen a los amigos de Trenque Lauquen, Ariel Grub, Juanjo Estevez, Vicente Benito y José Zucardi, por su colaboración para la realización de los trabajos de campo; a Paula Vitale por sus aportes a la discusión de los procesos químicos de las lagunas; a Luciana Stoessel por sus contribuciones al análisis de la muestra de peces y a Atilio F. Zangrando por atender las consultas realizadas. Por último, a los evaluadores anónimos que permitieron, con sus correcciones y sugerencias, mejorar el trabajo original. Estas investigaciones se realizaron con recursos económicos de los siguientes proyectos e instituciones: PICT 2018-0686, PICT 2019-2750, Instituto de Investigaciones Arqueológicas y Paleontológicas del Cuaternario Pampeano (INCUAPA, UNCPBA-CONICET), Municipalidad de Trenque Lauquen y Facultad de Ciencias Sociales (FACSO-UNCPBA) (Convenio de Cooperación, Ordenanza Municipal Nº 3629/10). Las interpretaciones aquí expresadas son de exclusiva responsabilidad de los autores.
1. Álvarez, M.C. (2018). Análisis faunístico del sitio Laguna de los Pampas (partido de Lincoln, Región Pampeana). Aportes a los estudios de la subsistencia en el área Oeste de la Pampa Húmeda. Arqueología, 24(1), 127-146. https://doi.org/10.34096/arqueologia.t24.n1.4229 [ Links ]
2. Álvarez, M.C., Massigoge, A., Scheifler, N.A., González, M.E., Kaufmann, C.A., Gutiérrez, M.A. y Rafuse, D.J. (2017). Taphonomic effects of a grassland fire on a modern faunal sample and its implications for the archaeological record. Journal of Taphonomy, 15(1-3), 77-90. https://journaltaphonomy.com/tag/grassland-fire/ [ Links ]
3. Álvarez, M.C., Santos Valero, F., Borges Vaz, E. y Messineo, P.G. (2022). Ocupaciones de cazadores-recolectores en el Campo de Dunas del Centro Pampeano durante el Holoceno Tardío. El Caso de la Localidad Arqueológica Laguna Giaconne (Buenos Aires, Argentina). Comechingonia. En Prensa. [ Links ]
4. Ávila, J.D. (2011). Resultados de los fechados radiocarbónicos del sitio Laguna El Doce, departamento General López, provincia de Santa Fe. Relaciones de la Sociedad Argentina de Antropología, XXXVI, 337-343. [ Links ]
5. Ariel Grub Laboratorio Agrícola (2022). Informe Físico-químico del agua. [ Links ]
6. Behrensmeyer, A.K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology, 4, 150-162. https://doi.org/10.2307/2400283 [ Links ]
7. Behrensmeyer, A.K. y Dechant Boaz, D.E. (1980). The recent bones of Amboseli Park, Kenya, in relation to East African paleoecology. En Behrensmeyer, A.K. y Hill, A.P. (Eds.). Fossils in the making, pp. 72-92. Chicago: University of Chicago Press. [ Links ]
8. Behrensmeyer, A.K. y Kidwell, S.M. (1985). Taphonomy’s contribution to Paleobiology. Paleobiology, 11, 105-119. https://www.jstor.org/stable/2400427
9. Behrensmeyer, A.K., Kidwell, S.M. y Gastaldo, R.A. (2000). Taphonomy and paleobiology. En Erwin, D.H. y Wing, S.L. (Eds). Deep time: paleobiology’s perspective, pp. 103-147. Lawrence: The Paleontological Society.
10. Behrensmeyer, A.K., Stayton, C.T. y Chapman R.E. (2003). Taphonomy and ecology of modern avifaunal remains from Amboseli Park, Kenya. Paleobiology, 29, 52-70. https://www.jstor.org/stable/4096874 [ Links ]
11. Behrensmeyer, A.K. y Miller, J.H. (2012). Building links between Ecology and Paleontology using taphonomic studies of recent vertebrate communities. En Louys, J., (Ed.). Paleontology in Ecology and Conservation, pp. 69-91. Berlin Heidelberg: Springer-Verlag. [ Links ]
12. Belardi, J.B. y Carballo Marina, F.C. (2003). Tafonomía regional en la cuenca media del río Coyle (Santa Cruz, Patagonia Argentina). Intersecciones en Antropología, 4, 59-73. http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S1850-373X2003000100005 [ Links ]
13. Berasain, G. E. y Argemi, F. (2007). Laguna las Tunas. Partido de Trenque Lauquen. Campaña de relevamientos limnológicos e ictiológicos. Ministerio de asuntos agrarios. Dirección de desarrollo pesquero. Informe técnico N 94 .https://www.maa.gba.gov.ar/pesca1/images/archivos/informes_biologicos/continentales/94_tunas07.pdf [ Links ]
14. Bickart, K.J. (1984). A field experiment in avian taphonomy. Journal of Vertebrate Paleontology, 4:4, 525-535. http://dx.doi.org/10.1080/02724634.1984.10012028 [ Links ]
15. Binford, L.R. (1978). Nunamiut Ethnoarchaeology. Nueva York: Academic Press New York. [ Links ]
16. Binford, L.R. (1981). Bones: ancient men and modern myths. Nueva York: Academic Press New York. [ Links ]
17. Blumenschine, R.J. (1988). An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages. Journal of Archaeological Science, 15, 483-502. [ Links ]
18. Borrero, L.A. (1988). Estudios tafonómicos en Tierra del Fuego: su relevancia para entender procesos de formación del registro arqueológico. En Yacobaccio, H.D. (Ed). Arqueología Contemporánea Argentina, pp. 13-32. Buenos Aires: Ediciones Búsqueda. [ Links ]
19. Borrero, L.A. (1989). Sites in action: the meaning of guanaco bones in Fueguian archaeological sites. ArchaeoZoologia, III, 9-24. [ Links ]
20. Borrero, L.A. (2001). Regional taphonomy: background noise and the integrity of the archaeological record. En Kuznar, L.A. (Ed.). Ethnoarchaeology of Andean South America. Contributions to archaeological method and theory, pp. 243-254. Ann Arbor: International Monographs in Prehistory. [ Links ]
21. Borrero, L.A. (2014). Taphonomy, regional. En Smith, C. (Ed.). Encyclopedia of global archaeology, SE-832, pp 7232-7235. Nueva York: Springer. [ Links ]
22. Borrero, L.A. y Martin, F.M. (1996). Tafonomía de carnívoros: un enfoque regional. En Gómez Otero, J. (Ed.). Arqueología. Sólo Patagonia, pp. 189-198. Puerto Madryn: CENPAT-CONICET. [ Links ]
23. Bovy, K.M. (2002). Differential avian skeletal part distribution: explaining the abundance of wings. Journal of Archaeological Science, 29, 965-978. https://doi.org/10.1006/jasc.2001.0795 [ Links ]
24. Butler, V. (1993). Natural Versus Cultural Salmonid Remains: Origin of The Dalles Roadcut Bones, Columbia River, Oregon, U.S.A. Journal of Archaeological Science, 20, 1-24. https://doi.org/10.1006/jasc.1993.1001 [ Links ]
25. Chiale, M.C., L. Pagano y S. Imberti (2021). Situación reproductiva del flamenco austral (Phoenicpterus chilensis) en el sur de su distribución: implicancias para su conservación. El Hornero, 36(2), 131-137. [ Links ]
26. Codesido, M., González-Fischer, C. y Bilenca, D. (2012). Agricultural land-use, avian nesting and rarity in the Pampas of central Argentina. Emu, 112, 46-54. http://dx.doi.org/10.1071/MU11049 [ Links ]
27. Conrad, N.J., Walker, S.J. y Kandel, A.W. (2008). How heating and cooling and wetting and drying can destroy dense faunal elements and lead to differential preservation. Palaeogeography, Palaeoclimatology, Palaeoecology, 266, 236–245. doi: 10.1016/j.palaeo.2008.03.036
28. Corbat, M. (2014). Una Aproximación Tafonómica a los Restos de Peces en Laguna Llancanelo (Mendoza). Revista Chilena de Antropología, 29(1), 116-121. https://doi.org/10.5354/rca.v0i29.36216 [ Links ]
29. Corbat, M., Giardina, M. y Zangrando, A.F. (2017). The influence of fish bone morphology on aquatic transport: An experimental approach through elements of Creole perch (Percichthyidae: Percichthys trucha; [Valenciennes, 1833]. Journal of Archaeological Science: Reports, 14, 252-261. https://doi.org/10.1016/j.jasrep.2017.05.056 [ Links ]
30. Cruz, I. (2003). Paisajes tafonómicos de restos de Aves en el sur de Patagonia continental. Aportes para la interpretación de conjuntos avifaunísticos en registros arqueológicos del Holoceno. Tesis Doctoral, Facultad de Filosofía y Letras, Universidad de Buenos Aires. [ Links ]
31. Cruz, I. (2005). La representación de partes esqueléticas de aves. Patrones naturales e interpretación arqueológica. Archaeofauna 14, 69-81. [ Links ]
32. Dangavs, N.V. (2005). Los ambientes acuáticos de la provincia de Buenos Aires. En R. R. de Barrio, R. O. Etcheverry, M. F. Caballé y E. Llambías (eds.), Relatorio XVI Congreso Geológico Argentino. Geología y Recursos Minerales de la Provincia de Buenos Aires, 219-236. La Plata, Asociación Geológica Argentina. [ Links ]
33. de la Peña, M.R. (1987). Características ecológicas y algunos ambientes que frecuentan las aves Argentinas. Santa Fe: Facultad de Agronomía y Veterinaria, U.N.L. [ Links ]
34. Domingo, M.S., Martín-Perea, D.M., Badgley, C., Cantero, E., López-Guerrero, P., A. Oliver y J.J. Negro (2020). Taphonomic information from the modern vertebrate death assemblage of Doñana National Park, Spain. PLoS ONE, 15(11), e0242082. https://doi.org/10.1371/journal.pone.0242082 [ Links ]
35. Ferrelli, F. y Aliaga, V.S. (2016). Variabilidad de las precipitaciones y sus efectos sobre la respuesta espaciotemporal de cuerpos de agua en la Región Pampeana, Argentina. Huellas, 20, 239-246. http://dx.doi.org/10.19137/huellas-2016-2015 [ Links ]
36. Forte Lay, J., O. Scarpati y A. Capriolo (2008). Precipitation variability and soil water content in Pampean Flatlands (Argentina). Geofísica Internacional, 47 (4), 341 -354. [ Links ]
37. Grayson, D.K. (1984). Quantitative zooarchaeology: topics in the analysis of archaeological faunas. Orlando: Academic Press. [ Links ]
38. Grayson, D.K. (1991). Alpine faunas from the White Mountains, California: adaptative change in the late prehistoric Great Basin? Journal of Archaeological Science, 18, 483-506. https://doi.org/10.1016/0305-4403(91)90039-R [ Links ]
39. Grosman, F. (2008). Una invitación a conocer nuestras lagunas pampeanas. En F. Grosman (ed.), Espejos en la llanura. Nuestras lagunas de la región pampeana, 19-38. Tandil, Universidad Nacional del Centro de la Provincia de Buenos Aires. [ Links ]
40. Gutiérrez, M.A. y Kaufmann, C.A. (2007). Criteria for the identification of formation processes in guanaco (Lama guanicoe) bone assemblages in fluvial-lacustrine environments. Journal of Taphonomy, 5, 151-175. [ Links ]
41. Gutiérrez, M.A., Kaufmann, C..A., González, M.E., Scheifler, N.A., Rafuse D.J., Massigoge, A. y Álvarez, M.C. (2016). The role of small carnivores in the movement of bones: implications for the Pampas archaeofaunal record, Argentina. Archaeological Anthropological Science, 8, 257-276. https://doi.org/10.1007/s12520-015-0272-1 [ Links ]
42. Gutiérrez, M.A., Rafuse, D.J., Álvarez, M.C., Massigoge, A., González, M.E., Scheifler, N.A. y Kaufmann, C.A. (2018). Ten years of actualistic taphonomic research in the Pampas region of Argentina: contributions to regional archaeology. Quaternary International, 492,40-52. https://doi.org/10.1016/j.quaint.2017.09.025 [ Links ]
43. Haynes, G. (1980). Evidence of carnivore gnawing on Pleistocene and recent mammalian bones. Paleobiology, 6, 341-351. https://www.jstor.org/stable/2400350 [ Links ]
44. Hedges, R.E.M. y A.R. Millard (1995). Bones and Groundwater: Towards the modeling of diagenetic processes. Journal of Archaeological Science, 22, 155–164.
45. Kendall, C., Eriksen, A.M.H., Kontopoulos, I., Collins, M.J., y Turner-Walker, G. (2018). Diagenesis of archaeological bone and tooth. Palaeogeography, Palaeoclimatology, Palaeoecology, 491, 21–37. https://doi.org/10.1016/j.palaeo.2017.11.041
46. Kidwell, S.M., y Flessa, K.W. (1995). The Quality of the Fossil Record: Populations, Species, and Communities. Annual Review of Ecology and Systematics, 26, 269-299. https://doi.org/10.1146/annurev.ecolsys.26.1.269 [ Links ]
47. Klein, R.G. y Cruz-Uribe, K. (1984). The Analysis of Animal Bones from Archaeological Sites. Chicago, The University of Chicago Press. [ Links ]
48. Kruse, E. y Rojo, A. (1991). Aspectos hidrológicos preliminares del Complejo Lagunar Hinojo-Las Tunas (Buenos Aires). Pautas de Investigación. Situación Ambiental de la Provincia de Buenos Aires. En Recursos y Rasgos Naturales en la Evaluación Ambiental, tomo 1, pp. 25-42. La Plata: Comisión de Investigaciones Científicas. [ Links ]
49. Livingston, S.D. (1989). The taphonomic interpretation of avian skeletal part frequencies. Journal of Archaeological Science, 16, 537-547. https://doi.org/10.1016/0305-4403(89)90072-1 [ Links ]
50. López, H., Capítulo, A., Casciotta, J. y Iwaszkiw, J. (1991). Caracterización limnologica preliminar de la Laguna El Hinojo (Trenque Lauquen, Provincia de Buenos Aires). Situación Ambiental de la Provincia de Buenos Aires. En Recursos y Rasgos Naturales en la Evaluación Ambiental, tomo 1, pp. 3-23. La Plata: Comisión de Investigaciones Científicas de la Provincia de Buenos Aires. [ Links ]
51. Lyman, R.L. (1994). Vertebrate Taphonomy. Cambridge: Cambridge University Press. [ Links ]
52. Lyman, R.L. (2003). The influence of time averaging and space averaging on the application of foraging theory in zooarchaeology. Journal of Archaeological Science, 30, 595-610. https://doi.org/10.1016/S0305-4403(02)00236-4 [ Links ]
53. Lyman, R. L. (2008). Quantitative Paleozoology. Cambridge: Cambridge University Press. [ Links ]
54. Maiztegui, T., Baigún, C.R.M., Garcia de Souza, J.R y Colautti, D.C. (2016). Invasion status of common carp (Cyprinus carpio) in Argentinean inland waters. Journal of Fish Biology, 89 (1), 417–430. https://doi.org/10.1111/jfb.13014
55. Manahan, S.E. (2007). Introducción a la Química Ambiental. Madrid: Reverte Editorial Sa. [ Links ]
56. Marchionni, L., Magnín, L.A., Hermo, D. y Mosquera, B. (2020). Advances in the definition of environmental contexts in the Deseado Massif (Santa Cruz, Argentina) and its effects on the modern bone record. Journal of Archaeological Science: Reports, 32, 102385. https://doi.org/10.1016/j.jasrep.2020.102385 [ Links ]
57. Massigoge, A., Rafuse, D.J., Álvarez, M.C., González, M.E., Gutiérrez, M.A., Kaufmann, C.A., Scheifler, N.A. (2015). Beached penguins on the Atlantic Coast in the Pampas region of Argentina: taphonomic analysis and implications for the archaeological record. Palaeogeography, Palaeoclimatology, Palaeoecology, 436, 85-95. https://doi.org/10.1016/j.palaeo.2015.06.045 [ Links ]
58. Messineo, P.G., Tonello, M.S., Stutz, S., Tripaldi, A., Scheifler, N.A., Pal, N., Sánchez Vuichard, G. y Navarro, D. (2019). Human Occupations and Related Environment-Climate during the Middle and Late Holocene in Central Pampas of Argentina. Holocene, 29, 244-261. [ Links ]
59. Miller, J.H. (2011). Ghosts of Yellowstone: multi-decadal histories of wildlife populations captured by bones on a modern landscape. PLOS ONE, 6, e18057. https://doi.org/10.1371/journal.pone.0018057. PMID: 21464921 [ Links ]
60. Miller, J.H., Behrensmeyer, A.K., Du, A., Lyons, S.K., Patterson, D., Tóth, A., Villasenor, A., Kanga, E. y Reed, D. (2014). Ecological fidelity of functional traits based on species presence-absence in a modern mammalian bone assemblage (Amboseli). Paleobiology, 40, 560-583. https://doi.org/10.1666/13062 [ Links ]
61. Oliver, J.S. y Graham, R.W. (1994). A catastrophic kill of ice-trapped coots: time-averaged versus scavenger-specific disarticulation patterns. Paleobiology, 20(2), 229-244. [ Links ]
62. Olsen, S.L. y Shipman, P. (1988). Surface modification on bone: trampling versus butchery. Journal of Archaeological Science, 15, 535-553. https://doi.org/10.1016/0305-4403(88)90081-7 [ Links ]
63. Paoli, C. y Giacosa, R. (2003). Características hidrológicas de la llanura pampeana central oeste (áreas de derrames del Río Quinto y arroyos del sur de Córdoba. En Maiola, O., Gabellone, A. y Hernández, M. (Eds.). Inundaciones en la Región Pampeana. Buenos Aires: Editorial de la Universidad Nacional de La Plata. Universidad Nacional de la Plata, Cámara de Diputados de la Provincia de Buenos Aires, 2003. [ Links ]
64. Petracci, P., Belenguer, F., Sotelo, M., Marbán, L., Belhey, K., y Pérez, C. (2020). Nidificación del flamenco (Phoenicopteurs chilensis Molina, 1782) en un ambiente marino-costero de la Bahía Anegada, provincia Buenos Aires, Argentina. Historia Natural, 10(2), 107-121. [ Links ]
65. Prassack, K.A. (2011). The effect of weathering on bird bone survivorship in modern and fossil saline-alkaline lake environments. Paleobiology, 37(4), 633-654. http://dx.doi.org/10.1666/10041.1 [ Links ]
66. Prego, A.J. (1965), Ruggiero, R.A., Rial Alberti, F., Alonso, R.A. (1965). Estabilización de médanos mediante forestación en la región pampeana semiárida (2a contribución). IDIA. Suplemento forestal 2, 72-98. Publicación Nº 100 del Instituto de Suelos y Agrotecnica. INTA. Buenos Aires. [ Links ]
67. Ringuelet, R. (1961). Rasgos fundamentales de la zoogeografía de la Argentina. Physis, 22(63): 151-170. [ Links ]
68. Rodríguez-Hidalgo, A., Saladié, P., Marín, J. y Canals, A. (2016). Bird-bone modifications by Iberian lynx: A taphonomic analysis of non-ingested red-legged partridge remains. Quaternary International, 421(9), 228-238. https://doi.org/10.1016/j.quaint.2015.11.078 [ Links ]
69. Scheifler, N.A. (2019). Ecología y Subsistencia de los Cazadores-Recolectores en el Campo de Dunas del Centro Pampeano. Buenos Aires: Sociedad Argentina de Antropología: pp. 267. [ Links ]
70. Scheifler, N.A. y Messineo, P.G. (2016). Exploitation of faunal resources by hunter-gatherers in the center of the Pampa grasslands during the Holocene. The archaeofauna of the laguna Cabeza de Buey 2 site (San Carlos de Bolívar, Buenos Aires, Argentina). Quaternary International, 391, 61-73. https://doi.org/10.1016/j.quaint.2015.08.078 [ Links ]
71. Scheifler, N.A., Massigoge, A., Kaufmann, C.A., Rafuse, D.J., González, M.E., Gutiérrez, M.A. y Álvarez, M.C. (2020). Modern bone distribution in the Pampas of Argentina: Taphonomic implications for the regional archaeological record. En Martínez, S. (Ed.). Actualistic Taphonomy in South America, pp. 193-220. Topics in Geobiology. Nueva York: Springer. https://link.springer.com/book/10.1007/978-3-030-20625-3 [ Links ]
72. Scheifler, N.A., Messineo, P.G. Tripaldi, A., González, M., Ozán, I.L., Santos Valero, F., Marini, N. y Politis, G.G. (2022). Estado actual de las investigaciones en la localidad arqueológica Laguna Chica (Sistema Lagunar Hinojo-Las Tunas, Trenque Lauquen). Manuscrito inédito. [ Links ]
73. Shipman, P. (1981). Life history of a fossil. An introduction to taphonomy and paleoecology. Cambridge: Harvard University Press. [ Links ]
74. Somoza, G.M., L.A., Miranda, G.E., Berasain, D.C., Colautti, M., Remes-Lenicov y Strüssmann, C. A., (2008). Historical aspects, current status and prospects of pejerrey aquaculture in South America. Aquaculture Research, 39, 784–793.
75. Soriano, A., León, R.J.C., Sala, O.E., Lavado, R.S., Deregibus, V.A., Cahuepé, M.A., Scaglis, O.A., Velázquez, C.A. y Lemcoff, J.H. (1992). Río de la Plata grasslands. En Coupland, R.T. (Ed.). Ecosystems of the world. Natural grasslands, pp. 367-407. Nueva York: Elsevier. [ Links ]
76. Svoboda, A. y Moreno. E. (2014). Experimentación Sobre los Efectos de la Meteorización en la Supervivencia de Elementos Óseos de Percichthys trucha: Implicaciones Ictioarqueológicas para el Sitio DV1, Lago Musters (Prov. Chubut, Argentina). Revista Chilena de Antropología, 29, 5-7. https://doi.org/10.5354/rca.v0i29.36208 [ Links ]
77. Stewart, K.M. (1991). Modern fishbone assemblages at lake Turkana, Kenya: A methodology to aid in recognition of hominid fish utilization. Journal of Archaeological Science, 18, 579-603. https://doi.org/10.1016/0305-4403(91)90054-Swiens [ Links ]
78. Taccari, I. (1948). El problema forestal en la Pampa semiárida. En La Erosión Eólica en la Región Pampeana y Plan para Conservación de los Suelos. Publicación Miscelánea Nº 303 del Instituto de Suelos y Agrotécnica. INTA. Buenos Aires. [ Links ]
79. Trueman, C.N., Behrensmeyer, A.K., Tuross, N. y Weiner, S. (2004). Mineralogical and compositional changes in bones exposed on soil surfaces in Amboseli National Park, Kenya: diagenetic mechanisms and the role of sediment pore fluids. Journal of Archaeological Science, 31, 721-739. https://doi.org/10.1016/j.jas.2003.11.003 [ Links ]
80. Wiens, J.A. (1989). The ecology of bird communities. Cambridge University Press, Cambridge. [ Links ]
81. Zangrando, A.F. (2003) Ictioarqueología del canal Beagle, explotación de peces y su implicación en la subsistencia humana. Sociedad Argentina de Antropología, Buenos Aires. [ Links ]
82. Zárate, M.A. y Rabassa, J. (2005). Geomorfología de la provincia de Buenos Aires. En de Barrio, R.E., Echeverri, R.O., Caballé, M.F. y Llambias, E. (Eds.). Geología y Recursos Minerales de la provincia de Buenos Aires. Relatorio del 16° Congreso Geológico Argentino, 119-138. La Plata. [ Links ]
83. Zárate, M.A. y Tripaldi, A. (2012). The aeolian system of central Argentina. Aeolian Research, 3, 401-417. https://doi.org/10.1016/j.aeolia.2011.08.002 [ Links ]
84. Zohar, I., Belmaker, M., Nadel, D., Gafny, S., Goren, M., Hershkovitz, I. y Dayan, T. (2008). The living and the dead: How do taphonomic processes modify relative abundance and skeletal completeness of freshwater fish? Palaeogeography, Palaeoclimatology, Palaeoecology, 258, 292–316. DOI: 10.1016/j.palaeo.2007.11.004

